
Expression Pattern of nos1 in the Developing Nervous System of Ray-Finned Fish

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Embryo Collections
2.2. Cloning and Probe Preparation
2.3. In Situ Hybridization
3. Results
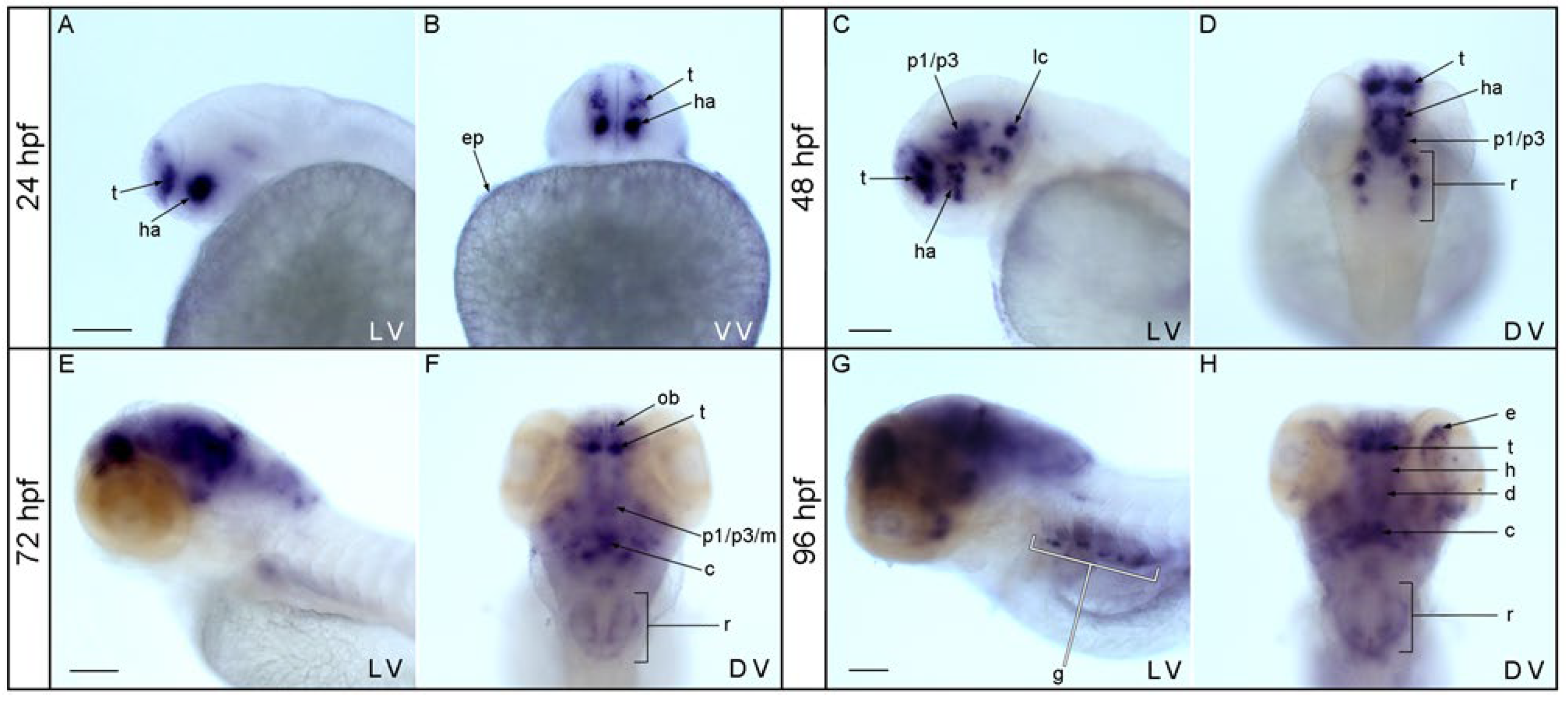
3.1. Expression Pattern of nos1 in the Brain of Zebrafish Embryos and Larvae
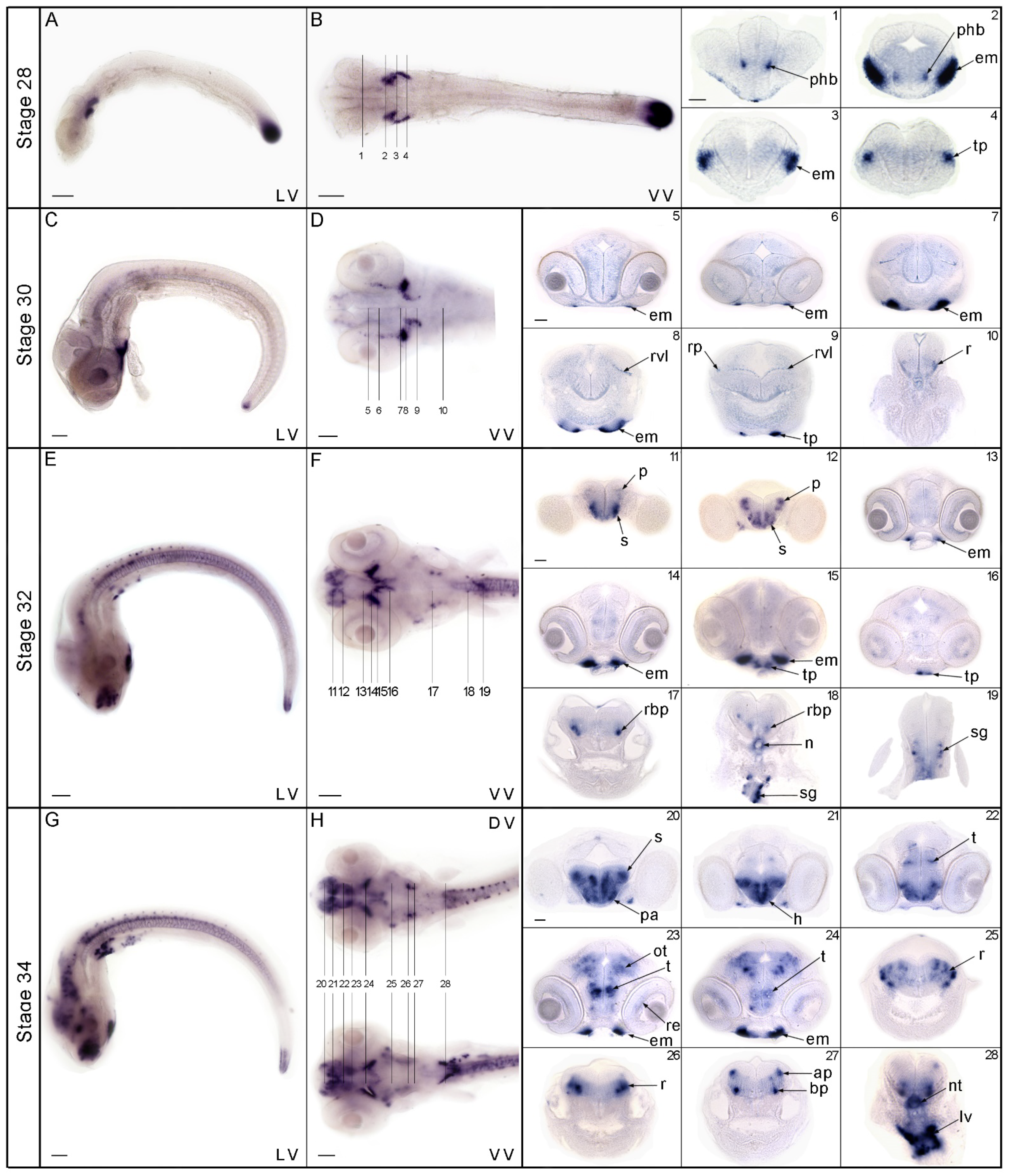
3.2. Expression Pattern of nos1 in the Brain of Medaka Embryos and Larvae
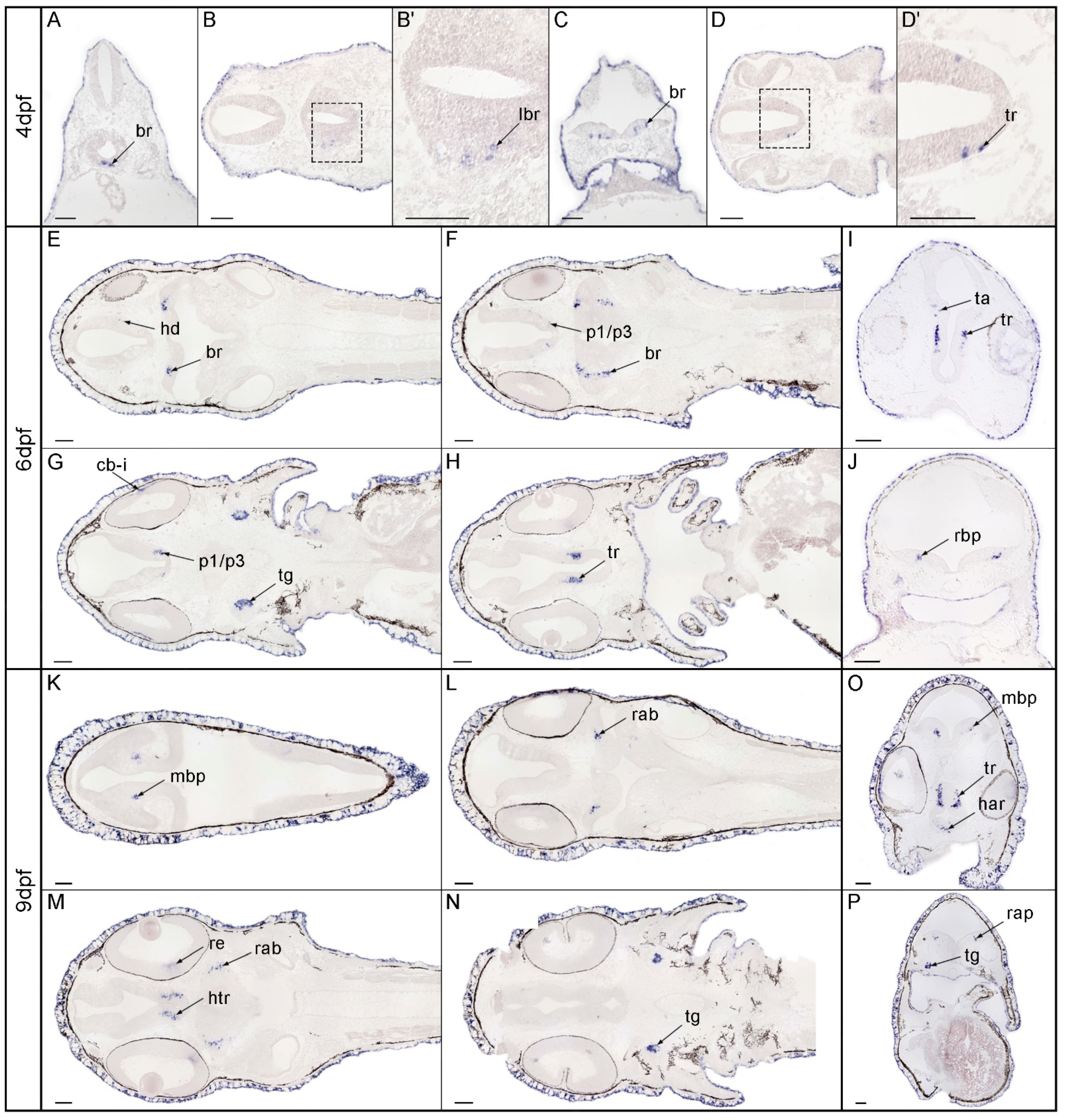
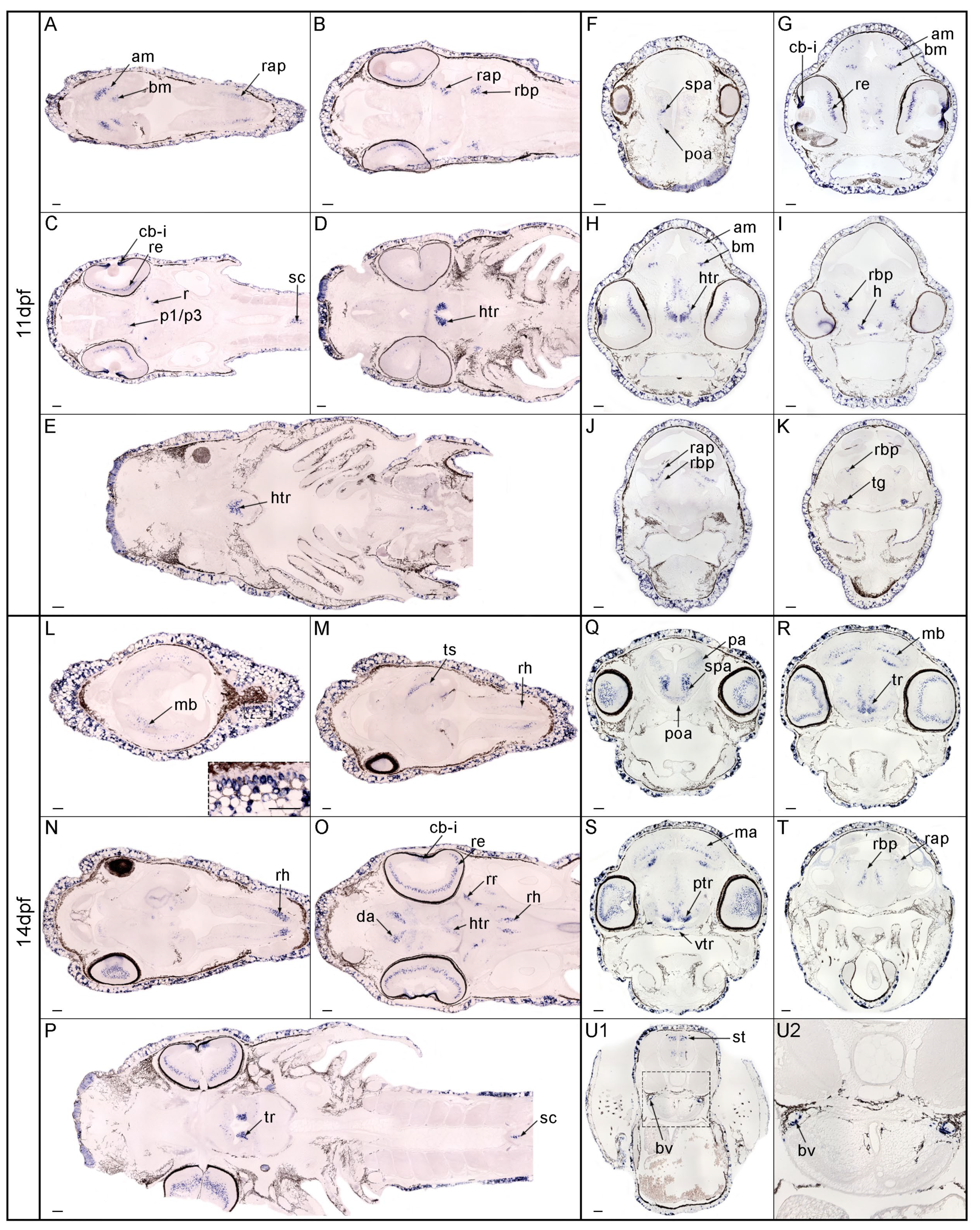
3.3. Expression Pattern of nos1 in the Brain of Gar Embryos and Larvae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Betancur-R, R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Ortí, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 162. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.C.; Ortí, G.; Huang, Y.; Sun, Y.; Baldwin, C.C.; Thompson, A.W.; Arcila, D.; Betancur-R., R.; Li, C.; Becker, L.; et al. Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proc. Natl. Acad. Sci. USA 2018, 115, 6249–6254. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.J. Origin and diversification of teleostean fishes. Ann. N. Y. Acad. Sci. 1969, 167, 18–30. [Google Scholar] [CrossRef]
- Moroz, L.L. Parallel evolution of Nitric Oxide signaling: Diversity of synthesis and memory pathways. Front. Biosci. 2011, 16, 2008. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Shi, Q.; Kusakabe, T.; Ohya, T.; Suzuki, N.; Urano, A. Localization of mRNAs encoding α and β subunits of soluble guanylyl cyclase in the brain of rainbow trout: Comparison with the distribution of neuronal nitric oxide synthase. Brain Res. 2004, 1013, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.; Ekambaram, P. Involvement of nitric oxide in learning & memory processes. Indian J. Med. Res. 2011, 133, 471–478. [Google Scholar] [PubMed]
- Penglee, R.; Gao, L.; Huang, Y.; Liu, L.; Nimitkul, S.; Bao, B. The role of nitric oxide and neuronal nitric oxide synthase in zebrafish (Danio rerio) shoaling. Aquac. Fish. 2021, 6, 565–573. [Google Scholar] [CrossRef]
- Andreakis, N.; D’Aniello, S.; Albalat, R.; Patti, F.P.; Garcia-Fernandez, J.; Procaccini, G.; Sordino, P.; Palumbo, A. Evolution of the Nitric Oxide Synthase Family in Metazoans. Mol. Biol. Evol. 2011, 28, 163–179. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Annona, G.; Sato, I.; Pascual-Anaya, J.; Braasch, I.; Voss, R.; Stundl, J.; Soukup, V.; Kuratani, S.; Postlethwait, J.H.; D’Aniello, S. Evolution of the nitric oxide synthase family in vertebrates and novel insights in gill development. bioRxiv 2021, 2021.06.14.448362. [Google Scholar] [CrossRef]
- Dreyer, J. Nitric Oxide Synthase (NOS)-Interacting Protein Interacts with Neuronal NOS and Regulates Its Distribution and Activity. J. Neurosci. 2004, 24, 10454–10465. [Google Scholar] [CrossRef] [PubMed]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br. J. Pharmacol. 2002, 135, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, R.; Alonso, J.R.; García-Ojeda, E.; Briñón, J.G.; Crespo, C.; Aijón, J. NADPH-Diaphorase in the central nervous system of the tench (Tinca tinca L., 1758). J. Comp. Neurol. 1995, 352, 398–420. [Google Scholar] [CrossRef] [PubMed]
- Bordieri, L.; Bonaccorsi di Patti, M.C.; Miele, R.; Cioni, C. Partial cloning of neuronal nitric oxide synthase (nNOS) cDNA and regional distribution of nNOS mRNA in the central nervous system of the Nile tilapia Oreochromis niloticus. Mol. Brain Res. 2005, 142, 123–133. [Google Scholar] [CrossRef]
- Brüning, G.; Katzbach, R.; Mayer, B. Histochemical and immunocytochemical localization of nitric oxide synthase in the central nervous system of the goldfish, Carassius auratus. J. Comp. Neurol. 1995, 358, 353–382. [Google Scholar] [CrossRef]
- Gaikwad, A.; Biju, K.C.; Barsagade, V.; Bhute, Y.; Subhedar, N. Neuronal nitric oxide synthase in the olfactory system, forebrain, pituitary and retina of the adult teleost Clarias batrachus. J. Chem. Neuroanat. 2009, 37, 170–181. [Google Scholar] [CrossRef]
- Holmqvist, B.I.; Östholm, T.; Alm, P.; Ekström, P. Nitric oxide synthase in the brain of a teleost. Neurosci. Lett. 1994, 171, 205–208. [Google Scholar] [CrossRef]
- Holmqvist, B.; Ellingsen, B.; Alm, P.; Forsell, J.; Øyan, A.M.; Goksøyr, A.; Fjose, A.; Seo, H.C. Identification and distribution of nitric oxide synthase in the brain of adult zebrafish. Neurosci. Lett. 2000, 292, 119–122. [Google Scholar] [CrossRef]
- Øyan, A.M.; Nilsen, F.; Goksøyr, A.; Holmqvist, B. Partial cloning of constitutive and inducible nitric oxide synthases and detailed neuronal expression of NOS mRNA in the cerebellum and optic tectum of adult Atlantic salmon (Salmo salar). Mol. Brain Res. 2000, 78, 38–49. [Google Scholar] [CrossRef]
- Virgili, M.; Poli, A.; Beraudi, A.; Giuliani, A.; Villani, L. Regional distribution of nitric oxide synthase and NADPH-diaphorase activities in the central nervous system of teleosts. Brain Res. 2001, 901, 202–207. [Google Scholar] [CrossRef]
- López, J.M.; Lozano, D.; Morales, L.; González, A. Pattern of Nitrergic Neuronal System Organization in the Brain of Two Holostean Fishes (Actinopterygii: Ginglymodi). Brain. Behav. Evol. 2017, 89, 117–152. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yao, Y.; Harumi, T.; Suzuki, N. Localization of the nitric oxide/cGMP signaling pathway-related genes and influences of morpholino knock-down of soluble guanylyl cyclase on medaka fish embryogenesis. Zoolog. Sci. 2003, 20, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.P.; Jadhao, A.G.; Bhoyar, R.C.; Palande, N.V.; Sinh, D.P. Neuroanatomical localization of nitric oxide synthase (nNOS) in the central nervous system of carp, Labeo rohita during post-embryonic development. Int. J. Dev. Neurosci. 2015, 46, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Poon, K.L.; Richardson, M.; Lam, C.S.; Khoo, H.E.; Korzh, V. Expression pattern of neuronal nitric acid oxide synthase in embryonic zebrafish. Gene Expr. Patterns 2003, 3, 463–466. [Google Scholar] [CrossRef]
- Holmqvist, B.; Ellingsen, B.; Forsell, J.; Zhdanova, I.; Alm, P. The early ontogeny of neuronal nitric oxide synthase systems in the zebrafish. J. Exp. Biol. 2004, 207, 923–935. [Google Scholar] [CrossRef]
- Braasch, I.; Gehrke, A.R.; Smith, J.J.; Kawasaki, K.; Manousaki, T.; Pasquier, J.; Amores, A.; Desvignes, T.; Batzel, P.; Catchen, J.; et al. The spotted gar genome illuminates vertebrate evolution and facilitates human-teleost comparisons. Nat. Genet. 2016, 48, 427–437. [Google Scholar] [CrossRef]
- Symonová, R.; Majtánová, Z.; Arias-Rodriguez, L.; Mořkovský, L.; Kořínková, T.; Cavin, L.; Pokorná, M.J.; Doležálková, M.; Flajšhans, M.; Normandeau, E.; et al. Genome Compositional Organization in Gars Shows More Similarities to Mammals than to Other Ray-Finned Fish. J. Exp. Zool. Part B Mol. Dev. Evol. 2017, 328, 607–619. [Google Scholar] [CrossRef]
- Puelles, L.; Ferran, J.L. Concept of neural genoarchitecture and its genomic fundament. Front. Neuroanat. 2012, 6, 47. [Google Scholar] [CrossRef]
- Braasch, I.; Guiguen, Y.; Loker, R.; Letaw, J.H.; Ferrara, A.; Bobe, J.; Postlethwait, J.H. Connectivity of vertebrate genomes: Paired-related homeobox (Prrx) genes in spotted gar, basal teleosts, and tetrapods. Comp. Biochem. Physiol. Part-C: Toxicol. Pharmacol. 2014, 163, 24–36. [Google Scholar] [CrossRef]
- Long, W.L.; Ballard, W.W. Normal embryonic stages of the longnose gar, Lepisosteus osseus. BMC Dev. Biol. 2001, 1, 6. [Google Scholar] [CrossRef]
- Nüsslein-Volhard, C.; Dham, R. Zebrafish: A Practical Approach; Oxford University Press: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000; ISBN 9994860577. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Conte, I.; Carrella, S.; Avellino, R.; Karali, M.; Marco-Ferreres, R.; Bovolenta, P.; Banfi, S. miR-204 is required for lens and retinal development via Meis2 targeting. Proc. Natl. Acad. Sci. USA 2010, 107, 15491–15496. [Google Scholar] [CrossRef] [PubMed]
- Strähle, U.; Blader, P.; Adam, J.; Ingham, P.W. A simple and efficient procedure for non-isotopic in situ hybridization to sectioned material. Trends Genet. 1994, 10, 75–76. [Google Scholar] [CrossRef]
- Jowett, T.; Ingham, P.W.; Henrique, D.; Lettice, L.; Wilkinson, D.; Yan, Y.L. An EMBO Practical Course; University of Newcastle: Newcastle, UK, 1995; p. 50. [Google Scholar]
- Schredelseker, T.; Veit, F.; Dorsky, R.I.; Driever, W. Bsx Is Essential for Differentiation of Multiple Neuromodulatory Cell Populations in the Secondary Prosencephalon. Front. Neurosci. 2020, 14, 525. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, D.R.; Saunders, L.M.; Miller, A.C. A single-cell transcriptome atlas for zebrafish development. Dev. Biol. 2020, 459, 100–108. [Google Scholar] [CrossRef]
- Richardson, M.K. Theories, laws, and models in evo-devo. J. Exp. Zool. Part B Mol. Dev. Evol. 2022, 338, 36–61. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L.; Rubenstein, J.L.R. A new scenario of hypothalamic organization: Rationale of new hypotheses introduced in the updated prosomeric model. Front. Neuroanat. 2015, 9, 27. [Google Scholar] [CrossRef]
- Brożko, N.; Baggio, S.; Lipiec, M.A.; Jankowska, M.; Szewczyk, Ł.M.; Gabriel, M.O.; Chakraborty, C.; Ferran, J.L.; Wiśniewska, M.B. Genoarchitecture of the Early Postmitotic Pretectum and the Role of Wnt Signaling in Shaping Pretectal Neurochemical Anatomy in Zebrafish. Front. Neuroanat. 2022, 16, 838567. [Google Scholar] [CrossRef]
- Bilbao, M.G.; Garrigós, D.; Martinez-Morga, M.P.; Toval, A.; Kutsenko, Y.; Bautista, R.; Barreda, A.; Ribeiro Do Couto, B.; Puelles, L.; Ferran, J.L. Prosomeric hypothalamic distribution of tyrosine hydroxylase positive cells in adolescent rats. Front. Neuroanat. 2022, 24. [Google Scholar] [CrossRef]
- Ferran, J.L. Architect genes of the brain: A look at brain evolution through genoarchitecture. Mètode Rev. difusió la Investig. 2016, 2017, 17–23. [Google Scholar] [CrossRef]
- Schredelseker, T.; Driever, W. Conserved Genoarchitecture of the Basal Hypothalamus in Zebrafish Embryos. Front. Neuroanat. 2020, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Villani, L. Development of NADPH-diaphorase activity in the central nervous system of the cichlid fish, Tilapia mariae. Brain. Behav. Evol. 1999, 54, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Harumi, T.; Watanabe, T.; Yamamoto, T.; Tanabe, Y.; Suzuki, N. Expression of membrane-bound and soluble guanylyl cyclase mRNAs in embryonic and adult retina of the medaka fish Oryzias latipes. Zoolog. Sci. 2003, 20, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Peter, V.S. Nitric oxide rectifies acid-base disturbance and modifies thyroid hormone activity during net confinement of air-breathing fish (Anabas testudineus Bloch). Gen. Comp. Endocrinol. 2013, 181, 115–121. [Google Scholar] [CrossRef]
- Cioni, C.; Angiulli, E.; Toni, M. Nitric Oxide and the Neuroendocrine Control of the Osmotic Stress Response in Teleosts. Int. J. Mol. Sci. 2019, 20, 489. [Google Scholar] [CrossRef]
- Winter, M.J.; Ashworth, A.; Bond, H.; Brierley, M.J.; McCrohan, C.R.; Balment, R.J. The caudal neurosecretory system: Control and function of a novel neuroendocrine system in fish. Biochem. Cell Biol. 2000, 78, 193–203. [Google Scholar] [CrossRef]
- Cioni, C.; Bordieri, L.; De Vito, L. Nitric oxide and neuromodulation in the caudal neurosecretory system of teleosts. Biochem. Physiol.-B Biochem. Mol. Biol. 2002, 132, 57–68. [Google Scholar] [CrossRef]
- Cioni, C.; Francia, N.; Greco, A.; De Vito, L.; Bordieri, L.; Crosetti, D. Development of the caudal neurosecretory system of the Nile tilapia Oreochromis niloticus: An immunohistochemical and electron microscopic study. J. Morphol. 2000, 243, 209–218. [Google Scholar] [CrossRef]
- Oka, S.; Chiba, A.; Honma, Y.; Iwanaga, T.; Fujita, T. Development of the caudal neurosecretory system of the chum salmon, Oncorhynchus keta, as revealed by immunohistochemistry for urotensins I and II. Cell Tissue Res. 1993, 272, 221–226. [Google Scholar] [CrossRef]
- Annona, G.; Caccavale, F.; Pascual-Anaya, J.; Kuratani, S.; De Luca, P.; Palumbo, A.; D’Aniello, S. Nitric Oxide regulates mouth development in amphioxus. Sci. Rep. 2017, 7, 8432. [Google Scholar] [CrossRef]
- Dong, X.; Fu, Q.; Liu, S.; Gao, C.; Su, B.; Tan, F.; Li, C. The expression signatures of neuronal nitric oxide synthase (NOS1) in turbot (Scophthalmus maximus L.) mucosal surfaces against bacterial challenge. Fish Shellfish Immunol. 2016, 59, 406–413. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Stage) | Head Region | Trunk Region | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tel | Di | Mes | Rho | Eye | Cn | Liv | Th | PNS | Nt | Sk | Tb | ||
| Early Embryo | Zebrafish (24 hpf) | + | ++ | ++ | |||||||||
| Medaka (st 28) | + | +++ | ++ | +++ | |||||||||
| Gar (4 dpf) | + | + | +++ | ||||||||||
| Late Embryo | Zebrafish (48 hpf) | ++ | ++ | ++ | |||||||||
| Medaka (st 30) | ++ | ++ | ++ | ++ | |||||||||
| Gar (6 dpf) | ++ | ++ | + | ++ | +++ | ||||||||
| Early Larva | Zebrafish (72 hpf) | ++ | ++ | ++ | |||||||||
| Medaka (st 32) | +++ | + | ++ | + | ++ | + | ++ | ||||||
| Gar (9 dpf) | + | + | + | + | + | +++ | |||||||
| Late Larva | Zebrafish (96 hpf) | ++ | ++ | ++ | ++ | ++ | |||||||
| Medaka (st 34) | ++ | ++ | ++ | +++ | ++ | ++ | +++ | ++ | ++ | ||||
| Gar (11–14 dpf) | +++ | +++ | ++ | +++ | +++ | ++ | ++ | +++ | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annona, G.; Ferran, J.L.; De Luca, P.; Conte, I.; Postlethwait, J.H.; D’Aniello, S. Expression Pattern of nos1 in the Developing Nervous System of Ray-Finned Fish. Genes 2022, 13, 918. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050918
Annona G, Ferran JL, De Luca P, Conte I, Postlethwait JH, D’Aniello S. Expression Pattern of nos1 in the Developing Nervous System of Ray-Finned Fish. Genes. 2022; 13(5):918. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050918
Chicago/Turabian StyleAnnona, Giovanni, José Luis Ferran, Pasquale De Luca, Ivan Conte, John H. Postlethwait, and Salvatore D’Aniello. 2022. "Expression Pattern of nos1 in the Developing Nervous System of Ray-Finned Fish" Genes 13, no. 5: 918. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050918