
Complete Chloroplast Genome Sequence of Triosteum sinuatum, Insights into Comparative Chloroplast Genomics, Divergence Time Estimation and Phylogenetic Relationships among Dipsacales

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, DNA Isolation, Sequencing and Assembly
2.2. Genome Annotation
2.3. Genome Structure and Comparative Analysis
2.4. Distribution of Simple Sequence Repeats (SSRs)
2.5. Codon Usage Pattern
2.6. Phylogenetic Analysis
2.7. Divergence Time Estimation
3. Results and Discussion
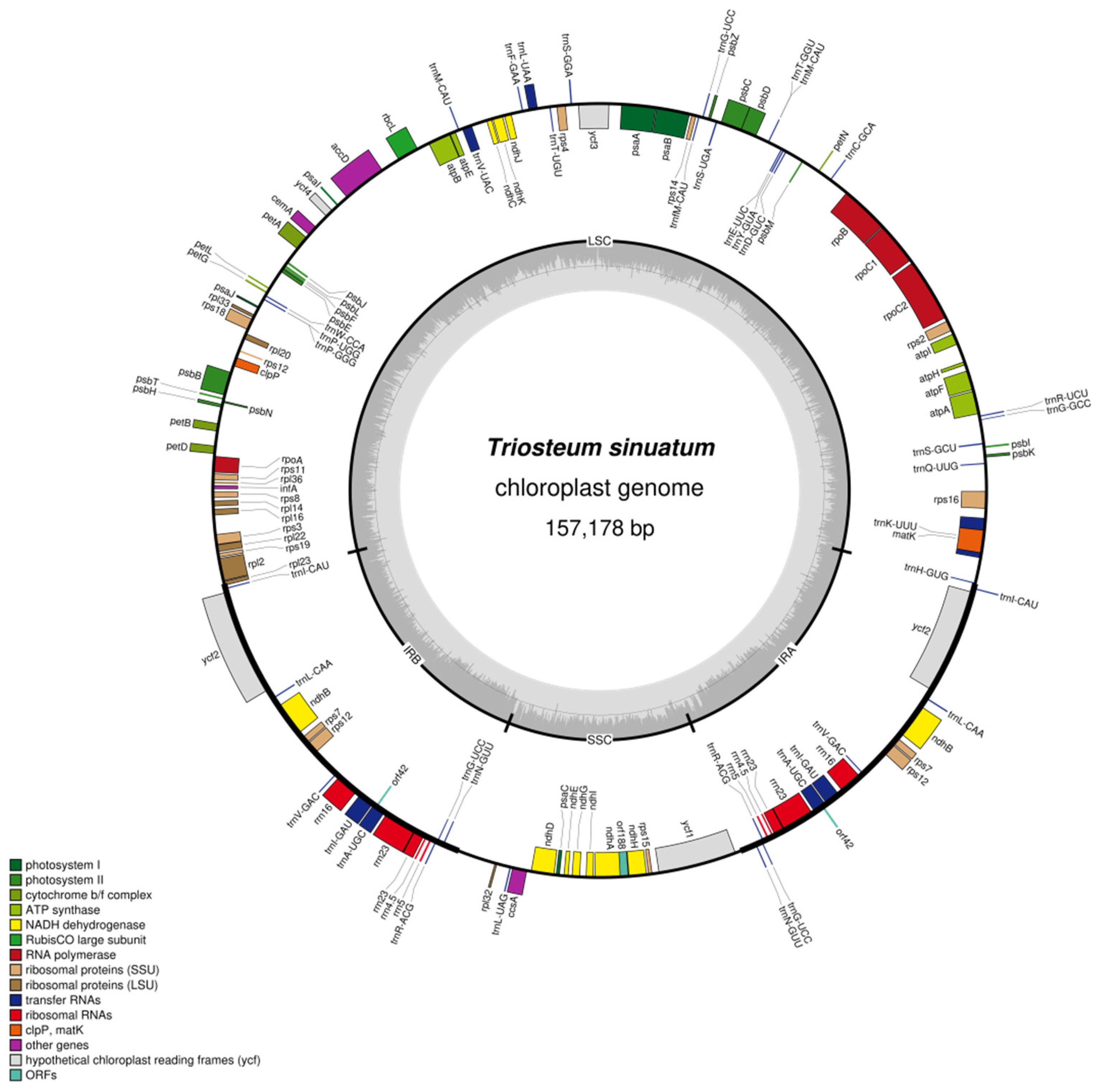
3.1. Plastome Features and Gene Content
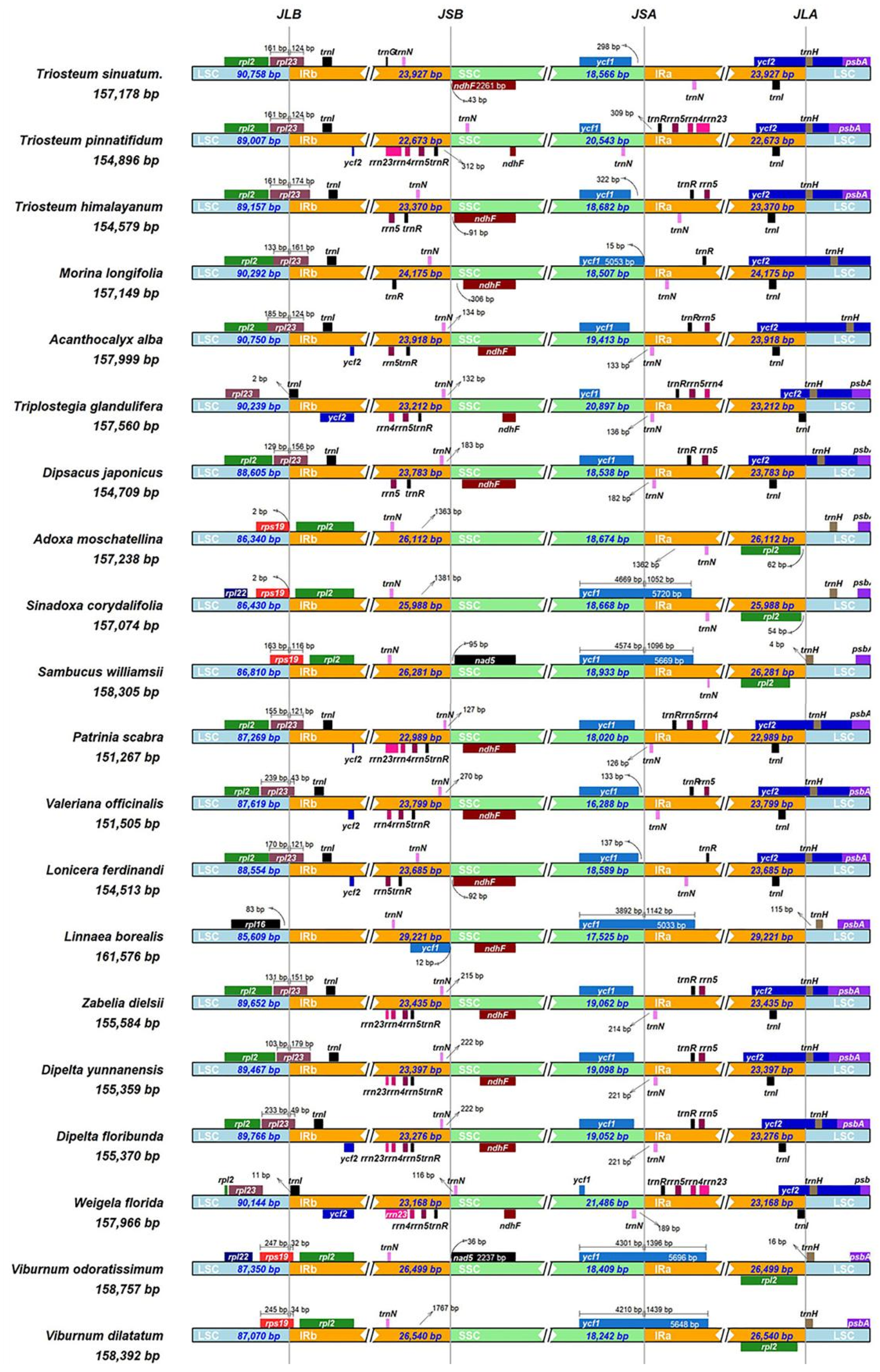
3.2. Comparative Analyses
3.3. SSR Analysis of Triosteum sinuatum Maxim. Chloroplast Genomes
3.4. Codon Usage Pattern
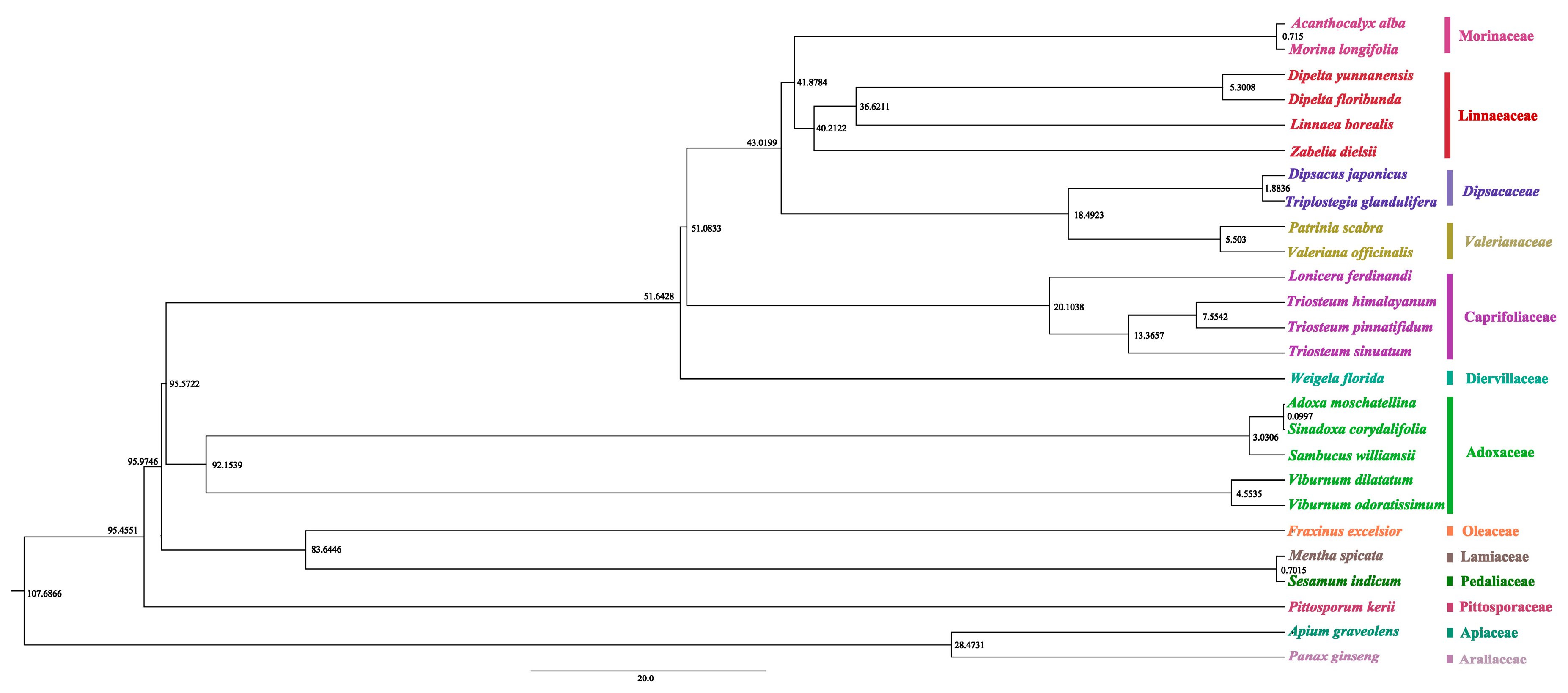
3.5. Phylogenetic Relationships
3.6. Divergence Time Estimation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dong, F.; Lin, Z.; Lin, J.; Ming, R.; Zhang, W. Chloroplast Genome of Rambutan and Comparative Analyses in Sapindaceae. Plants 2021, 10, 283. [Google Scholar] [CrossRef]
- Zha, X.; Wang, X.; Li, J.; Gao, F.; Zhou, Y. Complete chloroplast genome of Sophora alopecuroides (Papilionoideae): Molecular structures, comparative genome analysis and phylogenetic analysis. J. Genet. 2020, 99, 13. [Google Scholar] [CrossRef]
- Bai, G.Q.; Zhou, T.; Zhao, J.X.; Li, W.M.; Han, G.J.; Li, S.F. The complete chloroplast genome of Kolkwitzia amabilis (Caprifoliaceae), an endangered horticultural plant in China. Mitochondrial DNA 2017, 28, 296–297. [Google Scholar] [CrossRef]
- Wang, H.X.; Liu, H.; Moore, M.J.; Landrein, S.; Liu, B.; Zhu, Z.X.; Wang, H.F. Plastid phylogenomic insights into the evolution of the Caprifoliaceae s.l. (Dipsacales). Mol. Phylogenet. Evol. 2019, 142, 106641. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhang, C.; Guo, X.; Liu, Q.; Wang, K. Complete chloroplast genome of Camellia japonica genome structures, comparative and phylogenetic analysis. PLoS ONE 2019, 14, e0216645. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Zhang, K. Comparative analysis and phylogenetic evolution of the complete chloroplast genome of Ammopiptanthus. Acta Bot. Boreal.-Occident. Sin. 2020, 40, 1323–1332. [Google Scholar] [CrossRef]
- Fan, W.B.; Wu, Y.; Yang, J.; Shahzad, K.; Li, Z.H. Comparative Chloroplast Genomics of Dipsacales Species: Insights into Sequence Variation, Adaptive Evolution, and Phylogenetic Relationships. Front. Plant Sci. 2018, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gong, X.; Hu, C.; Hao, G. Phylogeography of an alpine species Primula secundiflora inferred from the chloroplast DNA sequence variation. J. Syst. Evol. 2008, 46, 13–22. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, Y.; Jiang, Z.; Liu, L.; Cai, J. Structure and Phylogeny of Chloroplast Genomes and Spermatophyte Flora in Chinese Theaceae. J. Northwest For. Univ. 2020, 35, 47–53. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Z.; Zhao, Y.; Li, X.; Zhao, Z.; Zhao, G. Chloroplast genome structural characteristics and phylogenetic relationships of Oleaceae. Chin. Bull. Bot. 2019, 54, 441–454. [Google Scholar]
- Donoghue, M.J.; Bell, C.D.; Winkworth, R.C. The Evolution of Reproductive Characters in Dipsacales. Int. J. Plant Sci. 2003, 164, S453–S464. [Google Scholar] [CrossRef]
- Xiang, C.L.; Dong, H.J.; Landrein, S.; Zhao, F.; Yu, W.B.; Soltis, D.E.; Soltis, P.S.; Backlund, A.; Wang, H.F.; Li, D.Z.; et al. Revisiting the phylogeny of Dipsacales: New insights from phylogenomic analyses of complete plastomic sequences. J. Syst. Evol. 2019, 58, 103–117. [Google Scholar] [CrossRef]
- Zhang, W.H.; Chen, Z.D.; Li, J.H.; Chen, H.B.; Tang, Y.C. Phylogeny of the Dipsacales, s.l. based on chloroplast trnL-F and ndhF sequences. Mol. Phylogenet. Evol. 2003, 26, 176–189. [Google Scholar] [CrossRef]
- Landrein, S.; Prenner, G.; Chase, M.W.; Clarkson, J.J. Abelia and relatives: Phylogenetics of Linnaeeae (Dipsacales–Caprifoliaceae s.l.) and a new interpretation of their inflorescence morphology. Bot. J. Linn. Soc. 2012, 169, 692–713. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.R.; Gao, Q.B.; Zhang, F.Q.; Khan, G.; Chen, S.L. Westwards and northwards dispersal of Triosteum himalayanum (Caprifoliaceae) from the Hengduan Mountains region based on chloroplast DNA phylogeography. PeerJ 2018, 6, e4748. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. Caprifoliaceae . In Flora of China; Yang, Q., Landrein, S., Osborne, J., Borosova, R., Eds.; Science Press: Beijing, China, 2011; Volume 19, pp. 616–617. [Google Scholar]
- Chai, X.; Su, Y.; Yan, S.; Huang, X. Chemical Constituents of the Roots of Triosteum pinnatifidum. Chem. Nat. Compd. 2014, 50, 1149–1152. [Google Scholar] [CrossRef]
- Chai, X.; Su, Y.-F.; Zheng, Y.-H.; Yan, S.-L.; Zhang, X.; Gao, X.-M. Iridoids from the roots of Triosteum pinnatifidum. Biochem. Syst. Ecol. 2010, 38, 210–212. [Google Scholar] [CrossRef]
- Ding, H.; Chen, Y.; Zhao, J.; Song, Q.; Gao, K. Chemical constituents from the aerial parts of Triosteum pinnatifidum. Chem. Nat. Compd. 2013, 49, 95–96. [Google Scholar] [CrossRef]
- Li, Z. Studies on the chemical constituents of Triosteum himalayanum. Master Thesis, Lanzhou University, Lanzhou, China, 2009. [Google Scholar]
- Liu, H.R.; Khan, G.; Gao, Q.; Zhang, F.; Liu, W.; Wang, Y.; Fang, J.; Chen, S.; Afridi, S.G. Dispersal into the Qinghai-Tibet plateau: Evidence from the genetic structure and demography of the alpine plant Triosteum pinnatifidum. PeerJ 2022, 10, e1275. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.-J.; Park, H.-S.; Koo, H.J.; Park, J.Y.; Lee, D.Y.; Kang, K.B.; Han, S.I.; Sung, S.H.; Yang, T.J. The complete chloroplast genome sequence of Korean Lonicera japonica and intra-species diversity. Mitochondrial DNA Part B 2018, 3, 941–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Liu, J.; An, J.; Wang, M.; Wang, Q. Characterization of the complete chloroplast genome of Lonicera macranthoides. Mitochondrial DNA Part B 2018, 3, 1002–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, J.; Li, P.; Wang, H. The characteristics of the chloroplast genome of the Michelia martini. Mitochondrial DNA Part B 2020, 5, 2725–2726. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvonen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Yin, X.; Wen, Q.; Wang, J.; Li, T.; Ye, J.; Xu, L. Characterization of microsatellites in complete chloroplast genome of the genus Camellia and marker development. Mol. Plant Breed. 2018, 16, 6761–6769. [Google Scholar]
- Yang, Y.; Jiao, J.; Fan, X.; Zhang, Y.; Jiang, J.; Li, M.; Liu, C. Complete Chloroplast Genome Sequence and Characteristics Analysis of Vitis ficifolia. Acta Hortic. Sin. 2019, 46, 635–648. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Yang, Y.; Fu, X.; Liu, L.; Jiang, Z.; Cai, J. Plastid genomes of Elaeagnus mollis: Comparative and phylogenetic analyses. J. Genet. 2020, 99, 85. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, C.; Zhao, X.; Fei, Z.; Wan, K.; Zhang, Z.; Pang, X.; Yin, X.; Bai, Y.; Sun, X.; et al. The jujube genome provides insights into genome evolution and the domestication of sweetness/acidity taste in fruit trees. PLoS Genet. 2016, 12, e1006433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.L.; Fan, W.B.; Wang, N.; Dong, P.B.; Zhang, T.T.; Yue, M.; Li, Z.H. Evolutionary Analysis of Plastid Genomes of Seven Lonicera, L. Species: Implications for Sequence Divergence and Phylogenetic Relationships. Int. J. Mol. Sci. 2018, 19, 4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzahrani, D.A. Complete Chloroplast Genome of Abutilon fruticosum: Genome Structure, Comparative and Phylogenetic Analysis. Plants 2021, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, S.; Liu, Y.; Yuan, Q.; Sun, J.; Guo, L. Chloroplast genome variation and phylogenetic relationships of Atractylodes species. BMC Genom. 2021, 22, 103. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, D.A.; Yaradua, S.S.; Albokhari, E.J.; Abba, A. Complete chloroplast genome sequence of Barleria prionitis, comparative chloroplast genomics and phylogenetic relationships among Acanthoideae. BMC Genom. 2020, 21, 393. [Google Scholar] [CrossRef]
- Li, Y.; Dong, Y.; Liu, Y.; Yu, X.; Yang, M.; Huang, Y. Comparative Analyses of Euonymus Chloroplast Genomes: Genetic Structure, Screening for Loci with Suitable Polymorphism, Positive Selection Genes, and Phylogenetic Relationships within Celastrineae. Front. Plant Sci. 2020, 11, 593984. [Google Scholar] [CrossRef]
- Raman, G.; Park, K.T.; Kim, J.H.; Park, S. Characteristics of the completed chloroplast genome sequence of Xanthium spinosum: Comparative analyses, identification of mutational hotspots and phylogenetic implications. BMC Genom. 2020, 21, 855. [Google Scholar] [CrossRef]
- Chincoya, D.A.; Sanchez-Flores, A.; Estrada, K.; Diaz-Velasquez, C.E.; Gonzalez-Rodriguez, A.; Vaca-Paniagua, F.; Davila, P.; Arias, S.; Solorzano, S. Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales). Genes 2020, 11, 830. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Yang, J.X.; Li, H.K.; Zhao, H.S. Chloroplast Genomes of Two Species of Cypripedium: Expanded Genome Size and Proliferation of AT-Biased Repeat Sequences. Front. Plant Sci. 2021, 12, 609729. [Google Scholar] [CrossRef] [PubMed]
- Ran, H.; Liu, Y.; Wu, C.; Cao, Y. Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae). Plants 2020, 9, 1143. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Wu, Q.; Yi, Y.; Yu, Z. The complete chloroplast genome of Lonicera hypoglauca Miq (Caprifoliaceae: Dipsacales) from Guangxi, China. Mitochondrial DNA Part B 2021, 6, 450–451. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, M.K.; Gangaraj, K.P.; Prabhudas, S.K.; Prasad, T.S.K. The complete chloroplast genome data of Areca catechu (Arecaceae). Data Brief 2020, 33, 106444. [Google Scholar] [CrossRef]
- Parvathy, S.T.; Udayasuriyan, V.; Bhadana, V. Codon usage bias. Mol. Biol. Rep. 2022, 49, 539–565. [Google Scholar] [CrossRef]
- Randall, T.A.; Dwyer, R.A.; Huitema, E.; Beyer, K.; Cvitanich, C.; Kelkar, H.; Fong, A.V.A.; Gates, K.; Roberts, S.; Yatzkan, E.; et al. Large-Scale gene discovery in the oomycete Phytophthora infestans reveals likely components of phytopathogenicity shared with true fungi. Mol. Plant-Microbe Interact. 2005, 18, 229. [Google Scholar] [CrossRef] [Green Version]
- Downie, S.R.; Jansen, R.K. A Comparative Analysis of Whole Plastid Genomes from the Apiales: Expansion and Contraction of the Inverted Repeat, Mitochondrial to Plastid Transfer of DNA, and Identification of Highly Divergent Noncoding Regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, L.; Wang, W.; Zhang, Z.; Du, H.; Qu, Z.; Li, X.Q.; Xiang, H. Differences in Codon Usage Bias between Photosynthesis-Related Genes and Genetic System-Related Genes of Chloroplast Genomes in Cultivated and Wild Solanum Species. Int. J. Mol. Sci. 2018, 19, 3142. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.; Pyck, N.; Smets, E. Phylogeny of the Linnaea clade: Are Abelia and Zabelia closely related? Mol. Phylogenet. Evol. 2010, 57, 741–752. [Google Scholar] [CrossRef]
- Winkworth, R.C.; Bell, C.D.; Donoghue, M.J. Mitochondrial sequence data and Dipsacales phylogeny: Mixed models, partitioned Bayesian analyses, and model selection. Mol. Phylogenet. Evol. 2008, 46, 830–843. [Google Scholar] [CrossRef]
- Bell, C.D.; Donoghue, M.J. Dating the Dipsacales: Comparing models, genes and evolutionary implications. Am. J. Bot. 2005, 92, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.R.; Fang, J.; Xia, M.; Xiao, Q.; Zhang, D.; Xie, J.; Chen, S. The complete chloroplast genome of Triosteum himalayanum (Caprifoliaceae), a perennial alpine herb. Mitochondrial DNA Part B 2019, 4, 4194–4195. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Genebank | Size (bp) | LSC (bp) | SSC (bp) | IR (bp) | Total GC (%) | Total Genes | Protein Genes | tRNA Genes | rRNA Genes |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Morinaceae | Morina longifolia | NC045046 | 157,149 | 90,292 | 18,679 | 24,089 | 38.60 | 128 | 83 | 37 | 8 |
| Acanthocalyx alba | NC045055 | 157,999 | 90,750 | 19,413 | 23,918 | 38.36 | 128 | 83 | 37 | 8 | |
| Dipsacaceae | Triplostegia glandulifera | NC045051 | 157,560 | 90,239 | 21,099 | 23,111 | 37.93 | 128 | 83 | 37 | 8 |
| Dipsacus japonicus | MN524645 | 154,709 | 88,605 | 18,538 | 23,783 | 38.99 | 128 | 83 | 37 | 8 | |
| Adoxaceae | Adoxa moschatellina | NC034792 | 157,238 | 86,340 | 18,674 | 26,112 | 37.79 | 128 | 81 | 39 | 8 |
| Sinadoxa corydalifolia | NC032040 | 157,074 | 86,430 | 18,668 | 25,988 | 37.76 | 131 | 83 | 40 | 8 | |
| Sambucus williamsii | NC033878 | 158,305 | 86,810 | 18,933 | 26,281 | 37.93 | 132 | 84 | 40 | 8 | |
| Viburnum odoratissimum | MN894600 | 158,757 | 87,350 | 18,409 | 26,499 | 38.10 | 132 | 87 | 37 | 8 | |
| Viburnum dilatatum | MW346666 | 158,392 | 87,070 | 18,242 | 26,540 | 38.10 | 130 | 83 | 39 | 8 | |
| Valerianaceae | Patrinia scabra | NC045048 | 151,267 | 87,269 | 18,020 | 22,989 | 38.58 | 128 | 83 | 37 | 8 |
| Valeriana officinalis | NC045052 | 151,505 | 87,619 | 16,288 | 23,799 | 38.45 | 128 | 83 | 37 | 8 | |
| Caprifoliaceae | Triosteum pinnatifidum | NC037952 | 154,896 | 89,007 | 20,543 | 22,673 | 38.54 | 128 | 83 | 37 | 8 |
| Triosteum himalayanum | NC045219 | 154,579 | 89,157 | 18,682 | 23,370 | 38.38 | 132 | 86 | 38 | 8 | |
| Triosteum sinuatum | MW526077 | 157,178 | 90,758 | 18,656 | 23,882 | 38.31 | 135 | 86 | 41 | 8 | |
| Lonicera ferdinandi | NC040963 | 154,513 | 88,554 | 18,589 | 23,685 | 38.44 | 128 | 83 | 37 | 8 | |
| Linnaeaceae | Linnaea borealis | MN548092 | 161,576 | 85,609 | 17,547 | 29,210 | 38.26 | 134 | 89 | 37 | 8 |
| Zabelia dielsii | NC046599 | 155,584 | 89,652 | 19,062 | 23,435 | 38.39 | 128 | 83 | 37 | 8 | |
| Dipelta yunnanensis | NC042201 | 155,359 | 89,467 | 19,098 | 23,397 | 38.45 | 128 | 83 | 37 | 8 | |
| Dipelta floribunda | NC037955 | 155,370 | 89,431 | 19,052 | 23,378 | 38.38 | 128 | 83 | 37 | 8 | |
| Diervillaceae | Weigela florida | MN524626 | 157,966 | 90,144 | 21,486 | 23,168 | 38.01 | 128 | 83 | 37 | 8 |
| Category | Group of Genes | Name of Genes |
|---|---|---|
| Transcription and Translation | Large subunit of ribosome | rpl33, rpl20, rpl36, rpl14, rpl16 a, rpl22, rpl2 a, rpl23, rpl32 |
| Small subunit of ribosome | rps16, rps2, rps14, rps4, rps18, rps12 b,*, rps11, rps8, rps3, rps19, rps7*, rps15 | |
| RNA polymerase | rpoC2, rpoC1 a, rpoB, rpoA | |
| Ribosomal RNAs | rrn16*, rrn23*, rrn4.5*, rrn5* | |
| Transfer RNAs | trnH-GUG, trnK-UUU a, trnQ-UUG, trnS-GCU, trnG-GCC a, trnR-UCU, trnC-GCA, trnD-GUC, trnY-GUA, trnE-UUC, trnT-GGU, trnS-UGA, trnG-UCC, trnfM-CAU, trnS-GGA, trnT-UGU, trnL-UAAa, trnF-GAA, trnV-UAC a,*, trnM-CAU, trnW-CCA, trnP-UGG, trnI-CAU *, trnL-CAA *, trnV-GAC *, trnI-GAU *, trnA-UGC *, trnR-ACG *, trnN-GUU *, trnL-UAG | |
| Photosynthesis | Photosystem I | psaB, psaA, psaI, psaJ, psaC |
| Photosystem II | psbA, psbK, psbI, psbM, psbD, psbC, psbZ, psbJ, psbL, psbF, psbE, psbB, psbT, psbN, psbH | |
| NADH dehydrogenase | ndhJ, ndhK, ndhC, ndhB a,*, ndhF, ndhD, ndhE, ndhG, ndhI, ndhA a, ndhH | |
| Cytochrome b6/f complex | petN, petA, petL, petG, petB a, petD a | |
| ATP synthase | atpA, atpF a, atpH, atpI, atpE, atpB | |
| Rubisco large subunit | rbcL | |
| ATP-dependent protease subunit p | clpP a | |
| Other genes | Chloroplast envelope membrane protein | cemA |
| Maturase | matK | |
| c-type | ccsA | |
| Subunit Acetyl- CoA-Carboxylate | accD | |
| Genes of unknown function | Conserved ORFs | ycf1, ycf2 *, ycf3, ycf4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Liu, W.; Ahmad, I.; Xiao, Q.; Li, X.; Zhang, D.; Fang, J.; Zhang, G.; Xu, B.; Gao, Q.; et al. Complete Chloroplast Genome Sequence of Triosteum sinuatum, Insights into Comparative Chloroplast Genomics, Divergence Time Estimation and Phylogenetic Relationships among Dipsacales. Genes 2022, 13, 933. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050933
Liu H, Liu W, Ahmad I, Xiao Q, Li X, Zhang D, Fang J, Zhang G, Xu B, Gao Q, et al. Complete Chloroplast Genome Sequence of Triosteum sinuatum, Insights into Comparative Chloroplast Genomics, Divergence Time Estimation and Phylogenetic Relationships among Dipsacales. Genes. 2022; 13(5):933. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050933
Chicago/Turabian StyleLiu, HaiRui, WenHui Liu, Israr Ahmad, QingMeng Xiao, XuMin Li, DeJun Zhang, Jie Fang, GuoFan Zhang, Bin Xu, QingBo Gao, and et al. 2022. "Complete Chloroplast Genome Sequence of Triosteum sinuatum, Insights into Comparative Chloroplast Genomics, Divergence Time Estimation and Phylogenetic Relationships among Dipsacales" Genes 13, no. 5: 933. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050933