
Preliminary Interpretations of Epigenetic Profiling of Cord Blood in Preeclampsia


Abstract
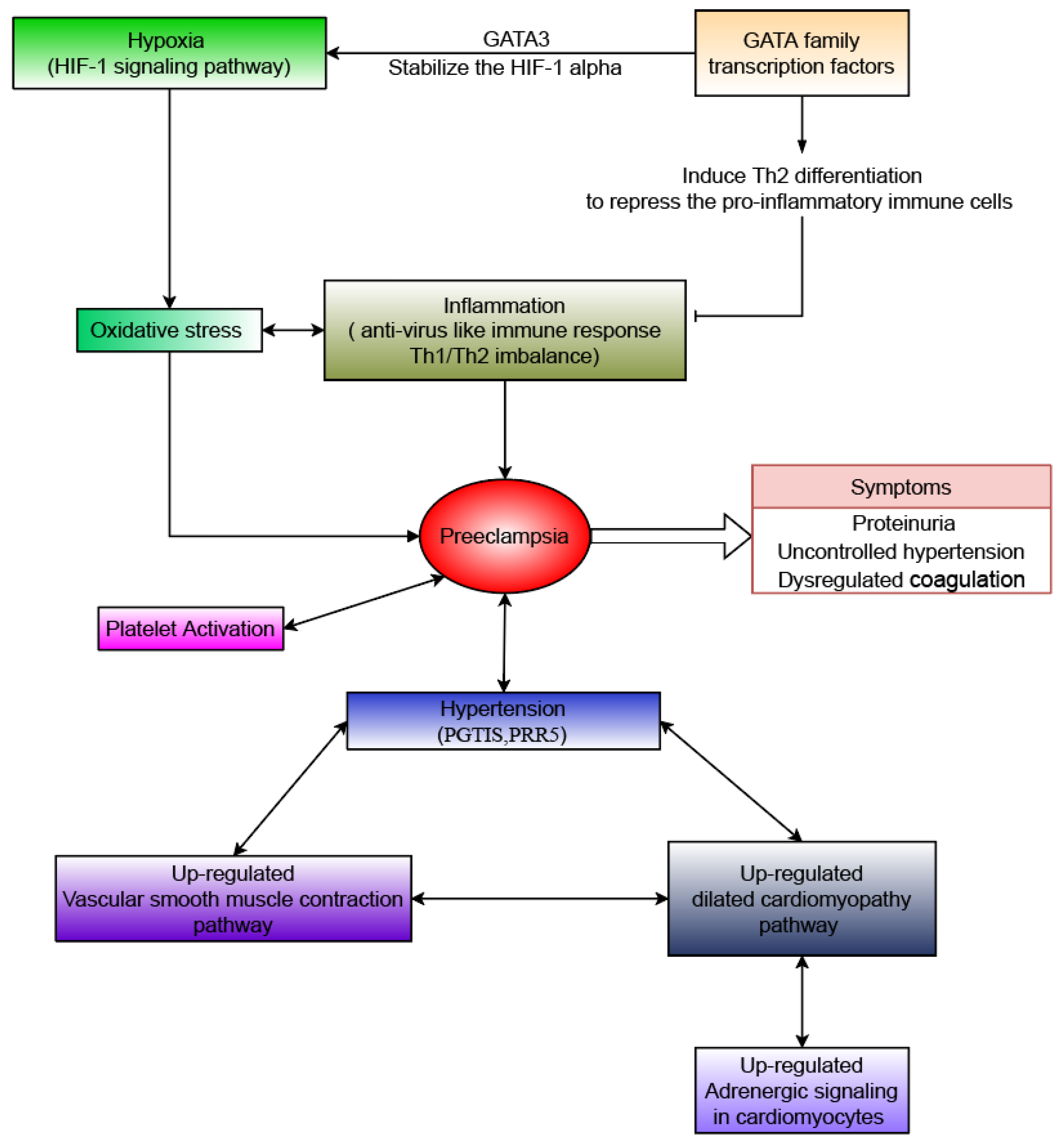
:1. Introduction
2. Materials and Methods
2.1. Patient Recruitment Criteria
2.2. Sample Preparation for ATAC-seq
2.3. Peak Calling
2.4. Peak Annotation
2.5. Peak Visualization
2.6. Pathway Enrichment Analysis
3. Results
3.1. Case Presentation
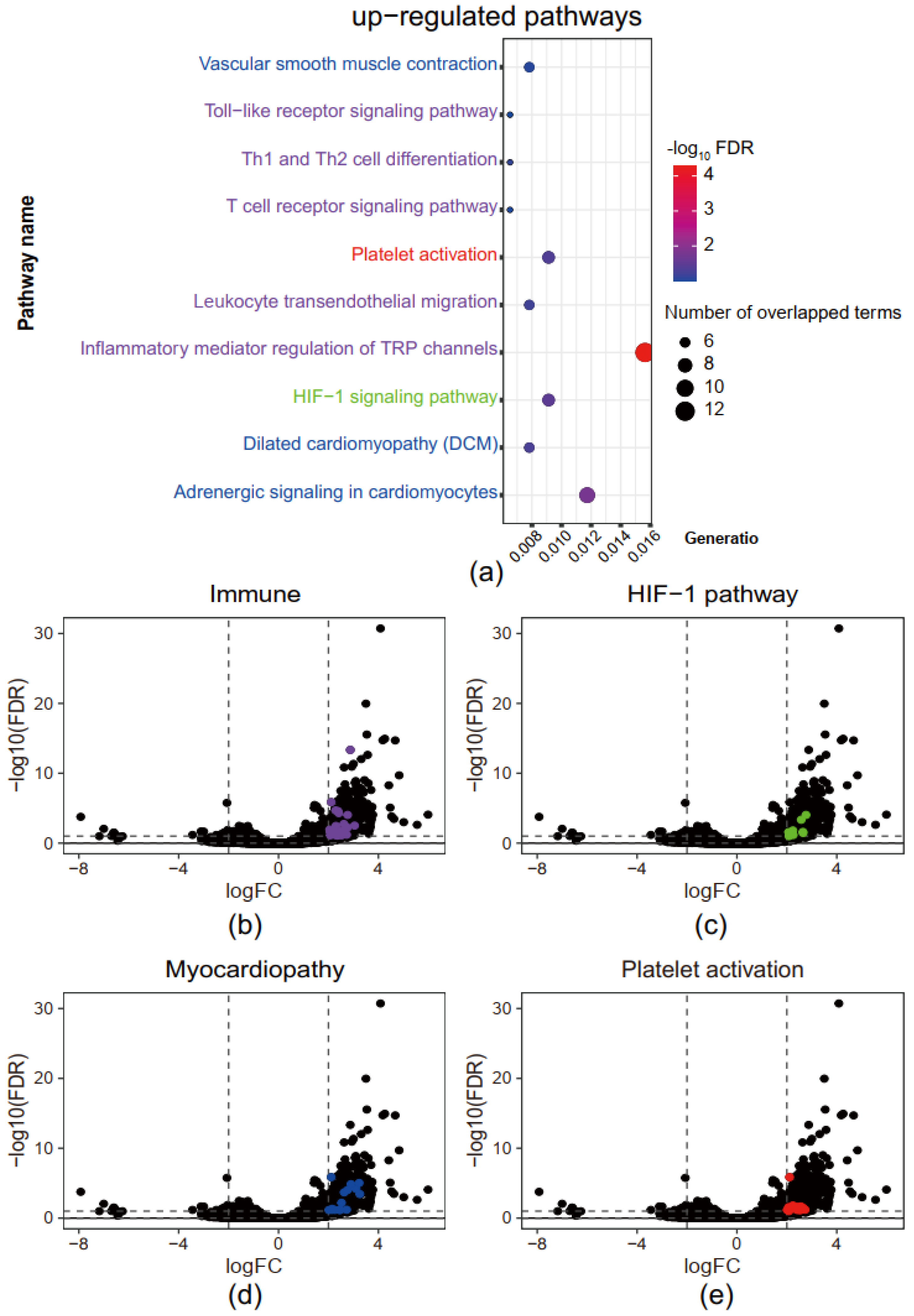
3.2. Immune, Myocardiopathy, and Hypoxia Response-Related Pathways Are Up-Regulated in Superimposed PE
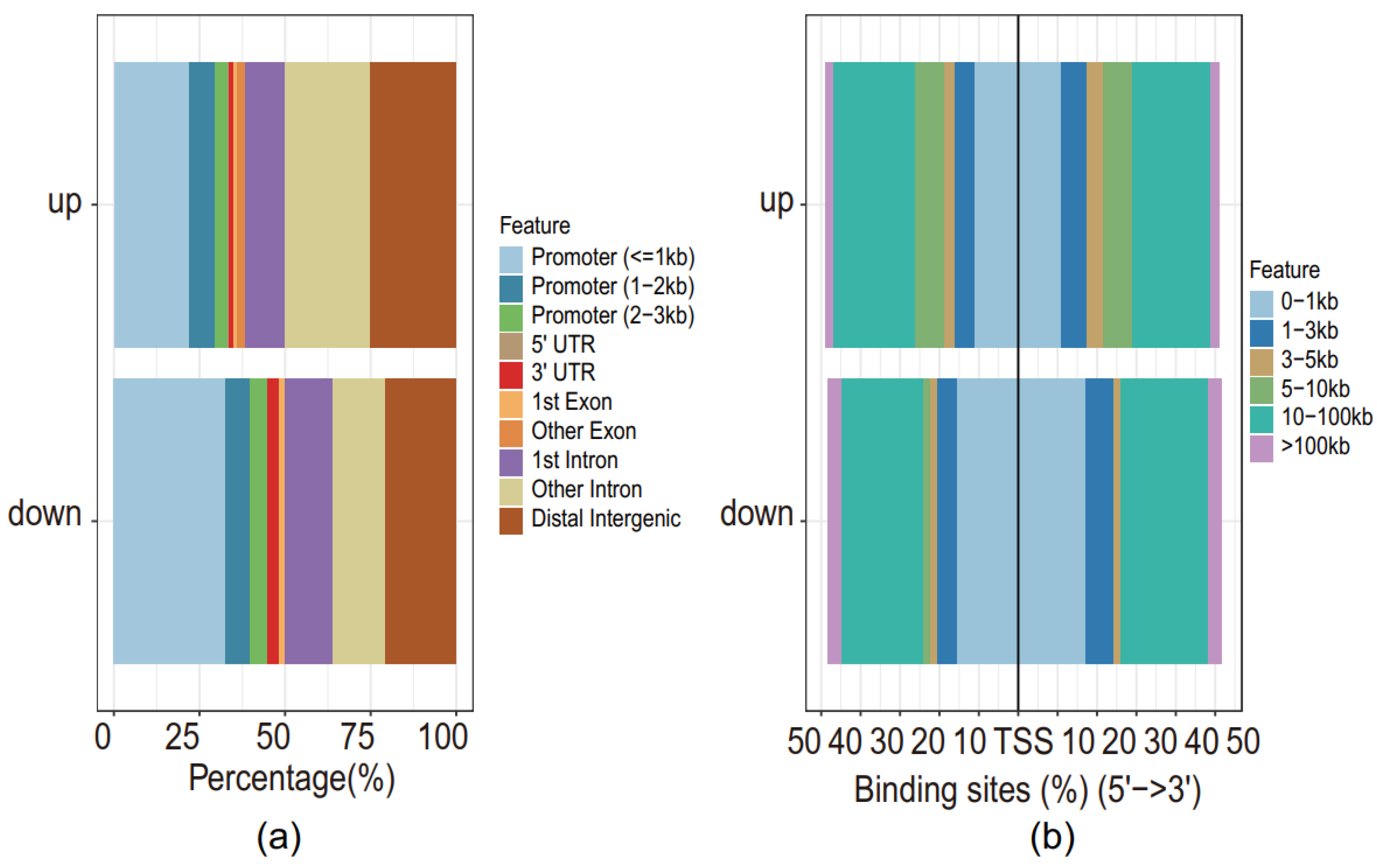
3.3. Higher Chromatin Accessibility across TSS of Genes of Differentially Expressed Peaks in Superimposed PE
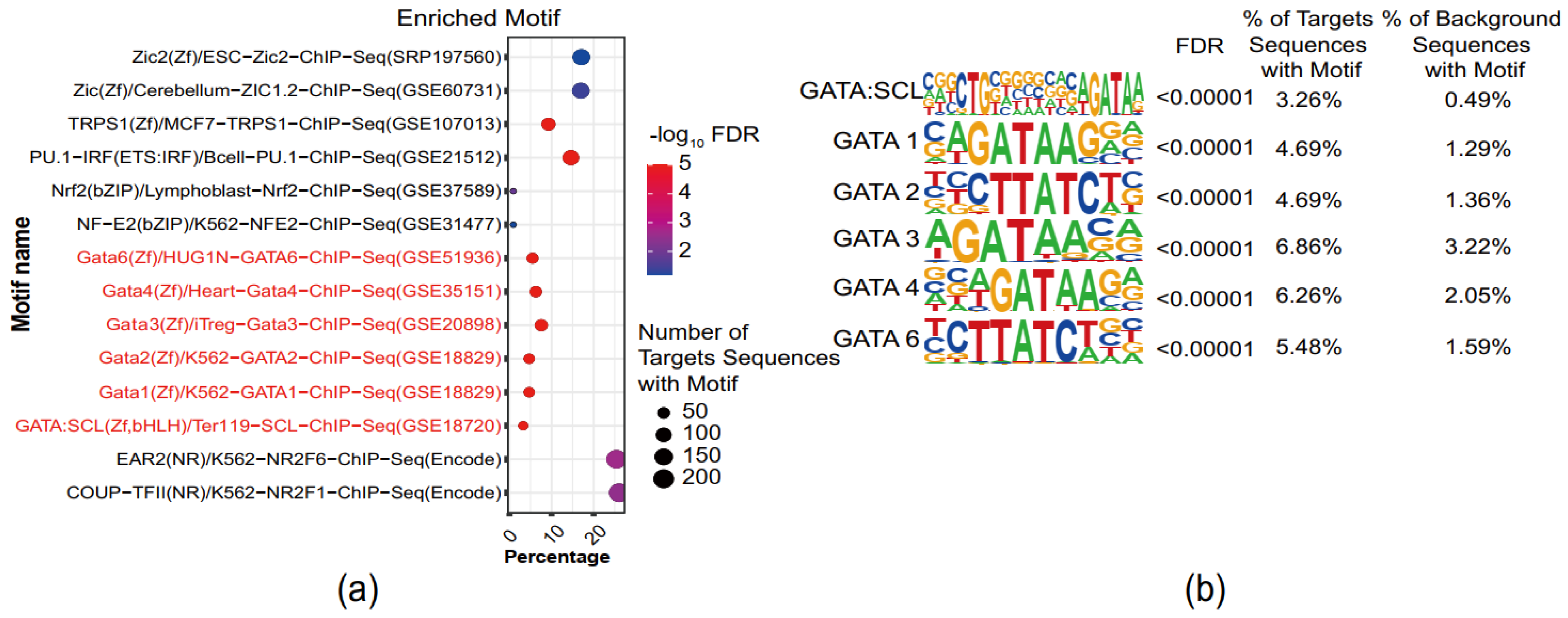
3.4. GATA-Binding Motifs Are Enriched in Superimposed PE
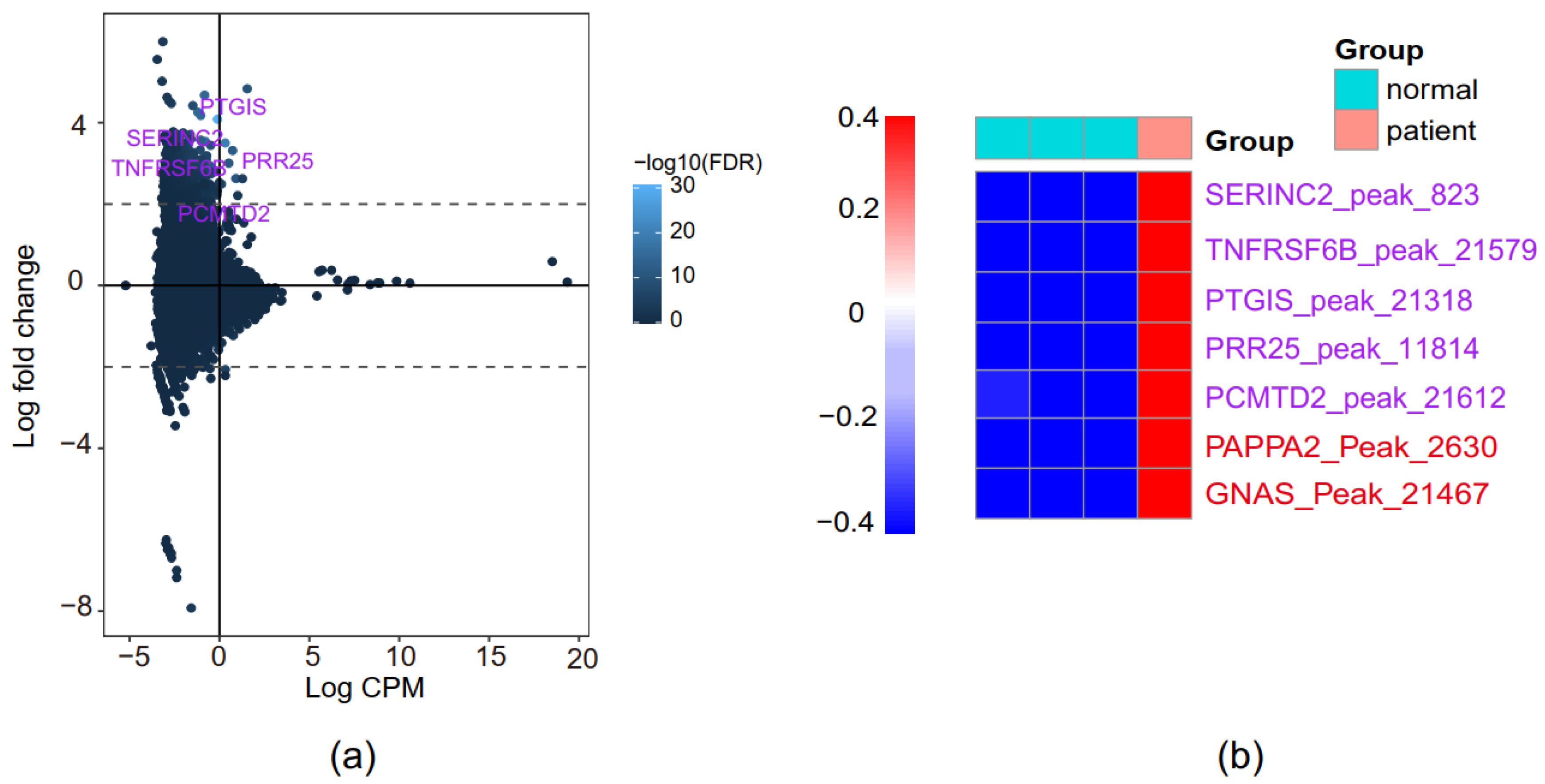
3.5. Identification of Novel PE-Associated Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mol, B.W.; Roberts, C.T.; Thangaratinam, S.; Magee, L.A.; De Groot, C.J.; Hofmeyr, G.J. Pre-eclampsia. Lancet 2016, 387, 999–1011. [Google Scholar] [CrossRef]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy: ISSHP Classification, Diagnosis, and Management Recommen-dations for International Practice. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, M.S.; Davey, M.-A.; Mol, B.W.; Rolnik, D.L. The evolution of the diagnostic criteria of preeclampsia-eclampsia. Am. J. Obstet. Gynecol. 2022, 226, S835–S843. [Google Scholar] [CrossRef] [PubMed]
- Socha, M.; Malinowski, B.; Puk, O.; Dubiel, M.; Wiciński, M. The NLRP3 Inflammasome Role in the Pathogenesis of Pregnancy Induced Hypertension and Preeclampsia. Cells 2020, 9, 1642. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Kondoh, E.; Chigusa, Y.; Kawamura, Y.; Mogami, H.; Takeda, S.; Horie, A.; Baba, T.; Matsumura, N.; Mandai, M.; et al. Metabolomic Profiles of Placenta in Preeclampsia. Hypertension 2019, 73, 671–679. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists. Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. Obstet. Gynecol. 2013, 122, 1122–1131. [Google Scholar]
- Seshadri Reddy, V.; Duggina, P.; Vedhantam, M.; Manne, M.; Varma, N.; Nagaram, S. Maternal serum and fetal cord-blood ische-mia-modified albumin concentrations in normal pregnancy and preeclampsia: A systematic review and meta-analysis. J. Matern. Fetal Neonatal Med. 2018, 31, 3255–3266. [Google Scholar] [CrossRef]
- Vargas-Rojas, M.I.; Solleiro-Villavicencio, H.; Soto-Vega, E. Th1, Th2, Th17 and Treg levels in umbilical cord blood in preeclamp-sia. J. Matern. Fetal Neonatal Med. 2016, 29, 1642–1645. [Google Scholar] [CrossRef]
- Gumina, D.L.; Black, C.P.; Balasubramaniam, V.; Winn, V.D.; Baker, C.D. Umbilical Cord Blood Circulating Progenitor Cells and Endothelial Colony-Forming Cells Are Decreased in Preeclampsia. Reprod. Sci. 2017, 24, 1088–1096. [Google Scholar] [CrossRef]
- Sukenik-Halevy, R.; Amiel, A.; Kidron, D.; Liberman, M.; Ganor-Paz, Y.; Biron-Shental, T. Telomere homeostasis in trophoblasts and in cord blood cells from pregnancies complicated with preeclampsia. Am. J. Obstet. Gynecol. 2016, 214, 283.e1–283.e7. [Google Scholar] [CrossRef]
- Wojczakowski, W.; Kimber-Trojnar, Ż.; Dziwisz, F.; Słodzińska, M.; Słodziński, H.; Leszczyńska-Gorzelak, B. Preeclampsia and Cardiovascular Risk for Offspring. J. Clin. Med. 2021, 10, 3154. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.; McCarthy, C.M.; O’Keeffe, G.W. Preeclampsia and Neurodevelopmental Outcomes: Potential Pathogenic Roles for Inflammation and Oxidative Stress? Mol. Neurobiol. 2021, 58, 2734–2756. [Google Scholar] [CrossRef] [PubMed]
- Holden, C.; Moses, N.; Fox, M.; Glose, G.; Vaughn, B.C.; Marshall-Taylor, S. Collaborating to Address Infant Mortality: Lessons Learned from the Brownsville Action Community for Health Equality. Prog. Community Health Partnersh. Res. Educ. Action 2011, 5, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Teng, R.J.; Wu, T.J.; Sharma, R.; Garrison, R.D.; Hudak, M.L. Early neonatal hypotension in premature infants born to preeclamptic mothers. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2006, 26, 471–475. [Google Scholar] [CrossRef]
- Sitras, V.; Paulssen, R.; Grønaas, H.; Leirvik, J.; Hanssen, T.; Vårtun, Å.; Acharya, G. Differential Placental Gene Expression in Severe Preeclampsia. Placenta 2009, 30, 424–433. [Google Scholar] [CrossRef]
- Catarino, C.; Santos-Silva, A.; Belo, L.; Rocha-Pereira, P.; Rocha, S.; Patrício, B.; Quintanilha, A.; Rebelo, I. Inflammatory Disturbances in Preeclampsia: Relationship between Maternal and Umbilical Cord Blood. J. Pregnancy 2012, 2012, 684384. [Google Scholar] [CrossRef]
- Rasmussen, M.; Reddy, M.; Nolan, R.; Camunas-Soler, J.; Khodursky, A.; Scheller, N.M.; Cantonwine, D.E.; Engelbrechtsen, L.; Mi, J.D.; Dutta, A.; et al. RNA profiles reveal signatures of future health and disease in pregnancy. Nature 2022, 601, 422–427. [Google Scholar] [CrossRef]
- Kamrani, A.; Alipourfard, I.; Ahmadi-Khiavi, H.; Yousefi, M.; Rostamzadeh, D.; Izadi, M.; Ahmadi, M. The role of epigenetic changes in preeclampsia. BioFactors 2019, 45, 712–724. [Google Scholar] [CrossRef]
- Apicella, C.; Ruano, C.S.M.; Méhats, C.; Miralles, F.; Vaiman, D. The Role of Epigenetics in Placental Development and the Etiology of Preeclampsia. Int. J. Mol. Sci. 2019, 20, 2837. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, J. Abnormal fetal-maternal interactions: An evolutionary value? Obs. Gynecol. 2012, 120 Pt 1, 370–374. [Google Scholar] [CrossRef]
- Ching, T.; Ha, J.; Song, M.-A.; Tiirikainen, M.; Molnár, J.; Berry, M.J.; Towner, D.; Garmire, L.X. Genome-scale hypomethylation in the cord blood DNAs associated with early onset preeclampsia. Clin. Epigenet. 2015, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenrostro, J.D.; Wu, B.; Chang, H.Y.; Greenleaf, W.J. ATAC-seq: A Method for Assaying Chromatin Accessibility Genome-Wide. Curr. Protoc. Mol. Biol. 2015, 109, 21.29.1–21.29.9. [Google Scholar] [CrossRef] [PubMed]
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin accessibility and the regulatory epigenome. Nat. Rev. Genet. 2019, 20, 207–220. [Google Scholar] [CrossRef]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research in-volving human subjects. JAMA 2013, 310, 2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Turner, D.; Mesirov, J.P. igv.js: An embeddable JavaScript implementation of the Integrative Genomics Viewer (IGV). bioRxiv 2020. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Romano, O.; Miccio, A. GATA factor transcriptional activity: Insights from genome-wide binding profiles. IUBMB Life 2020, 72, 10–26. [Google Scholar] [CrossRef]
- Tijssen, M.R.; Cvejic, A.; Joshi, A.; Hannah, R.L.; Ferreira, R.; Forrai, A.; Bellissimo, D.C.; Oram, S.H.; Smethurst, P.A.; Wilson, N.K.; et al. Genome-wide Analysis of Simultaneous GATA1/2, RUNX1, FLI1, and SCL Binding in Megakaryocytes Identifies Hematopoietic Regulators. Dev. Cell 2011, 20, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Beck, D.; Thoms, J.A.I.; Perera, D.; Schütte, J.; Unnikrishnan, A.; Knezevic, K.; Kinston, S.J.; Wilson, N.K.; O’Brien, T.A.; Göttgens, B.; et al. Genome-wide analysis of transcriptional regulators in human HSPCs reveals a densely interconnected network of coding and noncoding genes. Blood 2013, 122, e12–e22. [Google Scholar] [CrossRef] [PubMed]
- Kanhere, A.; Hertweck, A.; Bhatia, U.; Gökmen, M.R.; Perucha, E.; Jackson, I.; Lord, G.M.; Jenner, R.G. T-bet and GATA3 orchestrate Th1 and Th2 differentiation through lineage-specific targeting of distal regulatory elements. Nat. Commun. 2012, 3, 1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, A.; Kong, S.W.; Ma, Q.; Pu, W.T. Co-occupancy by multiple cardiac transcription factors identifies transcriptional enhancers active in heart. Proc. Natl. Acad. Sci. USA 2011, 108, 5632–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulahian, R.; Casey, F.; Shen, J.; Qian, Z.R.; Shin, H.; Ogino, S.; A Weir, B.; Vazquez, F.; Liu, X.S.; Hahn, W.C.; et al. An integrative analysis reveals functional targets of GATA6 transcriptional regulation in gastric cancer. Oncogene 2013, 33, 5637–5648. [Google Scholar] [CrossRef] [Green Version]
- Tal, R. The Role of Hypoxia and Hypoxia-Inducible Factor-1Alpha in Preeclampsia Pathogenesis. Biol. Reprod. 2012, 87, 134. [Google Scholar] [CrossRef]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular evidence of placental hypoxia in preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 4299–4308. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.-Q.; Zhang, L. Hypoxia and the integrated stress response promote pulmonary hypertension and preeclampsia: Implications in drug development. Drug Discov. Today 2021, 26, 2754–2773. [Google Scholar] [CrossRef]
- Backes, C.H.; Markham, K.; Moorehead, P.; Cordero, L.; Nankervis, C.A.; Giannone, P.J. Maternal Preeclampsia and Neonatal Outcomes. J. Pregnancy 2011, 2011, 214365. [Google Scholar] [CrossRef]
- Lee, A.; Chow, B.L.; Skeith, L.; A Nicholas, B.J.; Poon, M.-C.; Poole, M.A.W.; O Agbani, P.E. Platelet Membrane Procoagulation in Preeclampsia. Blood 2020, 136 (Suppl. S1), 7. [Google Scholar] [CrossRef]
- Macey, M.; Bevan, S.; Alam, S.; Verghese, L.; Agrawal, S.; Beski, S.; Thuraisingham, R.; MacCallum, P. Platelet activation and endogenous thrombin potential in pre-eclampsia. Thromb. Res. 2010, 125, e76–e81. [Google Scholar] [CrossRef]
- Rambaldi, M.P.; Weiner, E.; Mecacci, F.; Bar, J.; Petraglia, F. Immunomodulation and preeclampsia. Best Pract. Research. Clin. Obstet. Gynaecol. 2019, 60, 87–96. [Google Scholar] [CrossRef]
- Kanellopoulos-Langevin, C.; Caucheteux, S.M.; Verbeke, P.; Ojcius, D.M. Tolerance of the fetus by the maternal immune system: Role of inflammatory mediators at the feto-maternal interface. Reprod. Biol. Endocrinol. 2003, 1, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nieuwenhoven, A.V.; Heineman, M.; Faas, M. The immunology of successful pregnancy. Hum. Reprod. Updat. 2003, 9, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Sakai, M.; Sasaki, Y.; Tanebe, K.; Tsuda, H.; Michimata, T. Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1:Th2 cell ratio during normal human pregnancy and preeclampsia. Clin. Exp. Immunol. 1999, 117, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Tindemans, I.; Serafini, N.; Di Santo, J.; Hendriks, R.W. GATA-3 Function in Innate and Adaptive Immunity. Immunity 2014, 41, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranganath, S.; Murphy, K.M. Structure and Specificity of GATA Proteins in Th2 Development. Mol. Cell. Biol. 2001, 21, 2716–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romao-Veiga, M.; Ribeiro, V.R.; Matias, M.L.; Nunes, P.R.; Romagnoli, G.G.; Peracoli, J.C.; Peracoli, M.T.S. DAMPs are able to skew CD4+ T cell subsets and increase the inflammatory profile in pregnant women with preeclampsia. J. Reprod. Immunol. 2022, 149, 103470. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-C.; Lin, J.-J.; Hsu, C.-L.; Juan, H.-F.; Lou, P.-J.; Huang, M.-C. GATA3 interacts with and stabilizes HIF-1α to enhance cancer cell invasiveness. Oncogene 2017, 36, 4243–4252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Before Labor | After Labor | Reference Range | |
|---|---|---|---|
| Urine | |||
| Protein | + + | − | − |
| 24 h urine protein (mg/24 h) | 876.58 ↑↑ | − | <150 |
| Blood * | |||
| BNP (pg/mL) | 909 ↑↑ | NA | <100 |
| PT (s) | 9.3 ↓ | 10.3 | 10~15 |
| FIB (g/L) | 3.88 ↑ | 5.75 ↑ | 1.8~3.5 |
| PT-R (%) | 121.10 ↑ | 111.70 | 75~120 |
| D-D (mg/L) | 0.79 ↑ | 1.12 ↑ | <0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Zhan, Z.; Li, N.; Huang, Y.; Li, Y.; Liu, L.; Shen, Q.; Chu, Q.; Wang, X.; Wu, B.; et al. Preliminary Interpretations of Epigenetic Profiling of Cord Blood in Preeclampsia. Genes 2022, 13, 888. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050888
Ma J, Zhan Z, Li N, Huang Y, Li Y, Liu L, Shen Q, Chu Q, Wang X, Wu B, et al. Preliminary Interpretations of Epigenetic Profiling of Cord Blood in Preeclampsia. Genes. 2022; 13(5):888. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050888
Chicago/Turabian StyleMa, Junrui, Zhongqun Zhan, Ning Li, Yanli Huang, Yan Li, Lu Liu, Qi Shen, Qiao Chu, Xiaonan Wang, Benqing Wu, and et al. 2022. "Preliminary Interpretations of Epigenetic Profiling of Cord Blood in Preeclampsia" Genes 13, no. 5: 888. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050888