
Genetic Profile in Genes Associated with Cardiorespiratory Fitness in Elite Spanish Male Endurance Athletes




Abstract
:1. Introduction
2. Materials and Methods
2.1. Design
2.2. Subjects
2.3. Genetic Analyses
2.3.1. ACE Genotyping
2.3.2. NOS3 Genotyping
2.3.3. ADRA2A Genotyping
2.3.4. ADRB2 Genotyping
2.3.5. BDKRB2 Genotyping
2.4. TGS Determination
2.5. Statistical Analyses
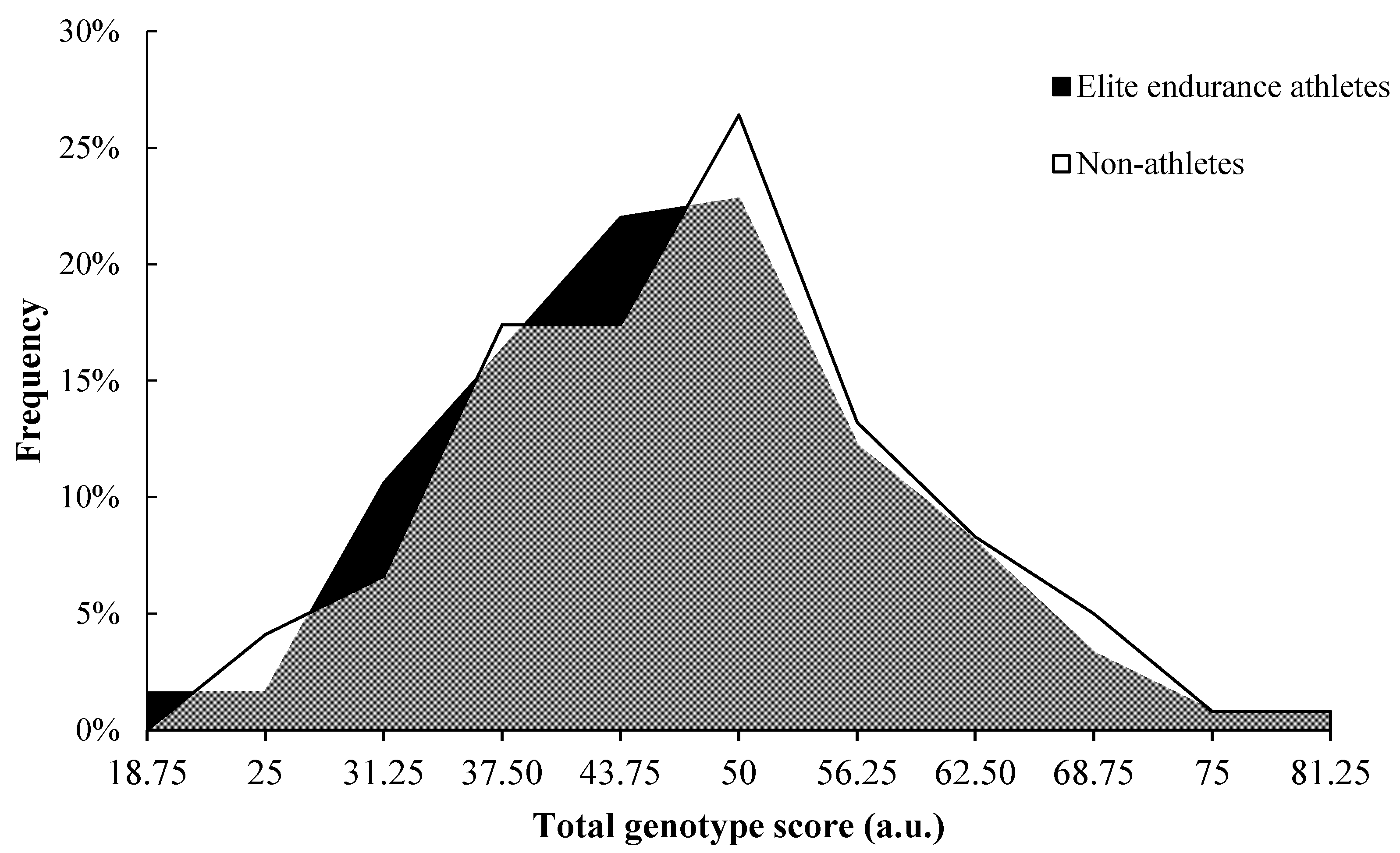
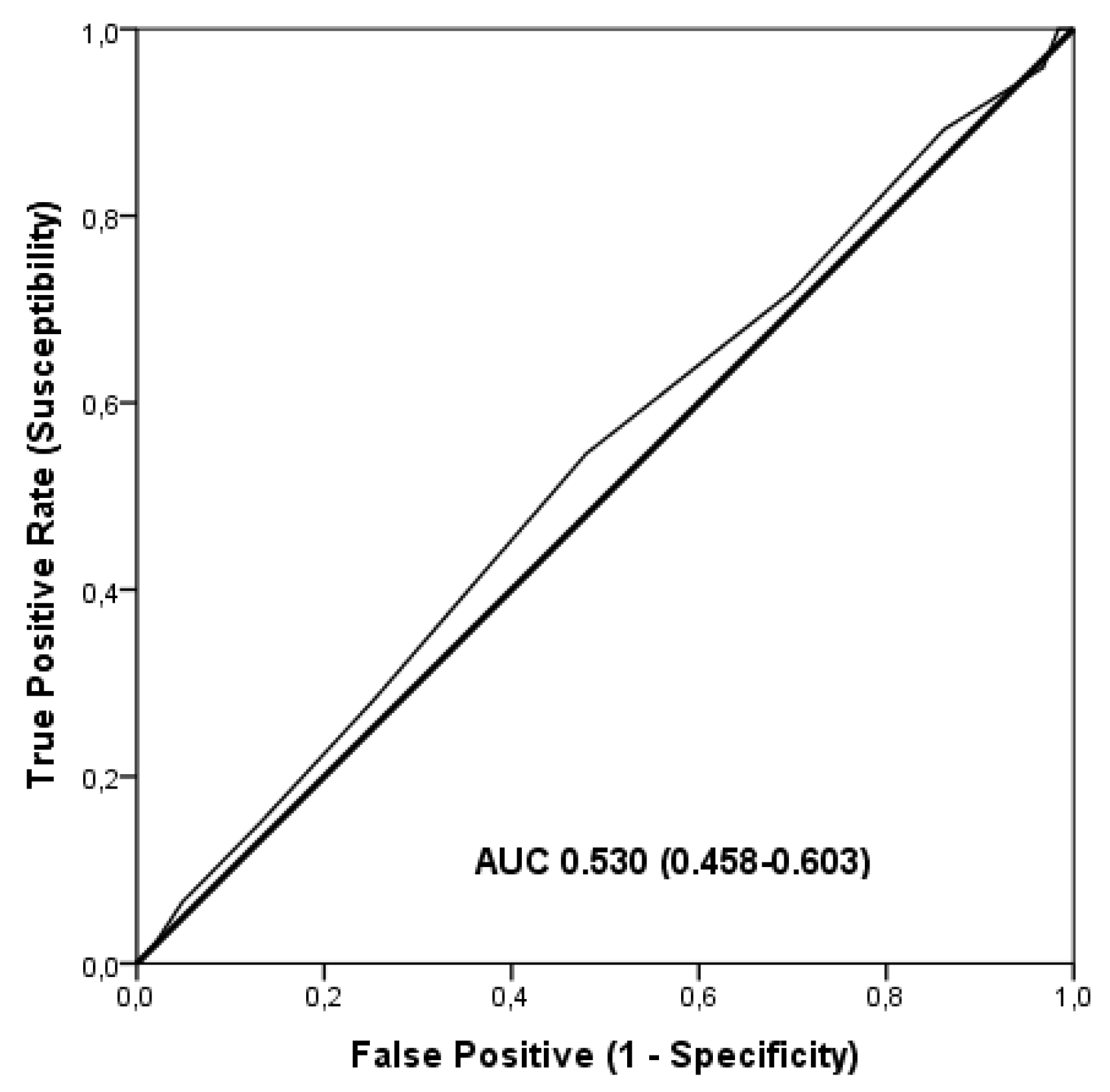
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eynon, N.; Hanson, E.D.; Lucia, A.; Houweling, P.J.; Garton, F.; North, K.N.; Bishop, D.J. Genes for elite power and sprint performance: ACTN3 leads the way. Sports Med. 2013, 43, 803–817. [Google Scholar] [CrossRef]
- Loos, R.J.; Hagberg, J.M.; Perusse, L.; Roth, S.M.; Sarzynski, M.A.; Wolfarth, B.; Rankinen, T.; Bouchard, C. Advances in exercise, fitness, and performance genomics in 2014. Med. Sci. Sports Exerc. 2015, 47, 1105–1112. [Google Scholar] [CrossRef] [Green Version]
- Ben-Zaken, S.; Meckel, Y.; Nemet, D.; Eliakim, A. Genetic score of power-speed and endurance track and field athletes. Scand. J. Med. Sci. Sports 2015, 25, 166–174. [Google Scholar] [CrossRef]
- Egorova, E.S.; Borisova, A.V.; Mustafina, L.J.; Arkhipova, A.A.; Gabbasov, R.T.; Druzhevskaya, A.M.; Astratenkova, I.V.; Ahmetov, I.I. The polygenic profile of Russian football players. J. Sports Sci. 2014, 32, 1286–1293. [Google Scholar] [CrossRef]
- Grealy, R.; Herruer, J.; Smith, C.L.; Hiller, D.; Haseler, L.J.; Griffiths, L.R. Evaluation of a 7-Gene Genetic Profile for Athletic Endurance Phenotype in Ironman Championship Triathletes. PLoS ONE 2015, 10, e0145171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmetov, I.I.; Fedotovskaya, O.N. Current Progress in Sports Genomics. Adv. Clin. Chem. 2015, 70, 247–314. [Google Scholar] [CrossRef]
- Varillas Delgado, D.; Telleria Orriols, J.J.; Martin Saborido, C. Liver-Metabolizing Genes and Their Relationship to the Performance of Elite Spanish Male Endurance Athletes; a Prospective Transversal Study. Sports Med. Open 2019, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and Athletic Performance: An Update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar] [CrossRef]
- Jacob, Y.; Spiteri, T.; Hart, N.H.; Anderton, R.S. The Potential Role of Genetic Markers in Talent Identification and Athlete Assessment in Elite Sport. Sports 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, C.; Ruiz, J.R.; Muniesa, C.A.; Gonzalez-Freire, M.; Gomez-Gallego, F.; Lucia, A. Does the polygenic profile determine the potential for becoming a world-class athlete? Insights from the sport of rowing. Scand. J. Med. Sci. Sports 2010, 20, e188–e194. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.R.; Arteta, D.; Buxens, A.; Artieda, M.; Gomez-Gallego, F.; Santiago, C.; Yvert, T.; Moran, M.; Lucia, A. Can we identify a power-oriented polygenic profile? J. Appl. Physiol. 2010, 108, 561–566. [Google Scholar] [CrossRef]
- Del Coso, J.; Valero, M.; Salinero, J.J.; Lara, B.; Gallo-Salazar, C.; Areces, F. Optimum polygenic profile to resist exertional rhabdomyolysis during a marathon. PLoS ONE 2017, 12, e0172965. [Google Scholar] [CrossRef]
- Cieszczyk, P.; Krupecki, K.; Maciejewska, A.; Sawczuk, M. The angiotensin converting enzyme gene I/D polymorphism in Polish rowers. Int. J. Sports Med. 2009, 30, 624–627. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou, I.D.; Lockey, S.J.; Voisin, S.; Herbert, A.J.; Garton, F.; Houweling, P.J.; Cieszczyk, P.; Maciejewska-Skrendo, A.; Sawczuk, M.; Massidda, M.; et al. No association between ACTN3 R577X and ACE I/D polymorphisms and endurance running times in 698 Caucasian athletes. BMC Genom. 2018, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gallego, F.; Ruiz, J.R.; Buxens, A.; Artieda, M.; Arteta, D.; Santiago, C.; Rodriguez-Romo, G.; Lao, J.I.; Lucia, A. The -786 T/C polymorphism of the NOS3 gene is associated with elite performance in power sports. Eur. J. Appl. Physiol. 2009, 107, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Drozdovs’ka, S.B.; Lysenko, O.M.; Dosenko, V.; Il’in, V.M.; Moibenko, O.O. T(-786) --> C-polymorphism of the endothelial nitric oxide synthase promoter gene (eNOS) and exercise performance in sport. Fiziol. Zh. 2013, 59, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.; Song, X.; Xue, H.; Liu, Z.; Wang, X.; Wang, H.; Sun, K.; Bai, Y.; Liu, J.; Hui, R. A common variant of the eNOS gene (E298D) is an independent risk factor for left ventricular hypertrophy in human essential hypertension. Clin. Sci. 2009, 117, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfarth, B.; Rivera, M.A.; Oppert, J.M.; Boulay, M.R.; Dionne, F.T.; Chagnon, M.; Gagnon, J.; Chagnon, Y.; Perusse, L.; Keul, J.; et al. A polymorphism in the alpha2a-adrenoceptor gene and endurance athlete status. Med. Sci. Sports Exerc. 2000, 32, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, J.; Komine, H.; Hayashi, K.; Yoshizawa, M.; Otsuki, T.; Shimojo, N.; Miyauchi, T.; Yokoi, T.; Maeda, S.; Tanaka, H. Reduction in alpha-adrenergic receptor-mediated vascular tone contributes to improved arterial compliance with endurance training. Int. J. Cardiol. 2009, 135, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Sarpeshkar, V.; Bentley, D.J. Adrenergic-beta(2) receptor polymorphism and athletic performance. J. Hum. Genet. 2010, 55, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.G.; Dhamrait, S.S.; Wootton, P.T.; Day, S.H.; Hawe, E.; Payne, J.R.; Myerson, S.G.; World, M.; Budgett, R.; Humphries, S.E.; et al. Bradykinin receptor gene variant and human physical performance. J. Appl. Physiol. 2004, 96, 938–942. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.G.; Folland, J.P. Similarity of polygenic profiles limits the potential for elite human physical performance. J. Physiol. 2008, 586, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankinen, T.; Fuku, N.; Wolfarth, B.; Wang, G.; Sarzynski, M.A.; Alexeev, D.G.; Ahmetov, I.I.; Boulay, M.R.; Cieszczyk, P.; Eynon, N.; et al. No Evidence of a Common DNA Variant Profile Specific to World Class Endurance Athletes. PLoS ONE 2016, 11, e0147330. [Google Scholar] [CrossRef] [PubMed]
- Gronek, P.; Gronek, J.; Lulinska-Kuklik, E.; Spieszny, M.; Niewczas, M.; Kaczmarczyk, M.; Petr, M.; Fischerova, P.; Ahmetov, I.I.; Zmijewski, P. Polygenic Study of Endurance-Associated Genetic Markers NOS3 (Glu298Asp), BDKRB2 (−9/+9), UCP2 (Ala55Val), AMPD1 (Gln45Ter) and ACE (I/D) in Polish Male Half Marathoners. J. Hum. Kinet 2018, 64, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Saunders, C.J.; Xenophontos, S.L.; Cariolou, M.A.; Anastassiades, L.C.; Noakes, T.D.; Collins, M. The bradykinin beta 2 receptor (BDKRB2) and endothelial nitric oxide synthase 3 (NOS3) genes and endurance performance during Ironman Triathlons. Hum. Mol. Genet. 2006, 15, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, H.E.; Marshall, R.; Hemingway, H.; Myerson, S.; Clarkson, P.; Dollery, C.; Hayward, M.; Holliman, D.E.; Jubb, M.; World, M.; et al. Human gene for physical performance. Nature 1998, 393, 221–222. [Google Scholar] [CrossRef] [PubMed]
- Myerson, S.; Hemingway, H.; Budget, R.; Martin, J.; Humphries, S.; Montgomery, H. Human angiotensin I-converting enzyme gene and endurance performance. J. Appl. Physiol 1999, 87, 1313–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Yang, Y.; Li, X.; Zhou, F.; Gao, C.; Li, M.; Gao, L. The association of sport performance with ACE and ACTN3 genetic polymorphisms: A systematic review and meta-analysis. PLoS ONE 2013, 8, e54685. [Google Scholar] [CrossRef] [Green Version]
- Franchini, M.; Peyvandi, F.; Mannucci, P.M. The genetic basis of coronary artery disease: From candidate genes to whole genome analysis. Trends Cardiovasc. Med. 2008, 18, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Buxens, A.; Ruiz, J.R.; Arteta, D.; Artieda, M.; Santiago, C.; Gonzalez-Freire, M.; Martinez, A.; Tejedor, D.; Lao, J.I.; Gomez-Gallego, F.; et al. Can we predict top-level sports performance in power vs endurance events? A genetic approach. Scand. J. Med. Sci. Sports 2011, 21, 570–579. [Google Scholar] [CrossRef]
- Drozdovska, S.B.; Dosenko, V.E.; Ahmetov, I.I.; Ilyin, V.N. The association of gene polymorphisms with athlete status in ukrainians. Biol. Sport 2013, 30, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Sawczuk, M.; Maciejewska-Karlowska, A.; Cieszczyk, P.; Skotarczak, B.; Ficek, K. Association of the ADRB2 Gly16Arg and Glu27Gln polymorphisms with athlete status. J. Sports Sci. 2013, 31, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Nunes, R.A.B.; Lima, L.B.; Tanaka, N.I.; da Costa Pereira, A.; Krieger, J.E.; Mansur, A.J. Genetic associations of bradykinin type 2 receptor, alpha-adrenoceptors and endothelial nitric oxide synthase with blood pressure and left ventricular mass in outpatients without overt heart disease. Int. J. Cardiol. Heart Vasc. 2018, 21, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Eynon, N.; Meckel, Y.; Alves, A.J.; Nemet, D.; Eliakim, A. Is there an interaction between BDKRB2 -9/+9 and GNB3 C825T polymorphisms and elite athletic performance? Scand. J. Med. Sci. Sports 2011, 21, e242–e246. [Google Scholar] [CrossRef] [Green Version]
- Grenda, A.; Leonska-Duniec, A.; Cieszczyk, P.; Zmijewski, P. Bdkrb2 gene -9/+9 polymorphism and swimming performance. Biol. Sport 2014, 31, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyner, M.J. Genetic Approaches for Sports Performance: How Far Away Are We? Sports Med. 2019, 49, 199–204. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene | Polymorphism | dbSNP | Genotype Score | Elite Endurance Athletes | Non-Athletes | p Value |
|---|---|---|---|---|---|---|---|
| ACE | Angiotensin I-converting enzyme | Alu 287bp (I/D) | rs4340 | 2 = II | 4.88% | 10.66% | <0.001 |
| 1 = ID | 28.46% | 53.28% | |||||
| 0 = DD | 66.67% | 36.07% | |||||
| NOS3 | Nitric Oxide Synthase 3 | c.-786T/C | rs2070744 | 2 = TT | 50.41% | 31.97% | 0.01 |
| 1 = TC | 29.27% | 44.26% | |||||
| 0 = CC | 20.33% | 23.77% | |||||
| NOS3E298D | Nitric Oxide Synthase 3 | c.894G/T | rs1799983 | 2 = GG | 43.09% | 38.84% | 0.111 |
| 1 = GT | 53.65% | 51.24% | |||||
| 0 = TT | 3.25% | 9.91% | |||||
| ADRA2A | α-2a-adrenoceptor | c.-1291C/G | rs1800544 | 2 = CC | 5.61% | 2.46% | 0.016 |
| 1 = GC | 25.23% | 42.62% | |||||
| 0 = GG | 69.16% | 54.92% | |||||
| ADRA2A | α-2a-adrenoceptor | c.1780A/G | rs553668 | 2 = GG | 76.42% | 71.07% | 0.268 |
| 1 = GA | 21.14% | 22.31% | |||||
| 0 = AA | 2.44% | 6.61% | |||||
| ADRB2R16G | β-2-adrenergic receptor | c.46A/G | rs1042713 | 2 = AA | 8.94% | 8.20% | 0.943 |
| 1 = GA | 55.28% | 54.10% | |||||
| 0 = GG | 35.77% | 37.70% | |||||
| ADRB2Q27E | β-2-adrenergic receptor | c.79C/G | rs1042714 | 2 = CC | 25.20% | 17.21% | 0.003 |
| 1 = CG | 60.98% | 50.82% | |||||
| 0 = GG | 13.82% | 31.97% | |||||
| BDKRB2 | Bradykinin Receptor B2 | +9 pb/−9 pb | rs5810761 | 2 = −9/−9 | 21.14% | 24.59% | 0.084 |
| 1 = −9/+9 | 43.90% | 53.28% | |||||
| 0 = +9/+9 | 34.96% | 22.13% |
| Genotype Score | Elite Endurance Athletes | Non-Athletes | p Value |
|---|---|---|---|
| 0 | 0 (0.0%) | 0 (0.0%) | 0.692 |
| 1 | 0 (0.0%) | 0 (0.0%) | |
| 2 | 0 (0.0%) | 0 (0.0%) | |
| 3 | 2 (1.6%) | 0 (0.0%) | |
| 4 | 2 (3.3%) | 5 (4.1%) | |
| 5 | 13 (13.8%) | 8 (10.7%) | |
| 6 | 20 (30.1%) | 21 (28.1%) | |
| 7 | 27 (52.0%) | 21 (45.5%) | |
| 8 | 28 (74.8%) | 32 (71.9%) | |
| 9 | 15 (87.0%) | 16 (85.1%) | |
| 10 | 10 (95.1%) | 10 (93.4%) | |
| 11 | 4 (98.4%) | 6 (98.3%) | |
| 12 | 1 (99.2%) | 1 (99.2%) | |
| 13 | 1 (100.0%) | 1 (100.0%) | |
| 14 | - | - | |
| 15 | - | - | |
| 16 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varillas-Delgado, D.; Tellería Orriols, J.J.; Del Coso, J. Genetic Profile in Genes Associated with Cardiorespiratory Fitness in Elite Spanish Male Endurance Athletes. Genes 2021, 12, 1230. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081230
Varillas-Delgado D, Tellería Orriols JJ, Del Coso J. Genetic Profile in Genes Associated with Cardiorespiratory Fitness in Elite Spanish Male Endurance Athletes. Genes. 2021; 12(8):1230. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081230
Chicago/Turabian StyleVarillas-Delgado, David, Juan José Tellería Orriols, and Juan Del Coso. 2021. "Genetic Profile in Genes Associated with Cardiorespiratory Fitness in Elite Spanish Male Endurance Athletes" Genes 12, no. 8: 1230. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081230