
Differential Expression of Kisspeptin System and Kisspeptin Receptor Trafficking during Spermatozoa Transit in the Epididymis


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Collection
2.2. Collection of Epididymal Spermatozoa, Epididymal Tissue and Fluid
2.3. Total Protein Extraction
2.4. Western Blot
2.5. Immunohistochemistry for Kiss1 and Kiss1R in Rat Seminiferous Tubules and Epididymis
2.6. Immunofluorescence Analysis
2.7. Dot Blot Analysis and Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Statistics
3. Results
3.1. Kiss1 and Kiss1R Protein in Spermatozoa
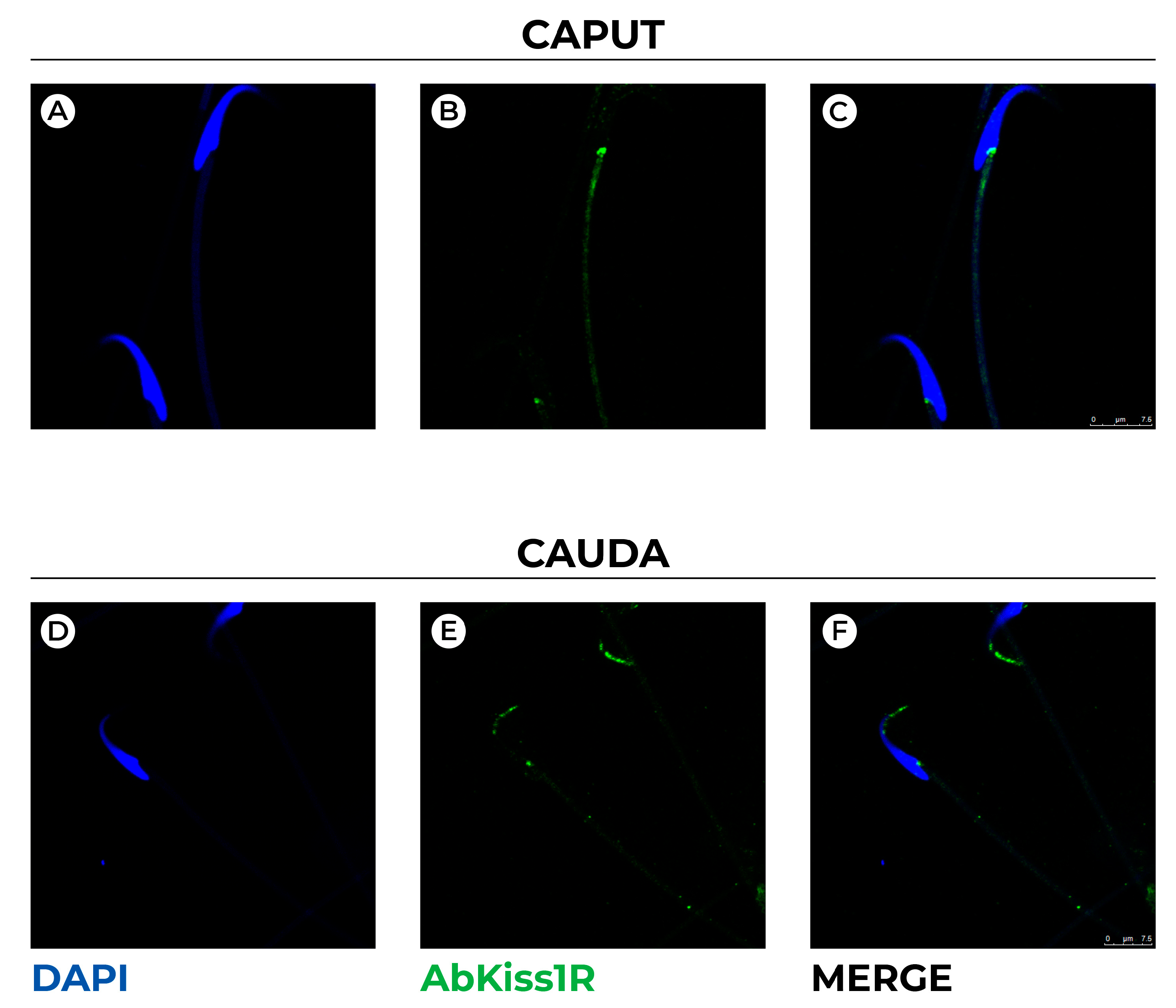
3.2. Expression and Localization of Kiss1 and Kiss1R Protein in the Epididymis
3.3. Dot Blot and ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pierantoni, R.; Cobellis, G.; Meccariello, R.; Fasano, S. Evolutionary aspects of cellular communication in the vertebrate hypothalamo-hypophysio-gonadal axis. Int. Rev. Cytol. 2002, 218, 69–141. [Google Scholar]
- Oakley, A.E.; Clifton, D.K.; Steiner, R.A. Kisspeptin signaling in the brain. Endocr. Rev. 2009, 30, 713–743. [Google Scholar] [CrossRef] [PubMed]
- de Roux, N.; Genin, E.; Carel, J.C.; Matsuda, F.; Chaussain, J.L.; Milgrom, E. Hypogonadotropic Hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funes, S.; Hedrick, J.A.; Vassileva, G.; Markowitz, L.; Abbondanzo, S.; Golovko, A.; Yang, S.; Monsma, F.J.; Gustafson, E.J. The KiSS-1 receptor GPR54 is essential for the development of the murine reproductive system. Biochem. Biophys. Res. Commun. 2003, 12, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S., Jr.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G.; et al. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [Green Version]
- de Tassigny, X.D.A.; Fagg, L.A.; Dixon, J.P.; Day, K.; Leitch, H.G.; Hendrick, A.G.; Zahn, S.; Franceschini, I.; Caraty, A.; Carlton, M.B.L.; et al. Hypogonadotropic Hypogonadism in mice lacking a functional Kiss1. Gene. Proc. Natl. Acad. Sci. USA 2007, 104, 10714–10719. [Google Scholar] [CrossRef] [Green Version]
- Teles, M.G.; Silveira, L.F.; Tusset, C.; Latronico, A.C. New genetic factors implicated in human GnRH-dependent precocious puberty: The role of kisspeptin system. Mol. Cell. Endocrinol. 2011, 346, 84–90. [Google Scholar] [CrossRef]
- Chianese, R.; Cobellis, G.; Chioccarelli, T.; Ciaramella, V.; Migliaccio, M.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptins, estrogens and male fertility. Curr. Med. Chem. 2016, 23, 4070–4091. [Google Scholar] [CrossRef]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and Reproduction: Physiological Roles and Regulatory Mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Colledge, W.H.; Fasano, S.; Meccariello, R. Editorial: The multiple facets of Kisspeptin activity in biological systems. Front. Endocrinol. 2018, 9, 727. [Google Scholar] [CrossRef]
- Yeo, S.-H.; Colledge, W.H. The role of Kiss1 neurons as integrators of endocrine, metabolic, and environmental factors in the hypothalamic–pituitary–gonadal axis. Front. Endocrinol. 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Motti, M.L.; Meccariello, R. Minireview: The epigenetic modulation of KISS1 in reproduction and cancer. Int. J. Environ. Res. Public Health 2019, 1, 2607. [Google Scholar] [CrossRef] [Green Version]
- Santoro, A.; Chianese, R.; Troisi, J.; Richards, S.; Nori, S.L.; Fasano, S.; Guida, M.; Plunk, E.; Viggiano, A.; Pierantoni, R.; et al. Neuro-toxic and Reproductive Effects of BPA. Curr. Neuropharmacol. 2019, 17, 1109–1132. [Google Scholar] [CrossRef] [PubMed]
- Meccariello, R.; Chianese, R.; Chioccarelli, T.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Cobellis, G. Intra-testicular signals regulate germ cell progression and production of qualitatively mature spermatozoa in vertebrates. Front. Endocrinol. 2014, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Thaventhiran, T.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Kisspeptin and testicular function-is it necessary? Int. J. Mol. Sci. 2020, 21, 2958. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, R.; Fasano, S.; Pierantoni, R. Kisspeptins, new local modulators of male reproduction: A comparative overview. Gen. Comp. Endocrinol. 2020, 299, 113618. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.C.; Wang, J.Y.; Lee, Y.J.; Jong, D.S.; Tsui, K.H.; Chiu, C.H. Kisspeptin modulates fertilization capacity of mouse spermatozoa. Reproduction 2014, 147, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhao, Y.; Si, W.; Jiang, X.; Wu, J.; Na, R.; Han, Y.; Li, K.; Yang, L.; Guangxin, E.; et al. Temporal expression of the KISS1/GPR54 system in goats’ testes and epididymides and its spatial expression in pubertal goats. Theriogenology 2020, 152, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Wahab, F.; Atika, B.; Shahab, M.; Behr, R. Kisspeptin signalling in the physiology and pathophysiology of the urogenital system. Nat. Rev. Urol. 2016, 13, 21–32. [Google Scholar] [CrossRef]
- León, S.; Barroso, A.; Vázquez, M.J.; García-Galiano, D.; Manfredi-Lozano, M.; Ruiz-Pino, F.; Heras, V.; Romero-Ruiz, A.; Roa, J.; Schutz, G.; et al. Direct actions of kisspeptins on GnRH neurons permit attainment of fertility but are insufficient to fully preserve gonadotropic axis activity. Sci. Rep. 2016, 6, 19206. [Google Scholar] [CrossRef]
- Samir, H.; Nagaoka, K.; Watanabe, G. Effect of kisspeptin antagonist on goat in vitro Leydig cell steroidogenesis. Theriogenology 2018, 121, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Petrucci, L.; Maranesi, M.; Verini Supplizi, A.; Dall’Aglio, C.; Mandara, M.T.; Quassinti, L.; Bramucci, M.; Miano, A.; Gobbetti, A.; Catone, G.; et al. Kisspeptin/GnRH1 system in Leydig cells of horse (Equus caballus): Presence and function. Theriogenology 2020, 152, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin receptor, GPR54, as a candidate for the regulation of testicular activity in the frog, Rana esculenta. Biol. Reprod. 2013, 88, 73. [Google Scholar] [CrossRef]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin drives germ cell progression in the anuran amphibian Pelophylax esculentus: A study carried out in ex vivo testes. Gen. Comp. Endocrinol. 2015, 211, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin regulates steroidogenesis and spermiation in the anuran amphibian Pelophylax esculentus testis. Reproduction 2017, 154, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Mei, H.; Doran, J.; Kyle, V.; Yeo, S.H.; Colledge, W.H. Does kisspeptin signalling have a role in the testes? Front. Endocrinol. 2013, 4, 198. [Google Scholar] [CrossRef]
- Toolee, H.; Rastegar, T.; Solhjoo, S.; Mortezaee, K.; Mohammadipour, M.; Kashani, I.R.; Akbari, M. Roles for Kisspeptin in proliferation and differentiation of spermatogonial cells isolated from mice offspring when the cells are cocultured with somatic cells. J. Cell. Biochem. 2019, 120, 5042–5054. [Google Scholar] [CrossRef]
- Hussain, A.; Nabi, W.; Zubair, H.; Bano, R.; Ahmed, H.; Andrabi, S.M.H.; Shahab, M. Immunocytochemical detection of kisspeptin receptor and its association with motility of buffalo bull (Bubalus bubalis) spermatozoa. Pak. Vet. J. 2020, 40, 244–248. [Google Scholar] [CrossRef]
- Pinto, F.M.; Cejudo-Román, A.; Ravina, C.G.; Fernández-Sánchez, M.; Martín-Lozano, D.; Illanes, M.; Tena-Sempere, M.; Candenas, M.L. Characterization of the kisspeptin system in human spermatozoa. Int. J. Androl. 2012, 35, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Gloria, A.; Contri, A.; Mele, E.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog. Int. J. Mol. Sci. 2021, 22, 10120. [Google Scholar] [CrossRef]
- Zou, P.; Wang, X.; Chen, Q.; Yang, H.; Zhou, N.; Sun, L.; Chen, H.; Liu, J.; Ao, L.; Cui, Z. Kisspeptin Protein in Seminal Plasma Is Positively Associated with Semen Quality: Results from the MARHCS Study in Chongqing, China. Biomed. Res. Int. 2019, 2019, 5129263. [Google Scholar] [CrossRef]
- Xu, L.; Xue, H.; Li, S.; Xu, J.; Chen, L. Seasonal differential expression of KiSS-1/GPR54 in the striped hamsters (Cricetulus barabensis) among different tissues. Integr. Zool. 2017, 12, 260–268. [Google Scholar] [CrossRef]
- Lowry, O.; Rosenbrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Berruti, G.; Martegani, E. The deubiquitinating enzyme mUBPy interacts with the sperm specific molecular chaperone MSJ-1: The relation with the proteasome, acrosome and centrosome in mouse male germ cells. Biol. Reprod. 2005, 72, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, R.; Cobellis, G.; Berruti, G.; Junier, M.-P.; Ceriani, M.; Boilée, S.; Pierantoni, R.; Fasano, S. Mouse Sperm Cell-Specific DnaJ First Homologue: An Evolutionarily Conserved Protein for Spermiogenesis. Biol. Reprod. 2002, 66, 1328–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschini, I.; Yeo, S.H.; Beltramo, M.; Desroziers, E.; Okamura, H.; Herbison, A.E.; Caraty, A. Immunohistochemical evidence for the presence of various kisspeptin isoforms in the mammalian brain. J. Neuroendocrinol. 2013, 25, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Berruti, G.; Martegani, E. MSJ-1, a Mouse Testis-Specific DnaJ Protein, Is Highly Expressed in Haploid Male Germ Cells and Interacts with the Testis-Specific Heat Shock Protein Hsp70-2. Biol. Reprod. 2001, 65, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Tena-Sempere, M. Kisspeptin signaling in the brain: Recent developments and future challenges. Mol. Cell. Endocrinol. 2010, 314, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, S.; Krishna, A.; Sridaran, R.; Tsutsui, K. Localization of gonadotropin-releasing hormone (GnRH), gonadotropin-inhibitory hormone (GnIH), kisspeptin and GnRH receptor and their possible roles in testicular activities from birth to senescence in mice. J. Exp. Zool. A Ecol. Genet. Physiol. 2012, 317, 630–644. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.R.; Shahab, M. Effect of kisspeptin challenge on testosterone and inhibin secretion from in vitro testicular tissue of adult male rhesus monkey (Macaca mulatta). Andrologia 2017, 49, e12590. [Google Scholar] [CrossRef]
- Salehi, S.; Adeshina, I.; Chen, H.; Zirkin, B.R.; Hussain, M.A.; Wondisford, F.; Wolfe, A.; Radovick, S. Developmental and endocrine regulation of kisspeptin expression in mouse Leydig cells. Endocrinology 2015, 156, 1514–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samir, H.; Nagaoka, K.; Karen, A.; Ahmed, E.; El Sayed, M.; Watanabe, G. Investigation the mRNA expression of KISS1 and localization of kisspeptin in the testes of Shiba goats and its relationship with the puberty and steriodogenic enzymes. Small Rum. Res. 2015, 133, 1–6. [Google Scholar] [CrossRef]
- Irfan, S.; Ehmcke, J.; Shahab, M.; Wistuba, J.; Schlatt, S. Immunocytochemical localization of kisspeptin and kisspeptin receptor in the primate testis. J. Med. Primatol. 2016, 45, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, Q.; Liu, X.; Xu, Y.; Song, X.; Shi, B. Molecular characterization of kiss2 and differential regulation of reproduction-related genes by sex steroids in the hypothalamus of half-smooth tongue sole (Cynoglossus semilaevis). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 213, 46–55. [Google Scholar] [CrossRef]
- Breton, S.; Ruan, Y.C.; Park, Y.-J.; Kim, B. Regulation of epithelial function, differentiation, and remodeling in the epididymis. Asian J. Androl. 2016, 18, 3–9. [Google Scholar] [CrossRef]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: Their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef] [Green Version]
- Protopapas, N.; Hamilton, L.; Warkentin, R.; Xu, W.; Sutovsky, P.; Oko, R. The perforatorium and postacrosomal sheath of rat spermatozoa share common developmental origins and protein constituents. Biol. Reprod. 2019, 100, 1461–1472. [Google Scholar] [CrossRef]
- Hamilton, L.E.; Lion, M.; Aguila, L.; Suzuki, J.; Acteau, G.; Protopapas, N.; Xu, W.; Sutovsky, P.; Baker, M.; Oko, R. Core Histones Are Constituents of the Perinuclear Theca of Murid Spermatozoa: An Assessment of Their Synthesis and Assembly during Spermiogenesis and Function after Gametic Fusion. Int. J. Mol. Sci. 2021, 22, 8119. [Google Scholar] [CrossRef]
- Costello, S.; Michelangeli, F.; Nash, K.; Lefievre, L.; Morris, J.; Machado-Oliveira, G.; Barratt, C.; Kirkman-Brown, J.; Publicover, S. Ca2+-stores in sperm: Their identities and functions. Reproduction 2009, 138, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Jin, W.; Peng, Y.; He, Z.; Wei, L.; Li, S.; Wang, X.; Change, M.; Wang, R. In vivo and vitro characterization of the effects of kisspeptin-13, endogenous ligands for GPR54, on mouse gastrointestinal motility. Eur. J. Pharmacol. 2017, 794, 216–223. [Google Scholar] [CrossRef]
- Cho, S.G.; Li, D.; Tan, K.; Siwko, S.K.; Liu, M. KiSS1 and its G-protein-coupled receptor GPR54 in cancer development and metastasis. Cancer Metas. Rev. 2012, 31, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Oktar, S.; Yonden, Z.; Ozturk, O.H.; Yilmaz, B. Direct and indirect effects of kisspeptin on liver oxidant and antioxidant systems in young male rats. Cell. Biochem. Funct. 2010, 28, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Aslan, M.; Erkanli Senturk, G.; Akkaya, H.; Sahin, S.; Yılmaz, B. The effect of oxytocin and Kisspeptin-10 in ovary and uterus of ischemia-reperfusion injured rats. Taiwan. J. Obstetr. Gynecol. 2017, 56, 456–462. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mele, E.; D’Auria, R.; Scafuro, M.; Marino, M.; Fasano, S.; Viggiano, A.; Pierantoni, R.; Santoro, A.; Meccariello, R. Differential Expression of Kisspeptin System and Kisspeptin Receptor Trafficking during Spermatozoa Transit in the Epididymis. Genes 2022, 13, 295. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020295
Mele E, D’Auria R, Scafuro M, Marino M, Fasano S, Viggiano A, Pierantoni R, Santoro A, Meccariello R. Differential Expression of Kisspeptin System and Kisspeptin Receptor Trafficking during Spermatozoa Transit in the Epididymis. Genes. 2022; 13(2):295. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020295
Chicago/Turabian StyleMele, Elena, Raffaella D’Auria, Marika Scafuro, Marianna Marino, Silvia Fasano, Andrea Viggiano, Riccardo Pierantoni, Antonietta Santoro, and Rosaria Meccariello. 2022. "Differential Expression of Kisspeptin System and Kisspeptin Receptor Trafficking during Spermatozoa Transit in the Epididymis" Genes 13, no. 2: 295. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13020295