
When Two plus Two Is More than Four: Evidence for a Synergistic Effect of Fatty Acids on Peroxisome Proliferator—Activated Receptor Activity in a Bovine Hepatic Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fatty Acids
2.2. Cell Culture and Treatments
2.3. Plasmid, Transfection and Luciferase Assay
2.4. Estimate Differential PPAR Activation between Mixtures of Fatty Acids
- (1)
- A weighted average model where it is presupposed that each of the FA (at the dose that maximize PPAR activation when used alone) is in competition with the other to occupy a space in the LBD; thus, their estimated contribution to the total PPAR activation is proportional to their molarity, as greater concentration of a FA would increase the likelihood of a FA binding the LBD. Their combined activation can be estimated as:where FA refers to the activation of PPAR of an individual FA and µM refers to its molarity.
- (2)
- An additive model that presupposes that the effect of combined FA is equal to the sum of the effect of each FA included in the mixture, which can be summarized as:
2.5. Statistical Analysis
3. Results
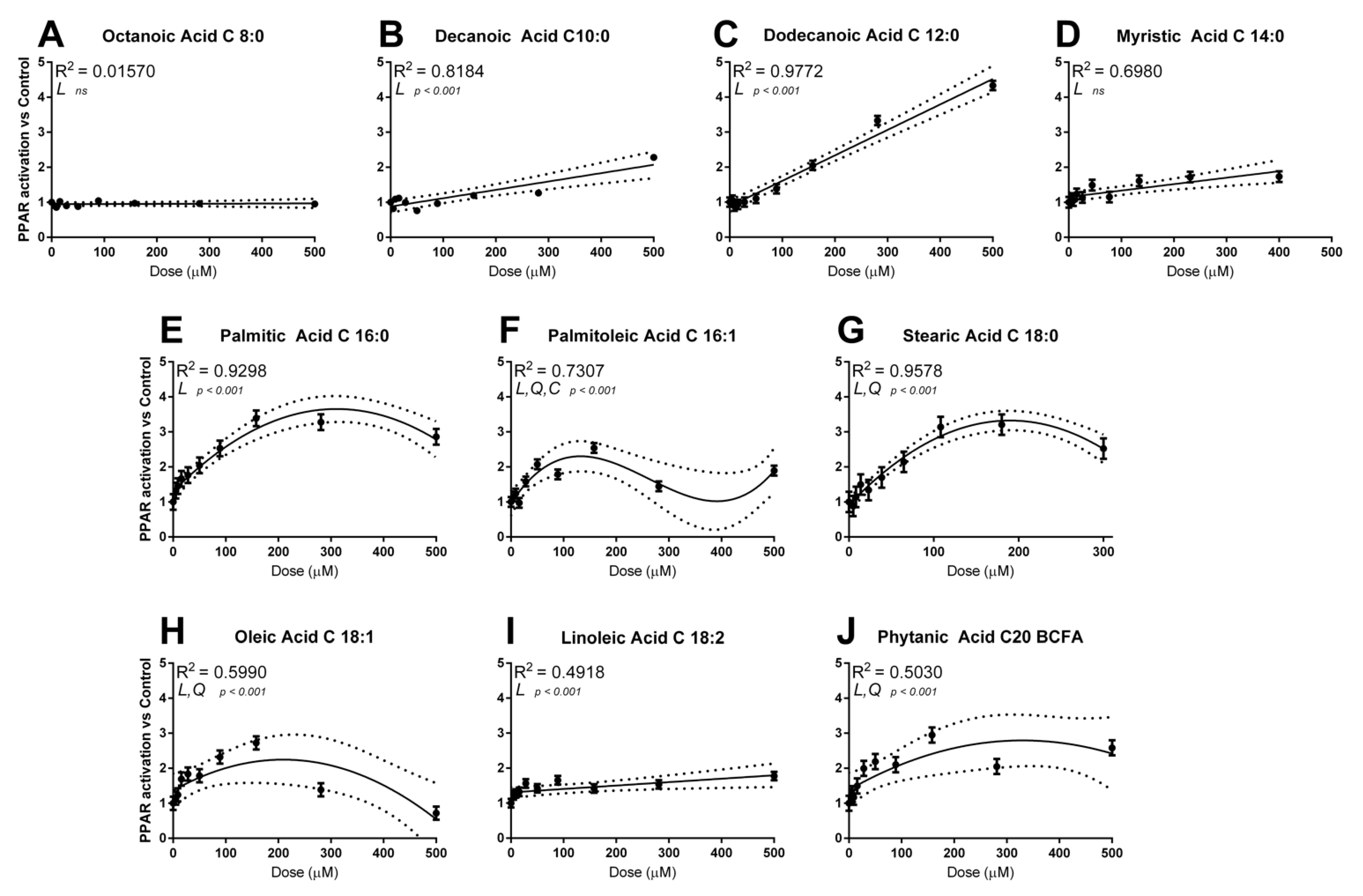
3.1. Dose–Response Activation of PPAR by Single Fatty Acids
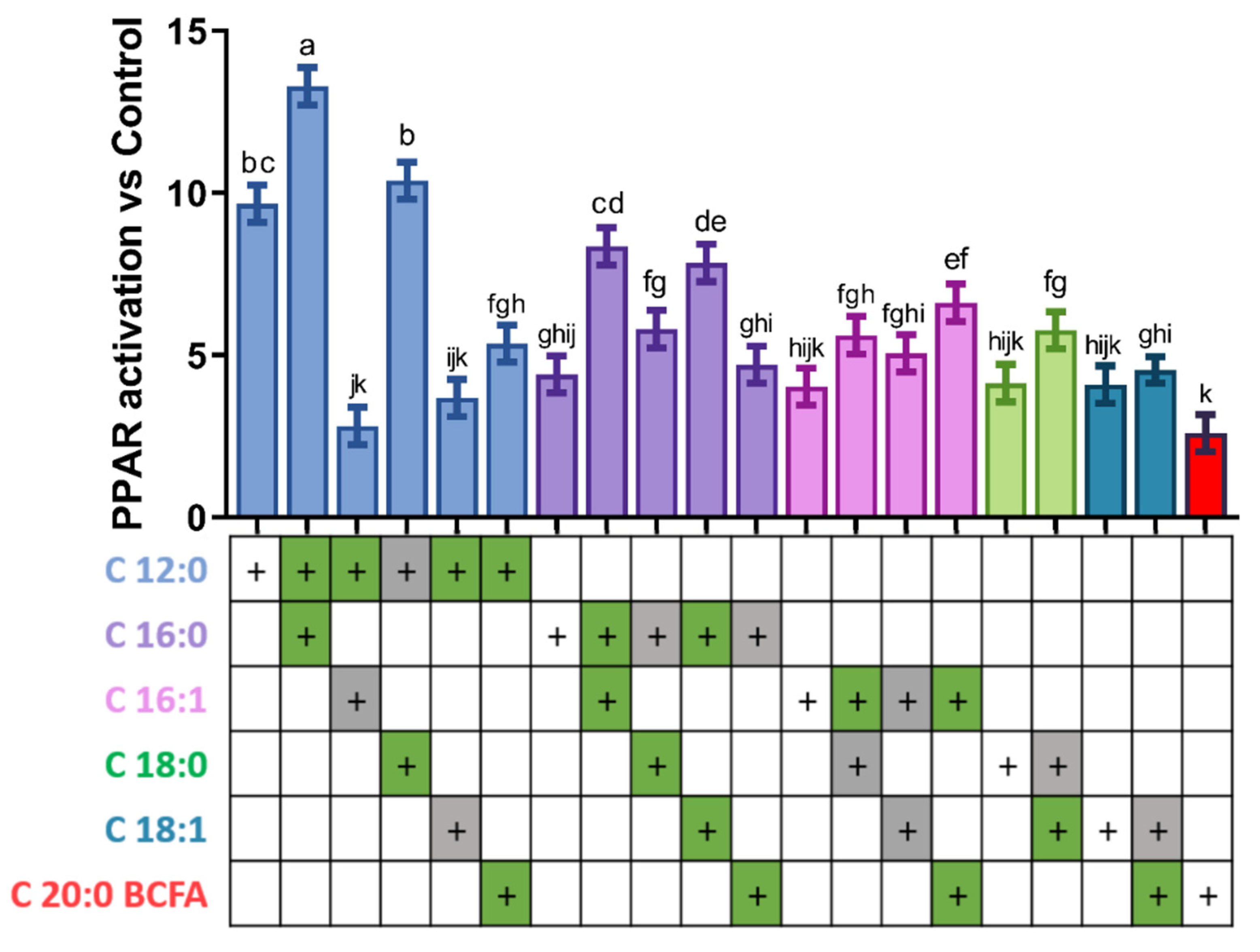
3.2. Activation of PPAR by Combination of Two Fatty Acids
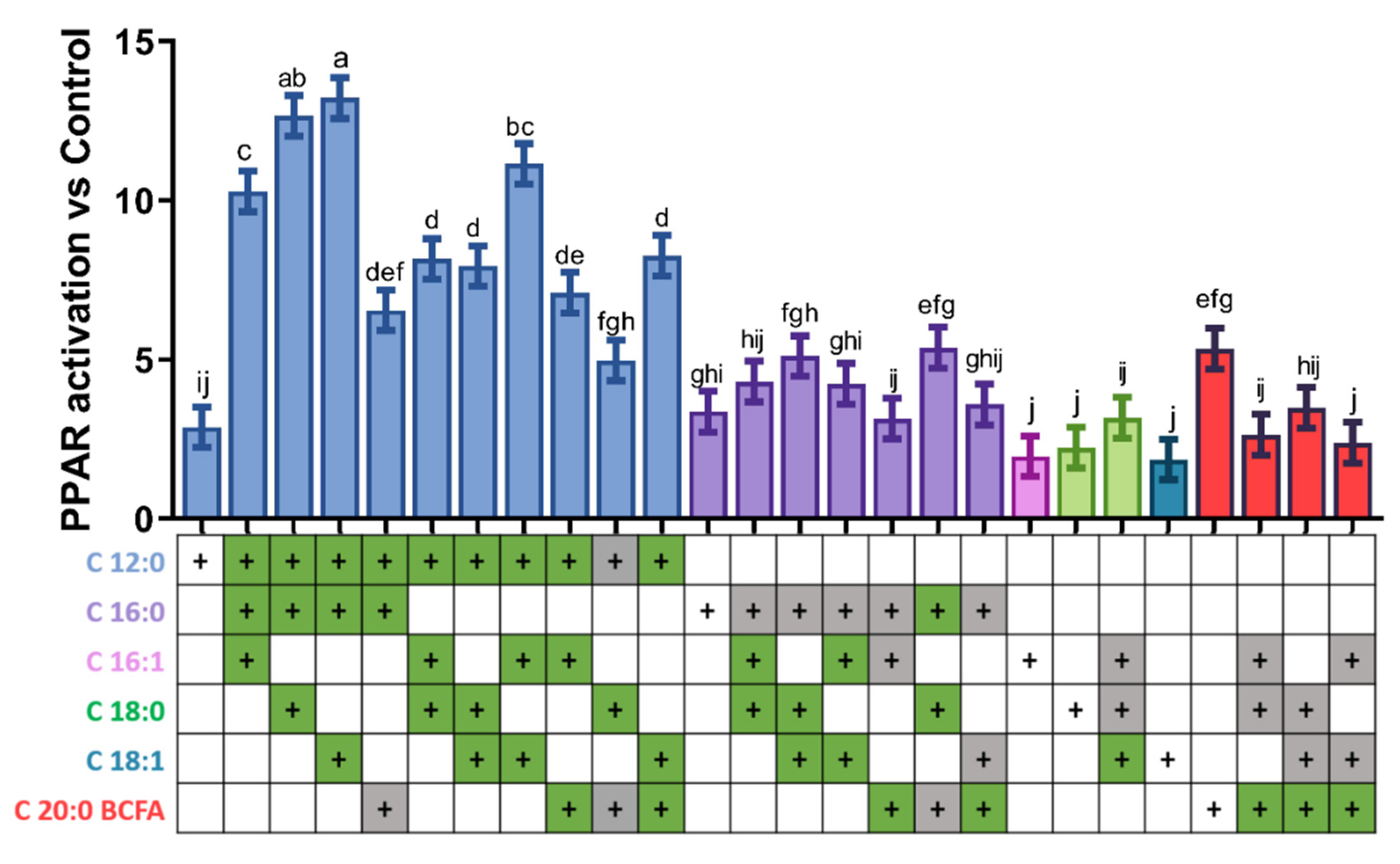
3.3. Activation of PPAR by Combination of Three Fatty Acids
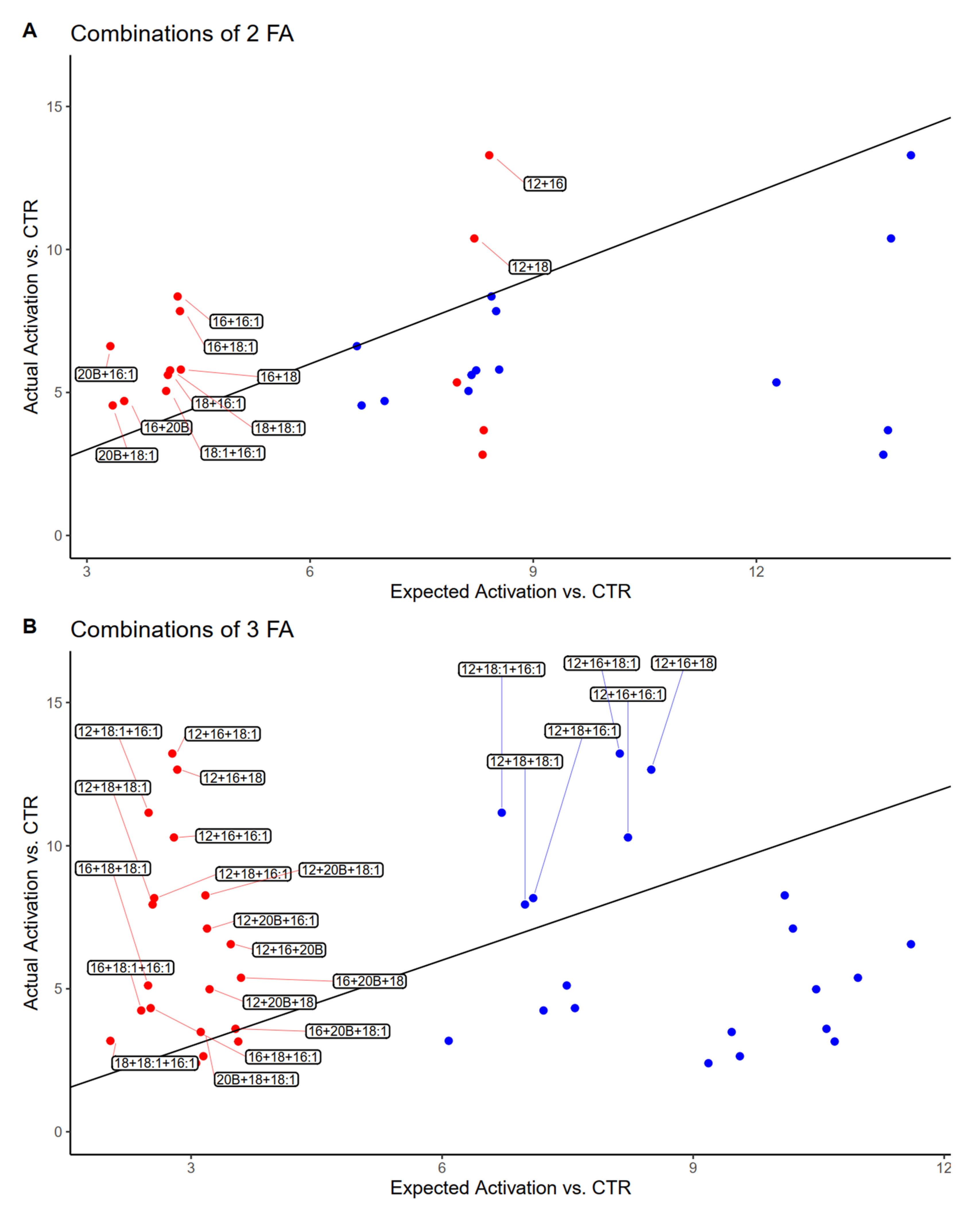
3.4. Estimated vs. Observed Activation of PPAR by Mixtures of Fatty Acids
4. Discussion
4.1. C16:0 and C18:0 Confirmed to Be PPAR Agonists
4.2. C12:0 Is a Strong PPAR Activator in Immortalized Bovine Hepatocytes
4.3. Activation of PPAR Is Differentially Modulated by Mixtures of FA and More Effective When Containing C12:0
4.4. Implications for Nutrigenomic Applications
5. Conclusions
6. Limitations of the Study
- (1)
- As mentioned in the discussion, relative affinity of each of the FA in our study could determine a prevalence in its binding to the LBD. Availability of such information would allow to truly test our hypothetical models of weighted average and additive effect; however, no studies on the affinity of FA to the bovine PPAR LBD are available in the literature;
- (2)
- Both the activity of the PPAR reporter and the activation of PPAR itself are variable on a time scale ([44] is an example of this). Consequently, the fact that the luciferase protocol requires lysis of the cells to obtain a reading does not allow repeated measures over time, and limits the scope of our results to the 18–24 h incubation period. This may explain discrepancies between the results presented in Figure 2 and Figure 3, where rates or PPAR activation in terms of fold change vs. vehicle control are greater than what is presented in Figure 1, as the plate which corresponds to Figure 2 and Figure 3 was incubated for ~2 h longer than that of Figure 1. However, the fact that results from the mixtures all come from the same cell culture plate, combined with the observation that relative differences between fatty acids within the same plates are maintained, suggests that the results from the mixtures can be considered reliable, and biologically relevant within the scope of the comparisons;
- (3)
- The FA supplemented in this study were diluted in culture media, and are thus more suited to mimic a local, concentrated release rather than physiological conditions. Utilizing albumin-bound FA would be more representative of a physiological state; however, it is unclear whether physiological mechanisms for the uptake of albumin-bound FA (discussed in [6]) would be maintained in a cell culture model. Ultimately, our approach provides the greatest degree of response, and as such allows to determine individual differences with greater precision;
- (4)
- BFH-12 are hepatocytes isolated and established from bovine fetuses [18] and are likely not fully representative of adult bovine hepatocytes, since the liver of calves is still maturing after few weeks post-birth [45]. However, the model should be reliable in determining the relative difference in activation and the additive effects of various FA on PPAR activation. While our model is far from perfect, its flexibility allowed us to try a vast number of treatments and combination, which is most suited to the broad scope of our study;
- (5)
- Our interpretation of the results is based on the assumption that activation of PPAR will lead to upregulation of canonical target genes which, as detailed in the introduction, are mostly involved in lipid metabolism; however, we did not measure expression of PPAR targets within the cells, which limits the ability to conclude about biological effects of the observed activation. As previously argued [9], one major limitation on using target genes to assess PPAR activation is the lack of specific bovine PPAR targets. Future endeavors must utilize the results highlighted within this manuscript as a starting point to determine response of PPAR target in models of bovine liver, and other tissues.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pantoja, J.; Firkins, J.L.; Eastridge, M.L.; Hull, B.L. Effects of Fat Saturation and Source of Fiber on Site of Nutrient Digestion and Milk Production by Lactating Dairy Cows1, 2. J. Dairy Sci. 1994, 77, 2341–2356. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Conrad, H.R. High Fat Rations for Dairy Cows. Effects on Feed Intake, Milk and Fat Production, and Plasma Metabolites1. J. Dairy Sci. 1978, 61, 890–901. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Conrad, H.R. High Fat Rations for Dairy Cows. Tallow and Hydrolyzed Blended Fat at Two Intakes1. J. Dairy Sci. 1980, 63, 391–395. [Google Scholar] [CrossRef]
- Maiga, H.A.; Schingoethe, D.J. Optimizing the Utilization of Animal Fat and Ruminal Bypass Proteins in the Diets of Lactating Dairy Cows1. J. Dairy Sci. 1997, 80, 343–352. [Google Scholar] [CrossRef]
- Ohajuruka, O.A.; Wu, Z.; Palmquist, D.L. Ruminal Metabolism, Fiber, and Protein Digestion by Lactating Cows Fed Calcium Soap or Animal-Vegetable Fat1. J. Dairy Sci. 1991, 74, 2601–2609. [Google Scholar] [CrossRef]
- Bionaz, M.; Vargas-Bello-Pérez, E.; Busato, S. Advances in Fatty Acids Nutrition in Dairy Cows: From Gut to Cells and Effects on Performance. J. Anim. Sci. Biotechnol. 2020, 11, 110. [Google Scholar] [CrossRef]
- Bionaz, M.; Chen, S.; Khan, M.J.; Loor, J.J. Functional Role of PPARs in Ruminants: Potential Targets for Fine-Tuning Metabolism during Growth and Lactation. PPAR Res. 2013, 2013, e684159. [Google Scholar] [CrossRef] [Green Version]
- Michalik, L.; Auwerx, J.; Berger, J.P.; Chatterjee, V.K.; Glass, C.K.; Gonzalez, F.J.; Grimaldi, P.A.; Kadowaki, T.; Lazar, M.A.; O’Rahilly, S.; et al. International Union of Pharmacology. LXI. Peroxisome Proliferator-Activated Receptors. Pharmacol. Rev. 2006, 58, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Osorio, J.; Loor, J.J. Triennial Lactation Symposium: Nutrigenomics in Dairy Cows: Nutrients, Transcription Factors, and Techniques. J. Anim. Sci. 2015, 93, 5531–5553. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lin, Y.; Liu, L.; Wang, L.; Bian, Y.; Gao, X.; Li, Q. Regulation of Peroxisome Proliferator-Activated Receptor Gamma on Milk Fat Synthesis in Dairy Cow Mammary Epithelial Cells. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 1044–1059. [Google Scholar] [CrossRef]
- Hihi, A.K.; Michalik, L.; Wahli, W. PPARs: Transcriptional Effectors of Fatty Acids and Their Derivatives. CMLS Cell. Mol. Life Sci. 2002, 59, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Kadegowda, A.K.G.; Bionaz, M.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Peroxisome Proliferator-Activated Receptor-γ Activation and Long-Chain Fatty Acids Alter Lipogenic Gene Networks in Bovine Mammary Epithelial Cells to Various Extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bionaz, M.; Thering, B.J.; Loor, J.J. Fine Metabolic Regulation in Ruminants via Nutrient–Gene Interactions: Saturated Long-Chain Fatty Acids Increase Expression of Genes Involved in Lipid Metabolism and Immune Response Partly through PPAR-α Activation. Br. J. Nutr. 2012, 107, 179–191. [Google Scholar] [CrossRef]
- Busato, S.; Bionaz, M. The Interplay between Non-Esterified Fatty Acids and Bovine Peroxisome Proliferator-Activated Receptors: Results of an in Vitro Hybrid Approach. J. Anim. Sci. Biotechnol. 2020, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Brossillon, V.; Reis, S.F.; Moura, D.C.; Galvão, J.G.B.; Oliveira, A.S.; Côrtes, C.; Brito, A.F. Production, Milk and Plasma Fatty Acid Profile, and Nutrient Utilization in Jersey Cows Fed Flaxseed Oil and Corn Grain with Different Particle Size. J. Dairy Sci. 2018, 101, 2127–2143. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Bello-Pérez, E.; Zhao, W.; Bionaz, M.; Luo, J.; Loor, J.J. Nutrigenomic Effect of Saturated and Unsaturated Long Chain Fatty Acids on Lipid-Related Genes in Goat Mammary Epithelial Cells: What Is the Role of PPARγ? Vet. Sci. 2019, 6, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, T.; Fairall, L.; Amin, K.; Inaba, Y.; Szanto, A.; Balint, B.L.; Nagy, L.; Yamamoto, K.; Schwabe, J.W.R. Structural Basis for the Activation of PPARγ by Oxidized Fatty Acids. Nat. Struct. Mol. Biol. 2008, 15, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Gleich, A.; Kaiser, B.; Honscha, W.; Fuhrmann, H.; Schoeniger, A. Evaluation of the Hepatocyte-Derived Cell Line BFH12 as an in Vitro Model for Bovine Biotransformation. Cytotechnology 2019, 71, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Bionaz, M. Plasmid Transfection in Bovine Cells: Optimization Using a Realtime Monitoring of Green Fluorescent Protein and Effect on Gene Reporter Assay. Gene 2017, 626, 200–208. [Google Scholar] [CrossRef]
- Thering, B.J.; Bionaz, M.; Loor, J.J. Long-Chain Fatty Acid Effects on Peroxisome Proliferator-Activated Receptor-α-Regulated Genes in Madin-Darby Bovine Kidney Cells: Optimization of Culture Conditions Using Palmitate. J. Dairy Sci. 2009, 92, 2027–2037. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Baumrucker, C.R.; Shirk, E.; Vanden Heuvel, J.P.; Block, E.; Varga, G.A. Short Communication: Characterization of Madin-Darby Bovine Kidney Cell Line for Peroxisome Proliferator-Activated Receptors: Temporal Response and Sensitivity to Fatty Acids. J. Dairy Sci. 2008, 91, 2808–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Park, S.K.; Johnson, B.J.; Chung, K.Y.; Choi, C.W.; Kim, K.H.; Kim, W.Y.; Smith, B. AMPKα, C/EBPβ, CPT1β, GPR43, PPARγ, and SCD Gene Expression in Single- and Co-Cultured Bovine Satellite Cells and Intramuscular Preadipocytes Treated with Palmitic, Stearic, Oleic, and Linoleic Acid. Asian-Australas J. Anim. Sci. 2015, 28, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, C.; Keller, H.; Mahfoudi, A.; Laudet, V.; Krey, G.; Wahli, W. Positive Regulation of the Peroxisomal β-Oxidation Pathway by Fatty Acids through Activation of Peroxisome Proliferator-Activated Receptors (PPAR). Biol. Cell 1993, 77, 67–74. [Google Scholar] [CrossRef]
- Abe, T.; Hirasaka, K.; Kohno, S.; Tomida, C.; Haruna, M.; Uchida, T.; Ohno, A.; Oarada, M.; Teshima-Kondo, S.; Okumura, Y.; et al. Capric Acid Up-Regulates UCP3 Expression without PDK4 Induction in Mouse C2C12 Myotubes. J. Nutr. Sci. Vitaminol. 2016, 62, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Liberato, M.V.; Nascimento, A.S.; Ayers, S.D.; Lin, J.Z.; Cvoro, A.; Silveira, R.L.; Martínez, L.; Souza, P.C.T.; Saidemberg, D.; Deng, T.; et al. Medium Chain Fatty Acids Are Selective Peroxisome Proliferator Activated Receptor (PPAR) γ Activators and Pan-PPAR Partial Agonists. PLOS ONE 2012, 7, e36297. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.-J.; Oh, H.; Kang, B.G.; Kang, M.-K.; Kim, D.Y.; Kim, Y.-H.; Lee, J.Y.; Ji, J.G.; Lim, S.S.; Kang, Y.-H. Lipid-Lowering Effects of Medium-Chain Triglyceride-Enriched Coconut Oil in Combination with Licorice Extracts in Experimental Hyperlipidemic Mice. J. Agric. Food Chem. 2018, 66, 10447–10457. [Google Scholar] [CrossRef]
- Graulet, B.; Gruffat-Mouty, D.; Durand, D.; Bauchart, D. Effects of Milk Diets Containing Beef Tallow or Coconut Oil on the Fatty Acid Metabolism of Liver Slices from Preruminant Calves. Br. J. Nutr. 2000, 84, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Arunima, S.; Rajamohan, T. Lauric Acid Beneficially Modulates Apolipoprotein Secretion and Enhances Fatty Acid Oxidation via PPARα-Dependent Pathways in Cultured Rat Hepatocytes. J. Explor. Res. Pharmacol. 2018, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, M.; Oyama, T.; Miyachi, H. Different Structures of the Two Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Ligand-Binding Domains in Homodimeric Complex with Partial Agonist, but Not Full Agonist. Bioorganic Med. Chem. Lett. 2015, 25, 2639–2644. [Google Scholar] [CrossRef]
- Ohashi, M.; Gamo, K.; Oyama, T.; Miyachi, H. Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Has Multiple Binding Points That Accommodate Ligands in Various Conformations: Structurally Similar PPARγ Partial Agonists Bind to PPARγ LBD in Different Conformations. Bioorg. Med. Chem. Lett. 2015, 25, 2758–2762. [Google Scholar] [CrossRef]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; et al. Molecular Recognition of Fatty Acids by Peroxisome Proliferator–Activated Receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Shang, J.; Brust, R.; Mosure, S.A.; Bass, J.; Munoz-Tello, P.; Lin, H.; Hughes, T.S.; Tang, M.; Ge, Q.; Kamenekca, T.M.; et al. Cooperative Cobinding of Synthetic and Natural Ligands to the Nuclear Receptor PPARγ. eLife 2018, 7, e43320. [Google Scholar] [CrossRef]
- Mathews, A.T.; Rico, J.E.; Sprenkle, N.T.; Lock, A.L.; McFadden, J.W. Increasing Palmitic Acid Intake Enhances Milk Production and Prevents Glucose-Stimulated Fatty Acid Disappearance without Modifying Systemic Glucose Tolerance in Mid-Lactation Dairy Cows. J. Dairy Sci. 2016, 99, 8802–8816. [Google Scholar] [CrossRef] [Green Version]
- Piantoni, P.; Lock, A.L.; Allen, M.S. Palmitic Acid Increased Yields of Milk and Milk Fat and Nutrient Digestibility across Production Level of Lactating Cows. J. Dairy Sci. 2013, 96, 7143–7154. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Hausman, G.J.; Loor, J.J.; Mandard, S. Physiological and Nutritional Roles of PPAR across Species. PPAR Res. 2013, 2013, 807156. [Google Scholar] [CrossRef] [PubMed]
- Kadegowda, A.K.G.; Khan, M.J.; Piperova, L.S.; Teter, B.B.; Rodriguez-Zas, S.L.; Erdman, R.A.; Loor, J.J. Trans-10, Cis 12-Conjugated Linoleic Acid-Induced Milk Fat Depression Is Associated with Inhibition of PPAR Signaling and Inflammation in Murine Mammary Tissue. Available online: https://www.hindawi.com/journals/jl/2013/890343/ (accessed on 11 April 2020).
- Bai, X.; Zheng, Z.; Liu, B.; Ji, X.; Bai, Y.; Zhang, W. Whole Blood Transcriptional Profiling Comparison between Different Milk Yield of Chinese Holstein Cows Using RNA-Seq Data. BMC Genom. 2016, 17, 512. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Shi, L.; Liu, L.; Liang, R.; Li, Q.; Li, J.; Han, B.; Sun, D. Analysis of Liver Proteome and Identification of Critical Proteins Affecting Milk Fat, Protein, and Lactose Metabolism in Dariy Cattle with ITRAQ. Proteomics 2019, 19, 1800387. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liang, R.; Li, Y.; Gao, Y.; Li, Q.; Sun, D.; Li, J. Identification of Candidate Genes for Milk Production Traits by RNA Sequencing on Bovine Liver at Different Lactation Stages. BMC Genet. 2020, 21, 72. [Google Scholar] [CrossRef]
- Gessner, D.K.; Gröne, B.; Rosenbaum, S.; Most, E.; Hillen, S.; Becker, S.; Erhardt, G.; Reiner, G.; Eder, K. Treatment of Lactating Sows with Clofibrate as a Synthetic Agonist of PPARα Does Not Influence Milk Fat Content and Gains of Litters. BMC Vet. Res. 2015, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Faciola, A.P.; Broderick, G.A. Effects of Feeding Lauric Acid or Coconut Oil on Ruminal Protozoa Numbers, Fermentation Pattern, Digestion, Omasal Nutrient Flow, and Milk Production in Dairy Cows1. J. Dairy Sci. 2014, 97, 5088–5100. [Google Scholar] [CrossRef]
- Hristov, A.N.; Vander Pol, M.; Agle, M.; Zaman, S.; Schneider, C.; Ndegwa, P.; Vaddella, V.K.; Johnson, K.; Shingfield, K.J.; Karnati, S.K.R. Effect of Lauric Acid and Coconut Oil on Ruminal Fermentation, Digestion, Ammonia Losses from Manure, and Milk Fatty Acid Composition in Lactating Cows. J. Dairy Sci. 2009, 92, 5561–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, A.N.; Lee, C.; Cassidy, T.; Long, M.; Heyler, K.; Corl, B.; Forster, R. Effects of Lauric and Myristic Acids on Ruminal Fermentation, Production, and Milk Fatty Acid Composition in Lactating Dairy Cows. J. Dairy Sci. 2011, 94, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.S.; Lee, W.J.; Kang, E.S.; Ham, S.A.; Yoo, T.; Paek, K.S.; Lim, D.S.; Do, J.T.; Seo, H.G. Ligand-Activated Peroxisome Proliferator-Activated Receptor- δ and-γ Inhibit Lipopolysaccharide-Primed Release of High Mobility Group Box 1 through Upregulation of SIRT1. Cell Death Dis. 2014, 5, e1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, F.; Busato, S.; Avaroma, F.C.; Linville, K.; Trevisi, E.; Osorio, J.S.; Bionaz, M. Transcriptional Changes Detected in Fecal RNA of Neonatal Dairy Calves Undergoing a Mild Diarrhea Are Associated with Inflammatory Biomarkers. PLOS ONE 2018, 13, e0191599. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination | Measured | Expected | |||
|---|---|---|---|---|---|
| Weighted Aver. | %Δ | Additive | %Δ | ||
| 12:0 + 16:0 + 16:1 | 10.3 | 2.8 | 268 | 8.2 | 25 |
| 12:0 + 16:0 + 18:1 | 13.2 | 2.8 | 376 | 8.1 | 63 |
| 12:0 + 16:0 + 18:0 | 12.7 | 2.8 | 346 | 8.5 | 49 |
| 12:0 + 16:1 + 18:0 | 8.2 | 2.6 | 219 | 7.1 | 15 |
| 12:0 + 16:1 + 18:1 | 11.2 | 2.5 | 348 | 6.7 | 66 |
| 12:0 + 18:0 + 18:1 | 7.9 | 2.5 | 213 | 7.0 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busato, S.; Bionaz, M. When Two plus Two Is More than Four: Evidence for a Synergistic Effect of Fatty Acids on Peroxisome Proliferator—Activated Receptor Activity in a Bovine Hepatic Model. Genes 2021, 12, 1283. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081283
Busato S, Bionaz M. When Two plus Two Is More than Four: Evidence for a Synergistic Effect of Fatty Acids on Peroxisome Proliferator—Activated Receptor Activity in a Bovine Hepatic Model. Genes. 2021; 12(8):1283. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081283
Chicago/Turabian StyleBusato, Sebastiano, and Massimo Bionaz. 2021. "When Two plus Two Is More than Four: Evidence for a Synergistic Effect of Fatty Acids on Peroxisome Proliferator—Activated Receptor Activity in a Bovine Hepatic Model" Genes 12, no. 8: 1283. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081283