
Common and Unique Genetic Background between Attention-Deficit/Hyperactivity Disorder and Excessive Body Weight

 , , , , ,
, , , , ,
Abstract
:1. Introduction
- (1)
- Obesity and/or obesity-related factors (e.g., impaired sleep breathing) lead to symptoms of ADHD;
- (2)
- ADHD increases the risk of obesity (e.g., impulsivity and deficits in executive functions favor inappropriate eating behavior);
- (3)
- -
- Polymorphisms of a unique set of genes selected based on gene prioritization tools are associated with the risk of ADHD;
- -
- There are relationships between the studied polymorphisms and overweight and obesity named excessive body weight (EBW);
- -
- The use of whole-exome sequencing allows for the determination of common and unique as well as rare and truncating protein variants between ADHD and EBW.
2. Materials and Methods
2.1. Ethical Statement
2.2. Participants
2.3. Anthropometric Data
2.4. Biological Material Collection and DNA Extraction
2.5. Gene and SNP Selection
2.6. Genotyping
2.7. Statistical Analysis
2.8. WES Sequencing
2.9. Bioinformatic Analysis
3. Results
3.1. Demographic Characteristic
3.2. Genotyping
- -
- ADRA2A—adrenoceptor α 2A gene (rs1800544 p = 0.018 for alleles),
- -
- KCNIP1—potassium voltage-gated channel interacting protein 1 gene (rs1541665 p = 0.044 for genotypes; p = 0.015 for alleles),
- -
- MTHFR—methylenetetrahydrofolate reductase gene (rs1801131 p = 0.013 for genotypes),
- -
- SLC1A3—solute carrier family 1 member 3 gene (rs1049522 p = 0.028 for genotypes; p = 0.022 for alleles),
- -
- SLC6A2—solute carrier family 6 member 2 gene (rs5569 p = 0.029 for alleles).
3.3. Rare and Protein-Truncating Variants
4. Discussion
4.1. Genes and Polymorphisms
4.2. Rare and Protein-Truncating Variant (PTV)
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADHD | Attention-Deficit/Hyperactivity Disorder |
| BMI | Body Mass Index |
| EBW | Excessive Body Weight |
| NBW | Normal Body Weight |
| IOTF | International Obesity Task Force |
| SNP | Single Nucleotide Polymorphism |
| WES | Whole Exome Sequencing |
References
- Cortese, S. The Association between ADHD and Obesity: Intriguing, Progressively More Investigated, but Still Puzzling. Brain Sci. 2019, 9, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortese, S.; Tessari, L. Attention-Deficit/Hyperactivity Disorder (ADHD) and Obesity: Update 2016. Curr. Psychiatry Rep. 2017, 19, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortese, S.; Vincenzi, B. Obesity and ADHD: Clinical and neurobiological implications. Curr. Top. Behav. Neurosci. 2012, 9, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Hanc, T. ADHD as a risk factor for obesity. Current state of research. Psychiatr. Pol. 2018, 52, 309–322. [Google Scholar] [CrossRef]
- de Zwaan, M.; Gruss, B.; Muller, A.; Philipsen, A.; Graap, H.; Martin, A.; Glaesmer, H.; Hilbert, A. Association between obesity and adult attention-deficit/hyperactivity disorder in a German community-based sample. Obes. Facts 2011, 4, 204–211. [Google Scholar] [CrossRef]
- Pagoto, S.L.; Curtin, C.; Lemon, S.C.; Bandini, L.G.; Schneider, K.L.; Bodenlos, J.S.; Ma, Y. Association between adult attention deficit/hyperactivity disorder and obesity in the US population. Obesity 2009, 17, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Agranat-Meged, A.N.; Deitcher, C.; Goldzweig, G.; Leibenson, L.; Stein, M.; Galili-Weisstub, E. Childhood obesity and attention deficit/hyperactivity disorder: A newly described comorbidity in obese hospitalized children. Int. J. Eat. Disord. 2005, 37, 357–359. [Google Scholar] [CrossRef]
- Erermis, S.; Cetin, N.; Tamar, M.; Bukusoglu, N.; Akdeniz, F.; Goksen, D. Is obesity a risk factor for psychopathology among adolescents? Pediatrics Int. Off. J. Jpn. Pediatric Soc. 2004, 46, 296–301. [Google Scholar] [CrossRef]
- Altfas, J.R. Prevalence of attention deficit/hyperactivity disorder among adults in obesity treatment. BMC Psychiatry 2002, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.P.; Levy, L.D.; Levitan, R.D. Symptoms of attention deficit hyperactivity disorder in severely obese women. Eat. Weight Disord. EWD 2005, 10, e10–e13. [Google Scholar] [CrossRef]
- Cortese, S. Attention-Deficit/Hyperactivity Disorder And Childhood Obesity, ebook.ecog-obesity.eu ed.; European Childchood Obesity Group: Bruxelles, Belgium, 2015. [Google Scholar]
- Cortese, S.; Morcillo Penalver, C. Comorbidity between ADHD and obesity: Exploring shared mechanisms and clinical implications. Postgrad. Med. 2010, 122, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Nigg, J.T.; Johnstone, J.M.; Musser, E.D.; Long, H.G.; Willoughby, M.T.; Shannon, J. Attention-deficit/hyperactivity disorder (ADHD) and being overweight/obesity: New data and meta-analysis. Clin. Psychol. Rev. 2016, 43, 67–79. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, V.M.; Curran, J.L.; Browne, N.T. The Co-occurrence of Pediatric Obesity and ADHD: An Understanding of Shared Pathophysiology and Implications for Collaborative Management. Curr. Obes. Rep. 2020, 9, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Albayrak, O.; Putter, C.; Volckmar, A.L.; Cichon, S.; Hoffmann, P.; Nothen, M.M.; Jockel, K.H.; Schreiber, S.; Wichmann, H.E.; Faraone, S.V.; et al. Common obesity risk alleles in childhood attention-deficit/hyperactivity disorder. Am. J. Med Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatr. Genet. 2013, 162B, 295–305. [Google Scholar] [CrossRef]
- Choudhry, Z.; Sengupta, S.M.; Grizenko, N.; Thakur, G.A.; Fortier, M.E.; Schmitz, N.; Joober, R. Association between obesity-related gene FTO and ADHD. Obesity 2013, 21, E738–E744. [Google Scholar] [CrossRef]
- Loos, R.J.; Yeo, G.S. The bigger picture of FTO: The first GWAS-identified obesity gene. Nat. Rev. Endocrinol. 2014, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wolanczyk, T.; Kołakowski, A. 2015. Available online: https://tpd-szczecin.home.pl/TPD/images/ADHD/jansen/12.pdf (accessed on 23 August 2021).
- Kaufman, J.; Birmaher, B.; Brent, D.; Rao, U.; Flynn, C.; Moreci, P.; Williamson, D.; Ryan, N. Schedule for Affective Disorders and Schizophrenia for School-Age Children-Present and Lifetime Version (K-SADS-PL): Initial reliability and validity data. J. Am. Acad. Child Adolesc. Psychiatry 1997, 36, 980–988. [Google Scholar] [CrossRef]
- Atkins, M.S.; Pelham, W.E.; Licht, M.H. The differential validity of teacher ratings of inattention/overactivity and aggression. J. Abnorm. Child Psychol. 1989, 17, 423–435. [Google Scholar] [CrossRef]
- Adamowska, S.; Adamowski, T.; Frydecka, D.; Kiejna, A. Diagnostic validity Polish language version of the questionnaire MINI-KID (Mini International Neuropsychiatry Interview for Children and Adolescent). Compr. Psychiatry 2014, 55, 1744–1750. [Google Scholar] [CrossRef]
- Conners, C.K. Conners’ Rating ScalesRevised Technical Manual; Multi Health Systems: North Tonawanda, NY, USA, 2001. [Google Scholar]
- Cole, T.J.; Bellizzi, M.C.; Flegal, K.M.; Dietz, W.H. Establishing a standard definition for child overweight and obesity worldwide: International survey. BMJ 2000, 320, 1240–1243. [Google Scholar] [CrossRef] [Green Version]
- Cole, T.J.; Flegal, K.M.; Nicholls, D.; Jackson, A.A. Body mass index cut offs to define thinness in children and adolescents: International survey. BMJ 2007, 335, 194. [Google Scholar] [CrossRef] [Green Version]
- PrepITL2P. Laboratory protocol for manual purification of DNA from the whole sample. Available online: https://www.dnagenotek.com/ROW/pdf/PD-PR-015.pdf (accessed on 23 August 2021).
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.P.; Oliveira, I.O.; Santos, B.R.; Millech, C.; Silva, L.P.; Gonzalez, D.A.; Hallal, P.C.; Menezes, A.M.; Araujo, C.L.; Barros, F.C. Quality of DNA extracted from saliva samples collected with the Oragene DNA self-collection kit. BMC Med. Res. Methodol. 2012, 12, 65. [Google Scholar] [CrossRef] [Green Version]
- Pardo-Diaz, C.; Salazar, C.; Jiggins, C.D. Towards the identification of the loci of adaptive evolution. Methods Ecol. Evol. 2015, 6, 445–464. [Google Scholar] [CrossRef]
- Moreau, Y.; Tranchevent, L.C. Computational tools for prioritizing candidate genes: Boosting disease gene discovery. Nat. Rev. Genet. 2012, 13, 523–536. [Google Scholar] [CrossRef]
- Tranchevent, L.C.; Capdevila, F.B.; Nitsch, D.; De Moor, B.; De Causmaecker, P.; Moreau, Y. A guide to web tools to prioritize candidate genes. Brief. Bioinform. 2011, 12, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Wulf, A.; Liu, T.; Khoury, M.J.; Gwinn, M. Gene Prospector: An evidence gateway for evaluating potential susceptibility genes and interacting risk factors for human diseases. BMC Bioinform. 2008, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Knox, C.; Young, N.; Stothard, P.; Damaraju, S.; Wishart, D.S. PolySearch: A web-based text mining system for extracting relationships between human diseases, genes, mutations, drugs and metabolites. Nucleic Acids Res. 2008, 36, W399–W405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biosearch. Available online: https://biosearch-cdn.azureedge.net/assetsv6/KASP-genotyping-chemistry-User-guide.pdf (accessed on 23 August 2021).
- Biosearch. Available online: https://biosearch-cdn.azureedge.net/assetsv6/Analysis-of-KASP-genotyping-data-using-cluster-plots.pdf (accessed on 23 August 2021).
- Biosearch. Available online: https://biosearch-cdn.azureedge.net/assetsv6/KASP-thermal-cycling-61-55%C2%BAC-touchdown-protocol.pdf (accessed on 23 August 2021).
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Giliani, S.; Lanzi, G.; Mias, G.I.; Lonardi, S.; Dobbs, K.; Manis, J.; Im, H.; Gallagher, J.E.; Phanstiel, D.H.; et al. Whole-exome sequencing identifies tetratricopeptide repeat domain 7A (TTC7A) mutations for combined immunodeficiency with intestinal atresias. J. Allergy Clin. Immunol. 2013, 132, 656–664.e617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Im, H.; Snyder, M. Whole-Exome Enrichment with the Agilent SureSelect Human All Exon Platform. Cold Spring Harb. Protoc. 2015, 2015, 626–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modi, A.; Vai, S.; Caramelli, D.; Lari, M. The Illumina Sequencing Protocol and the NovaSeq 6000 System. Methods Mol. Biol. 2021, 2242, 15–42. [Google Scholar] [CrossRef] [PubMed]
- Tombacz, D.; Maroti, Z.; Kalmar, T.; Palkovits, M.; Snyder, M.; Boldogkoi, Z. Whole-exome sequencing data of suicide victims who had suffered from major depressive disorder. Sci. Data 2019, 6, 190010. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 23 August 2021).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Picard. 2019. Available online: http://broadinstitute.github.io/picard/ (accessed on 23 August 2021).
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high confidence variant calls: The Genome Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Rovira, P.; Demontis, D.; Sanchez-Mora, C.; Zayats, T.; Klein, M.; Mota, N.R.; Weber, H.; Garcia-Martinez, I.; Pagerols, M.; Vilar-Ribo, L.; et al. Shared genetic background between children and adults with attention deficit/hyperactivity disorder. Neuropsychopharmacology 2020, 45, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Satterstrom, F.K.; Walters, R.K.; Singh, T.; Wigdor, E.M.; Lescai, F.; Demontis, D.; Kosmicki, J.A.; Grove, J.; Stevens, C.; Bybjerg-Grauholm, J.; et al. Autism spectrum disorder and attention deficit hyperactivity disorder have a similar burden of rare protein-truncating variants. Nat. Neurosci. 2019, 22, 1961–1965. [Google Scholar] [CrossRef] [PubMed]
- LGR. Locus Reference Genomic. Available online: https://www.lrg-sequence.org/about/ (accessed on 23 August 2021).
- HGMD. Human Gene Mutation Database. Available online: http://www.hgmd.cf.ac.uk/ac/index.php (accessed on 23 August 2021).
- HUGO. HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/ (accessed on 23 August 2021).
- ENSEMBL. Available online: https://www.ensembl.org/info/genome/variation/prediction/predicted_data.html (accessed on 23 August 2021).
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Povysil, G.; Petrovski, S.; Hostyk, J.; Aggarwal, V.; Allen, A.S.; Goldstein, D.B. Rare-variant collapsing analyses for complex traits: Guidelines and applications. Nat. Rev. Genet. 2019, 20, 747–759. [Google Scholar] [CrossRef]
- Gnanavel, S.; Sharma, P.; Kaushal, P.; Hussain, S. Attention deficit hyperactivity disorder and comorbidity: A review of literature. World J. Clin. Cases 2019, 7, 2420–2426. [Google Scholar] [CrossRef]
- Gershon, J. A meta-analytic review of gender differences in ADHD. J. Atten. Disord. 2002, 5, 143–154. [Google Scholar] [CrossRef]
- Mowlem, F.D.; Rosenqvist, M.A.; Martin, J.; Lichtenstein, P.; Asherson, P.; Larsson, H. Sex differences in predicting ADHD clinical diagnosis and pharmacological treatment. Eur. Child Adolesc. Psychiatry 2019, 28, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Hanc, T.; Dmitrzak-Weglarz, M.; Borkowska, A.; Wolanczyk, T.; Pytlinska, N.; Rybakowski, F.; Slopien, R.; Slopien, A. Overweight in Boys With ADHD Is Related to Candidate Genes and Not to Deficits in Cognitive Functions. J. Atten. Disord. 2018, 22, 1158–1172. [Google Scholar] [CrossRef]
- Skounti, M.; Giannoukas, S.; Dimitriou, E.; Nikolopoulou, S.; Linardakis, E.; Philalithis, A. Prevalence of attention deficit hyperactivity disorder in schoolchildren in Athens, Greece. Association of ADHD subtypes with social and academic impairment. Atten. Deficit Hyperact. Disord. 2010, 2, 127–132. [Google Scholar] [CrossRef]
- Sayal, K.; Prasad, V.; Daley, D.; Ford, T.; Coghill, D. ADHD in children and young people: Prevalence, care pathways, and service provision. Lancet. Psychiatry 2018, 5, 175–186. [Google Scholar] [CrossRef]
- Hvolgaard Mikkelsen, S.; Olsen, J.; Bech, B.H.; Obel, C. Parental age and attention-deficit/hyperactivity disorder (ADHD). Int. J. Epidemiol. 2017, 46, 409–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, F.F.; Gu, X.; Huang, X.; Hou, Y.W.; Zhong, Y.; Lin, J.; Wu, J. Attention-deficit/hyperactivity disorder associated with KChIP1 rs1541665 in Kv channels accessory proteins. PLoS ONE 2017, 12, e0188678. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, Q.; Chen, X.; Gu, X.; Wang, M.; Wu, J. A functional variant in SLC1A3 influences ADHD risk by disrupting a hsa-miR-3171 binding site: A two-stage association study. Genes Brain Behav. 2019, 18, e12574. [Google Scholar] [CrossRef]
- Saha, T.; Chatterjee, M.; Verma, D.; Ray, A.; Sinha, S.; Rajamma, U.; Mukhopadhyay, K. Genetic variants of the folate metabolic system and mild hyperhomocysteinemia may affect ADHD associated behavioral problems. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 84, 1–10. [Google Scholar] [CrossRef]
- Kiive, E.; Kurrikoff, T.; Maestu, J.; Harro, J. Effect of alpha2A-adrenoceptor C-1291G genotype and maltreatment on hyperactivity and inattention in adolescents. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Myer, N.M.; Boland, J.R.; Faraone, S.V. Pharmacogenetics predictors of methylphenidate efficacy in childhood ADHD. Mol. Psychiatry 2018, 23, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- Tortajada-Genaro, L.A.; Mena, S.; Ninoles, R.; Puigmule, M.; Viladevall, L.; Maquieira, A. Genotyping of single nucleotide polymorphisms related to attention-deficit hyperactivity disorder. Anal. Bioanal. Chem. 2016, 408, 2339–2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patte, K.A.; Davis, C.A.; Levitan, R.D.; Kaplan, A.S.; Carter-Major, J.; Kennedy, J.L. A Behavioral Genetic Model of the Mechanisms Underlying the Link Between Obesity and Symptoms of ADHD. J. Atten. Disord. 2020, 24, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Breasson, L.; Becattini, B.; Sardi, C.; Molinaro, A.; Zani, F.; Marone, R.; Botindari, F.; Bousquenaud, M.; Ruegg, C.; Wymann, M.P.; et al. PI3Kgamma activity in leukocytes promotes adipose tissue inflammation and early-onset insulin resistance during obesity. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FBXL17. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=FBXL17 (accessed on 23 August 2021).
- Blekhman, R.; Goodrich, J.K.; Huang, K.; Sun, Q.; Bukowski, R.; Bell, J.T.; Spector, T.D.; Keinan, A.; Ley, R.E.; Gevers, D.; et al. Host genetic variation impacts microbiome composition across human body sites. Genome Biol. 2015, 16, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Silva, T.; Salatino-Oliveira, A.; Genro, J.P.; Meyer, F.D.T.; Li, Y.; Rohde, L.A.; Hutz, M.H.; Tovo-Rodrigues, L. Host genetics influences the relationship between the gut microbiome and psychiatric disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110153. [Google Scholar] [CrossRef] [PubMed]
- Checa-Ros, A.; Jerez-Calero, A.; Molina-Carballo, A.; Campoy, C.; Munoz-Hoyos, A. Current Evidence on the Role of the Gut Microbiome in ADHD Pathophysiology and Therapeutic Implications. Nutrients 2021, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Mathee, K.; Cickovski, T.; Deoraj, A.; Stollstorff, M.; Narasimhan, G. The gut microbiome and neuropsychiatric disorders: Implications for attention deficit hyperactivity disorder (ADHD). J. Med. Microbiol. 2020, 69, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Tabrett, A.; Horton, M.W. The influence of host genetics on the microbiome. F1000Research 2020, 9, Rev-84. [Google Scholar] [CrossRef]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. New Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speakman, J.R. The ‘Fat Mass and Obesity Related’ (FTO) gene: Mechanisms of Impact on Obesity and Energy Balance. Curr. Obes. Rep. 2015, 4, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Velders, F.P.; De Wit, J.E.; Jansen, P.W.; Jaddoe, V.W.; Hofman, A.; Verhulst, F.C.; Tiemeier, H. FTO at rs9939609, food responsiveness, emotional control and symptoms of ADHD in preschool children. PLoS ONE 2012, 7, e49131. [Google Scholar] [CrossRef] [Green Version]
- Amare, A.T.; Schubert, K.O.; Klingler-Hoffmann, M.; Cohen-Woods, S.; Baune, B.T. The genetic overlap between mood disorders and cardiometabolic diseases: A systematic review of genome wide and candidate gene studies. Transl. Psychiatry 2017, 7, e1007. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Matsunaga, T.; Adachi, T.; Aoki, N.; Tsujimoto, G.; Tsuda, K. Adrenergic receptor polymorphisms and autonomic nervous system function in human obesity. Trends Endocrinol. Metab. TEM 2006, 17, 269–275. [Google Scholar] [CrossRef]
- de Cerqueira, C.C.; Polina, E.R.; Contini, V.; Marques, F.Z.; Grevet, E.H.; Salgado, C.A.; da Silva, P.O.; Picon, F.A.; Belmonte-de-Abreu, P.; Bau, C.H. ADRA2A polymorphisms and ADHD in adults: Possible mediating effect of personality. Psychiatry Res. 2011, 186, 345–350. [Google Scholar] [CrossRef]
- DYNC1H1. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=DYNC1H1 (accessed on 23 August 2021).
- Amabile, S.; Jeffries, L.; McGrath, J.M.; Ji, W.; Spencer-Manzon, M.; Zhang, H.; Lakhani, S.A. DYNC1H1-related disorders: A description of four new unrelated patients and a comprehensive review of previously reported variants. Am. J. Med. Genet. Part A 2020, 182, 2049–2057. [Google Scholar] [CrossRef]
- Becker, L.L.; Dafsari, H.S.; Schallner, J.; Abdin, D.; Seifert, M.; Petit, F.; Smol, T.; Bok, L.; Rodan, L.; Krapels, I.; et al. The clinical-phenotype continuum in DYNC1H1-related disorders-genomic profiling and proposal for a novel classification. J. Hum. Genet. 2020, 65, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Fliers, E.A.; Vasquez, A.A.; Poelmans, G.; Rommelse, N.; Altink, M.; Buschgens, C.; Asherson, P.; Banaschewski, T.; Ebstein, R.; Gill, M.; et al. Genome-wide association study of motor coordination problems in ADHD identifies genes for brain and muscle function. World J. Biol. Psychiatry 2012, 13, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.A.; Eberhardt, N.L.; Jensen, M.D. Role of nonexercise activity thermogenesis in resistance to fat gain in humans. Science 1999, 283, 212–214. [Google Scholar] [CrossRef] [Green Version]
- Stray, L.L.; Kristensen, O.; Lomeland, M.; Skorstad, M.; Stray, T.; Tonnessen, F.E. Motor regulation problems and pain in adults diagnosed with ADHD. Behav. Brain Funct. BBF 2013, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SEMA6D. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=SEMA6D&keywords=SEMA6D (accessed on 23 August 2021).
- Jogamoto, T.; Utsunomiya, R.; Sato, A.; Kihara, N.; Choudhury, M.E.; Miyanishi, K.; Kubo, M.; Nagai, M.; Nomoto, M.; Yano, H.; et al. Lister hooded rats as a novel animal model of attention-deficit/hyperactivity disorder. Neurochem. Int. 2020, 141, 104857. [Google Scholar] [CrossRef]
- Che, L.; Hu, Q.; Wang, R.; Zhang, D.; Liu, C.; Zhang, Y.; Xin, G.; Fang, Z.; Lin, Y.; Xu, S.; et al. Inter-correlated gut microbiota and SCFAs changes upon antibiotics exposure links with rapid body-mass gain in weaned piglet model. J. Nutr. Biochem. 2019, 74, 108246. [Google Scholar] [CrossRef] [PubMed]
- MAP1A. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=MAP1A (accessed on 23 August 2021).
- Nowacka-Chmielewska, M.; Liskiewicz, D.; Liskiewicz, A.; Marczak, L.; Wojakowska, A.; Jerzy Barski, J.; Malecki, A. Cerebrocortical proteome profile of female rats subjected to the western diet and chronic social stress. Nutr. Neurosci. 2020, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- ZNF536. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=ZNF536&keywords (accessed on 23 August 2021).
- Collaboration, N.C.D.R.F. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [Green Version]
- OECD. Available online: https://www.oecd.org/health/health-systems/Obesity-Update-2017.pdf (accessed on 23 August 2021).
- Dmitrzak-Weglarz, M.; Tyszkiewicz-Nwafor, M.; Duda, J.; Paszynska, E.; Matuszak-Wojciechowska, L.; Hanc, T.; Bryl, E.; Mamrot, P.; Dutkiewicz, A.; Slopien, A. Abnormal body weight and food-related behavior in school-aged children as measured by the Children’s Binge Eating Disorder Scale. Clin. Child Psychol. Psychiatry 2020, 25, 304–319. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Criteria for Inclusion into the ADHD Group | Criteria for Inclusion into the Non-ADHD Group | Criteria for Exclusion from ADHD and Non-ADHD Groups |
|---|---|---|
| Children of both sexes aged 6–17 | Children of both sexes aged 6–12 | Children with disorders of central nervous system (e.g., epilepsy, serious injuries, and CNS infections) |
| Children with diagnosed ADHD in accordance with ICD-10 and DSM-V diagnostic criteria (diagnosis confirmed by two independent psychiatrists based on a standardized and structured interview) | Lack of mental disorders—assessment with the use of MINI-Kid questionnaire | Co-existing: schizophrenia, bipolar affective disorder, any serious somatic disorders |
| Clinically significant ADHD symptoms lasting over 6 months | Parent or legal guardian approval | Chronic somatic diseases |
| Children without hereditary mental disorders (first-degree relatives) | Persistent pharmacotherapy, hormonotherapy | |
| Parent or legal guardian approval | Lack of acceptance from parents or legal guardians |
| ADHD N = 226 (%) * | non-ADHD N = 482 (%) * | ADHD vs. Non-ADHD p-Value (χ2 Test) | ||
|---|---|---|---|---|
| ADHD type | Combinet | 171 (75.5) | - | - |
| Attention-deficit disorder (ADD) | 39 (17.3) | - | - | |
| Hyperactive/impulsive type (H/I) | 16 (7.0) | - | - | |
| Comorbid disorders | At least one additional diagnosed disorder | 195 (86.3) | 49 (10.2) | <0.000 |
| Learning disorders | 102 (45.1) | 22 (4.6) | <0.000 | |
| Oppositional defiant disorder | 70 (31.0) | - | - | |
| Speech disorders | 56 (24.8) | 27 (5.6) | <0.000 | |
| Tic disorder | 39 (17.3) | 6 (1.2) | <0.000 | |
| Conduct disorder | 25 (11.1) | - | - | |
| Enuresis | 51 (22.6) | - | - | |
| Anxiety disorders | 10 (4.4) | - | - | |
| Mood disorders | 7 (3.1) | - | - | |
| Place of residence | Villages | 50 (22.1) | 35 (7.3) | <0.000 |
| City with less than 100,000 citizens | 66 (29.2) | 89 (18.4) | ||
| City with more than 100,000 citizens | 110 (48.7) | 358 (74.3) | ||
| Mother’s level of education | Elementary | 5 (2.4) | - | <0.000 |
| Vocational | 32 (15.2) | 54 (11.5) | ||
| Secondary | 106 (50.5) | 119 (25.3) | ||
| Higher | 67 (31.9) | 289 (63.3) | ||
| Father’s level of education | Elementary | 10 (4.8) | - | <0.000 |
| Vocational | 69 (33.3) | 103 (22.7) | ||
| Secondary | 83 (40.1) | 133 (29.3) | ||
| Higher | 45 (21.7) | 218 (48.0) | ||
| Mother’s age at child birth | <25 | 67 (33.5) | 89 (18.9) | <0.000 |
| 25–35 | 111 (55.5) | 346 (73.3) | ||
| >35 | 22 (11.0) | 37 (7.8) | ||
| Father’s age at child birth | <25 | 35 (18.0) | 54 (11.7) | 0.015 |
| 25–35 | 126 (64.9) | 350 (75.9) | ||
| >35 | 33 (17.0) | 57 (12.4) | ||
| Socioeconomic status | Low | 15 (7.1) | 16 (3.4) | <0.000 |
| Average | 78 (36.8) | 81 (17.5) | ||
| High | 119 (56.1) | 367 (79.1) | ||
| Birth term | <37 weeks of gestation | 29 (14.9) | 32 (7.1) | <0.000 |
| between 37 and 42 weeks | 138 (70.8) | 417 (92.7) | ||
| >43 weeks of gestation | 28 (14.3) | 1 (0.2) | ||
| The course of childbirth | the forces of nature | 140 (66.3) | 347 (72.0) | 0.028 |
| Caesarean section | 63 (29.9) | 103 (21.4) | ||
| vacuum lift or forceps | 8 (3.8) | 32 (6.6) | ||
| Birth body mass (g) | <2500 | 17 (8.5) | 20 (4.2) | 0.048 |
| 2500–4000 | 164 (81.6) | 390 (82.3) | ||
| >4000 | 20 (9.9) | 64 (13.5) | ||
| Apgar score | <4 | 9 (4.8) | 14 (3.2) | 0.001 |
| 4–7 | 22 (11.7) | 18 (4.1) | ||
| >7 | 157 (83.5) | 411 (92.7) | ||
| Weight status (IOTF) | Underweight | 2 (1.08) | 5 (1.05) | 0.99645 |
| Normal weight | 144 (77.42) | 372 (77.99) | ||
| Overweight | 29 (15.59) | 71 (14.88) | ||
| Obesity | 11 (5.91) | 29 (6.08) |
| Gene Symbol | rs | Genotype | non-ADHD N (%) | ADHD N (%) | p-Value Genotypes | p-Value Alleles | OR (95% CI) | p-Value HWE |
|---|---|---|---|---|---|---|---|---|
| ADRA2A | rs553668 | AA | 5 (1.0) | 3 (1.3) | 0.655 | 0.365 | 1.169 (0.833–1.640) | 0.471 |
| AG | 99 (20.7) | 53 (23.5) | ||||||
| GG | 375 (78.3) | 170 (75.2) | ||||||
| ADRA2A | rs1800544 | CC | 317 (65.9) | 128 (56.6) | 0.056 | 0.018 | 0.725 (0.554–0.949) | 0.973 |
| CG | 146 (30.4) | 86 (38.1) | ||||||
| GG | 18 (3.7) | 12 (5.3) | ||||||
| AGO1 | rs595961 | AA | 346 (72.5) | 177 (78.3) | 0.258 | 0.113 | 1.318 (0.935–1.858) | 0.137 |
| AG | 125 (26.2) | 47 (20.8) | ||||||
| GG | 6 (1.3) | 2 (0.9) | ||||||
| ARL14 | rs1920644 | CC | 115 (23.9) | 49 (21.9) | 0.165 | 0.377 | 0.903 (0.721–1.131) | 0.939 |
| CT | 241 (50.1) | 110 (49.1) | ||||||
| TT | 125 (26.0) | 65 (29.0) | ||||||
| BDNF | rs6265 | CC | 341 (71.0) | 157 (69.5) | 0.728 | 0.841 | 0.968 (0.715–1.311) | 0.834 |
| CT | 125 (26.0) | 64 (28.3) | ||||||
| TT | 14 (3.0) | 5 (2.2) | ||||||
| BHMT | rs3733890 | AA | 40 (8.4) | 25 (16.2) | 0.337 | 0.147 | 1.194 (0.939–1.519) | 0.786 |
| AG | 201 (42.4) | 100 (44.6) | ||||||
| GG | 233 (49.2) | 99 (44.2) | ||||||
| COMT | rs4680 | AA | 136 (28.7) | 49 (22.6) | 0.242 | 0.152 | 0.847 (0.674–1.063) | 0.579 |
| AG | 226 (47.7) | 112 (51.6) | ||||||
| GG | 112 (23.6) | 56 (25.8) | ||||||
| DBH | rs2519152 | CC | 120 (25.2) | 60 (26.5) | 0.861 | 0.577 | 1.065 (0.851–1.332) | 0.124 |
| CT | 224 (46.9) | 107 (47.4) | ||||||
| TT | 133 (27.9) | 59 (26.1) | ||||||
| DRD2 | rs1124491 | AA | 16 (3.3) | 7 (3.1) | 0.986 | 0.920 | 0.984 (0.733–1.320) | 0.787 |
| AG | 138 (28.8) | 65 (28.8) | ||||||
| GG | 326 (67.9) | 154 (68.1) | ||||||
| DRD4 | rs1800955 | CC | 110 (23.3) | 42 (18.8) | 0.337 | 0.497 | 0.924 (0.737–1.160) | 0.149 |
| CT | 214 (45.2) | 112 (50.2) | ||||||
| TT | 149 (31.5) | 69 (30.9) | ||||||
| FTO | rs9939609 | AA | 101 (21.4) | 40 (19.3) | 0.158 | 0.527 | 1.077 (0.854–1.359) | 0.490 |
| AT | 216 (45.8) | 111 (53.6) | ||||||
| TT | 115 (32.8) | 56 (27.1) | ||||||
| HTR1B | rs6296 | CC | 267 (55.9) | 123 (54.4) | 0.612 | 1.000 | 1.000 (0.774–1.291) | 0.445 |
| CG | 173 (36.3) | 89 (39.4) | ||||||
| GG | 37 (7.8) | 14 (6.2) | ||||||
| HTR2C | rs518147 | CC | 114 (23.7) | 72 (32.0) | <0.000 | 0.603 | 1.064 (0.840–1.347) | <0.000 |
| CG | 92 (19.1) | 12 (5.3) | ||||||
| GG | 275 (57.2) | 141 (62.7) | ||||||
| HTR2C | rs3813929 | CC | 385 (80.5) | 182 (80.5) | 0.001 | 0.112 | 0.781 (0.576–1.060) | <0.000 |
| CT | 52 (10.9) | 10 (4.4) | ||||||
| TT | 41 (8.6) | 34 (15.1) | ||||||
| IPO11-HTR1A | rs10042956 | CC | 429 (89.2) | 197 (87.6) | 0.523 | 0.777 | 0.967 (0.772–1.212) | 0.111 |
| CT | 52 (10.8) | 28 (12.4) | ||||||
| KCNIP1 | rs1541665 | CC | 304 (63.7) | 164 (73.2) | 0.044 | 0.015 | 1.458 (1.074–1.979) | 0.751 |
| CT | 154 (32.3) | 54 (24.1) | ||||||
| TT | 19 (4.0) | 6 (2.7) | ||||||
| MC4R | rs17782313 | CC | 25 (5.3) | 12 (5.6) | 0.860 | 0.610 | 1.074 (0.812–1.421) | 0.096 |
| CT | 141 (30.5) | 69 (32.4) | ||||||
| TT | 297 (64.2) | 132 (62.0) | ||||||
| MTHFR | rs1801133 | AA | 45 (9.4) | 16 (7.1) | 0.548 | 0.314 | 0.880 (0.685–1.129) | 0.629 |
| AG | 194 (40.4) | 90 (39.8) | ||||||
| GG | 241 (50.2) | 120 (53.1) | ||||||
| MTHFR | rs1801131 | GG | 44 (9.2) | 37 (16.4) | 0.013 | 0.153 | 1.188 (0.937–1.505) | 0.226 |
| GT | 211 (43.9) | 84 (37.2) | ||||||
| TT | 225 (46.9) | 105 (46.4) | ||||||
| MTR | rs1805087 | AA | 287 (61.3) | 136 (62.4) | 0.830 | 1.000 | 1.008 (0.764–1.330) | 0.910 |
| AG | 161 (34.4) | 71 (32.6) | ||||||
| GG | 20 (4.3) | 11 (5.0) | ||||||
| PIK3CG | rs12667819 | AA | 100 (20.9) | 39 (17.3) | 0.515 | 0.442 | 0.915 (0.729–1.148) | 0.110 |
| AG | 215 (47.2) | 108 (48.0) | ||||||
| GG | 162 (33.9) | 78 (34.7) | ||||||
| RSPH3 | rs183882582 | AT | 15 (3.2) | 6 (2.7) | 0.769 | 0.777 | 0.867 (0.334–2.252) | 0.685 |
| TT | 459 (96.8) | 212 (97.3) | ||||||
| SLC1A3 | rs1049522 | AA | 220 (46.0) | 80 (35.4) | 0.028 | 0.022 | 0.763 (0.605–0.963) | 0.735 |
| AC | 201 (42.1) | 115 (50.9) | ||||||
| CC | 57 (11.9) | 31 (13.7) | ||||||
| SLC6A2 | rs5569 | AA | 40 (8.4) | 30 (13.3) | 0.064 | 0.029 | 1.297 (1.026–1.640) | 0.193 |
| AG | 220 (45.9) | 108 (47.8) | ||||||
| GG | 219 (45.7) | 88 (38.9) | ||||||
| SLC6A3 | rs463379 | CC | 26 (5.5) | 8 (3.5) | 0.322 | 0.841 | 1.026 (0.792–1.328) | 0.054 |
| CG | 184 (38.6) | 98 (43.4) | ||||||
| GG | 267 (55.9) | 120 (53.1) | ||||||
| SLC6A4 | rs6354 | GG | 18 (3.8) | 7 (3.1) | 0.188 | 0.310 | 1.164 (0.868–1.560) | 0.193 |
| GT | 117 (34.7) | 70 (31.3.2) | ||||||
| TT | 338 (71.5) | 147 (65.6) | ||||||
| SLC19A1 | rs1051266 | CC | 147 (30.6) | 59 (26.2) | 0.484 | 0.383 | 0.905 (0.723–1.132) | 0.530 |
| CT | 228 (47.4) | 115 (51.1) | ||||||
| TT | 106 (22.0) | 51 (22.7) | ||||||
| SNAP25 | rs1051312 | CC | 28 (5.9) | 11 (4.9) | 0.847 | 0.887 | 0.980 (0.752–1.277) | 0.997 |
| CT | 170 (35.6) | 83 (36.9) | ||||||
| TT | 279 (58.5) | 131 (58.2) |
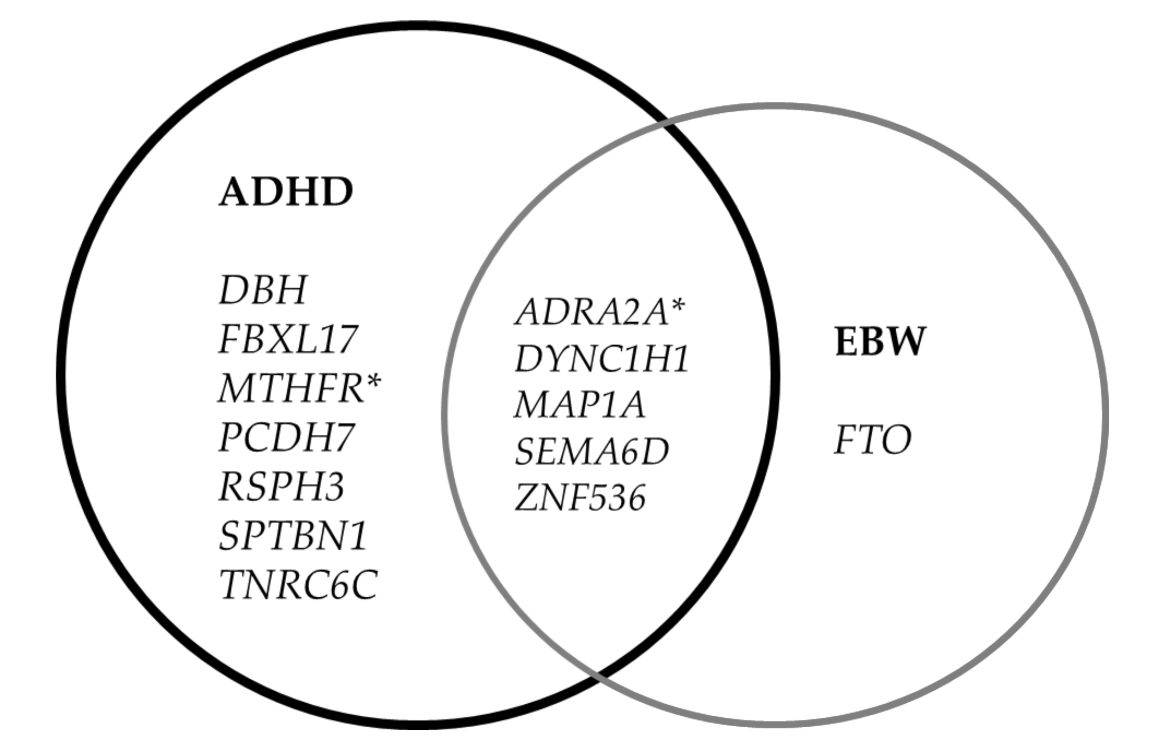
| Gene Symbol | HGVSg | ALT | Consequence | Amino Acids | Existing VARIATION | Impact | PolyPhen | ADHD with/without QV for Gene | non-ADHD with/without QV for Gene | P-Value Fisher Exact Test |
|---|---|---|---|---|---|---|---|---|---|---|
| (A) Detected Gene Variants in ADHD vs. Not Detect in Non-ADHD | ||||||||||
| ADRA2A | chr10:g.111078090G>C | C | MV | E/Q | rs753177273,COSV54528494 | M | benign | 2/47 | 0/46 | 0.263 |
| ADRA2A | chr10:g.111078097C>A | A * | MV | P/Q | - | M | probably damaging | |||
| DBH | chr9:g.133656524G>C | C * | MV, SRV | G/A | COSV67548769 | M | probably damaging | 1/48 | 0/46 | 0.516 |
| DYNC1H1 | chr14:g.102016033G>A | A | MV | A/T | rs766837403,COSV64136626 | M | benign | 1/48 | 0/46 | 0.516 |
| FBXL17 | chr5:g.108224144T>C | C | MV | K/E | rs141165823 | M | benign | 5/44 | 0/46 | 0.033 |
| FBXL17 | chr5:g.108380944C>A | A * | MV | A/S | - | M | benign | |||
| FBXL17 | chr5:g.108380956C>A | A * | MV | G/C | - | M | possibly damaging | |||
| FBXL17 | chr5:g.108381031_108381033dup | ACCG * | II | G/GG | rs906102490 | M | - | |||
| MAP1A | chr15:g.43527328A>C | C * | MV | E/A | - | M | probably damaging | 1/48 | 0/46 | 0.516 |
| MTHFR | chr1:g.11794399_11794400insAAA | CAAA * | II | -/F | - | M | - | 1/48 | 0/46 | 0.516 |
| PCDH7 | chr4:g.30721475G>T | T | MV | C/F | rs757643187 | M | benign | 1/48 | 0/46 | 0.516 |
| RSPH3 | chr6:g.158999732A>C | C * | MV | I/R | rs1217221445 | M | benign | 1/48 | 0/46 | 0.516 |
| SCN2A | chr2:g.165307860G>A | A * | MV | M/I | COSV51836794 | M | probably damaging | 1/48 | 0/46 | 0.516 |
| SEMA6D | chr15:g.47771337G>A | A | MV | R/Q | rs766660850 | M | probably damaging | 2/47 | 0/46 | 0.263 |
| SEMA6D | chr15:g.47771766T>C | C * | MV | L/P | rs540588380 | M | possibly damaging | |||
| SPTBN1 | chr2:g.54599190C>G | G * | MV | R/G | rs915376910,COSV61693274 | M | probably damaging | 1/48 | 0/46 | 0.516 |
| TNRC6C | chr17:g.78091580C>T | T * | MV | P/S | rs1302015314 | M | benign | 1/48 | 0/46 | 0.516 |
| ZNF536 | chr19:g.30548324_30548326del | A * | ID | RA/T | - | M | - | 2/47 | 0/46 | 0.263 |
| ZNF536 | chr19:g.30549475A>C | C * | MV | T/P | rs1257346923 | M | benign | |||
| (B) Detected Gene Variants in ADHD with EBW vs. Not Detect in Non-ADHD with NBW | ||||||||||
| ADRA2A | chr10:g.111078090G>C | C | MV | E/Q | rs753177273,COSV54528494 | M | benign | 1/13 | 0/44 | 0.241 |
| DYNC1H1 | chr14:g.102016033G>A | A | MV | A/T | rs766837403,COSV64136626 | M | benign | 1/13 | 0/44 | 0.241 |
| FTO | chr16:g.53826140G>A | A | MV | A/T | rs79206939 | M | benign | 1/13 | 0/44 | 0.241 |
| MAP1A | chr15:g.43527328A>C | C * | MV | E/A | - | M | probably damaging | 1/13 | 0/44 | 0.241 |
| SEMA6D | chr15:g.47771337G>A | A | MV | R/Q | rs766660850 | M | probably damaging | 1/13 | 0/44 | 0.241 |
| ZNF536 | chr19:g.30549475A>C | C * | MV | T/P | rs1257346923 | M | benign | 1/13 | 0/44 | 0.241 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dmitrzak-Weglarz, M.; Paszynska, E.; Bilska, K.; Szczesniewska, P.; Bryl, E.; Duda, J.; Dutkiewicz, A.; Tyszkiewicz-Nwafor, M.; Czerski, P.; Hanc, T.; et al. Common and Unique Genetic Background between Attention-Deficit/Hyperactivity Disorder and Excessive Body Weight. Genes 2021, 12, 1407. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12091407
Dmitrzak-Weglarz M, Paszynska E, Bilska K, Szczesniewska P, Bryl E, Duda J, Dutkiewicz A, Tyszkiewicz-Nwafor M, Czerski P, Hanc T, et al. Common and Unique Genetic Background between Attention-Deficit/Hyperactivity Disorder and Excessive Body Weight. Genes. 2021; 12(9):1407. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12091407
Chicago/Turabian StyleDmitrzak-Weglarz, Monika, Elzbieta Paszynska, Karolina Bilska, Paula Szczesniewska, Ewa Bryl, Joanna Duda, Agata Dutkiewicz, Marta Tyszkiewicz-Nwafor, Piotr Czerski, Tomasz Hanc, and et al. 2021. "Common and Unique Genetic Background between Attention-Deficit/Hyperactivity Disorder and Excessive Body Weight" Genes 12, no. 9: 1407. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12091407