
Haplotypes and Sequence Variation in the Ovine Adiponectin Gene (ADIPOQ)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sheep Investigated and DNA Collection
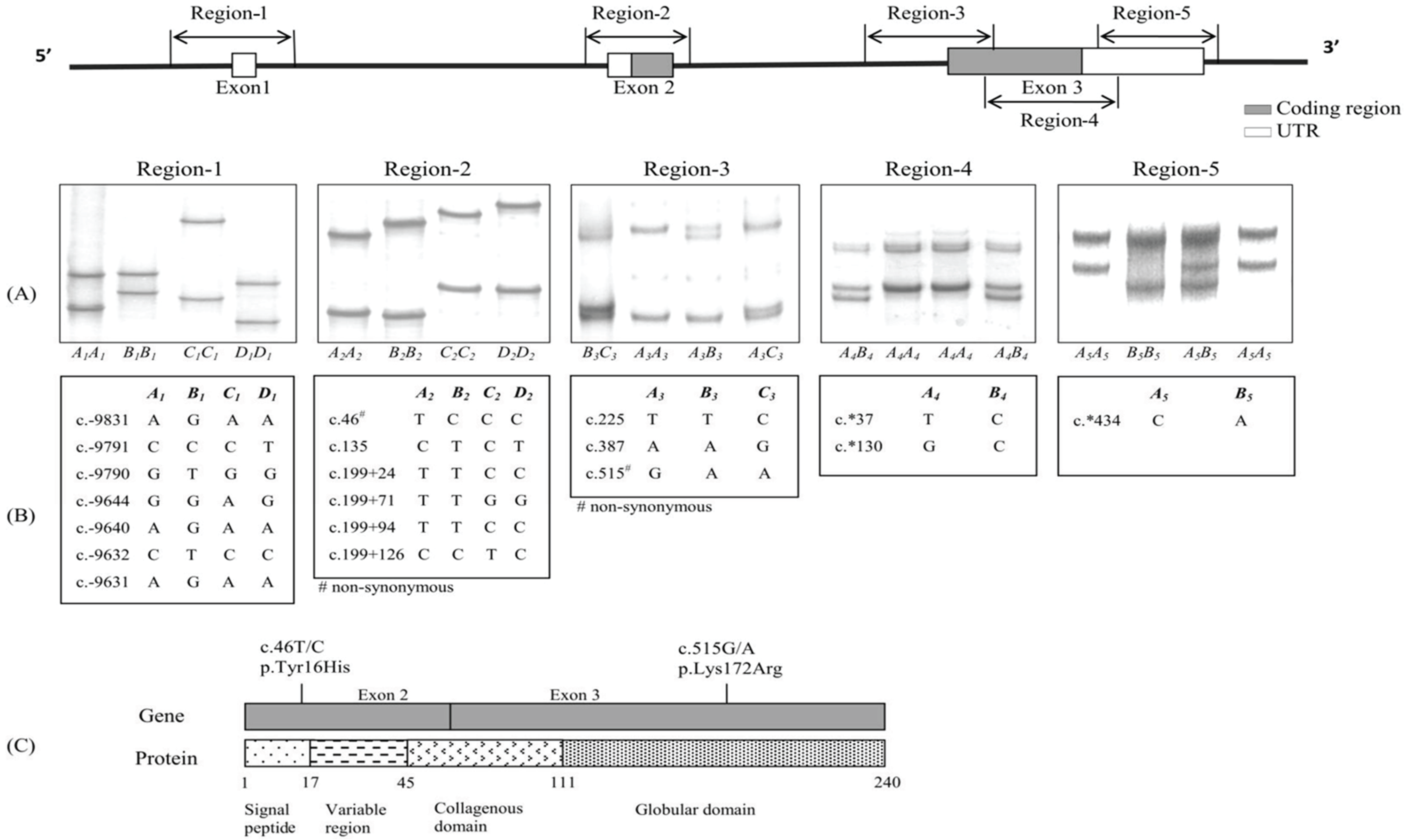
2.2. PCR Amplification and SSCP Analysis
2.3. Sequence Analyses
2.4. Haplotype Determination

{kind=link}
| Region Amplified | Primer Binding Region a | Primer Sequence (5'–3') | PCR Annealing Temperature | Amplified Size | SSCP Condition |
|---|---|---|---|---|---|
| Region-1 | 198617873_198617893 | F: TTCCTGCTTCTGATCTTGACC | 58 °C | 388 bp | 300 V, 14%, 17 °C |
| 198618239_198618260 | R: CAGCCTAGAAATTGAATCAGTC | ||||
| Region-2 | 198627771_198627789 | F: ACAGCGTGGATCTGGGTTC | 62 °C | 390 bp | 200 V, 14%, 32.5 °C |
| 198628140_198628159 | R: CACAATTCACTTTCGGCTGC | ||||
| Region-3 | 198628889_198628909 | F: GGTCTTCTTGTTCTCTAGGTC | 58 °C | 391 bp | 200 V, 12%, 20 °C |
| 198629260_198629279 | R: TGGTCCACGTTCTGGTTCTG | ||||
| Region-4 | 198629237_198629256 | F: TGCTCTTCACCCACGACCAG | 58 °C | 373 bp | 200 V, 14%, 26 °C |
| 198629583_198629605 | R: GTCCTGCGAACATAGTATATC | ||||
| Region-5 | 198629532_198629553 | F: GGATTCTGAACATCATTCATTC | 58 °C | 455 bp | 390 V, 14%, 5 °C |
| 198629967_198629986 | R: CAGATGAGTTGGTGGGAGAC |

3. Results
3.1. Identification of Sequence Variants of Ovine ADIPOQ
3.2. Initial Screen for Variation in ADIPOQ
3.3. Frequencies of the Sequence Variants in Different Breeds
| Breed | n | Region-1 | Region-2 | Region-3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | B1 | C1 | D1 | A2 | B2 | C2 | D2 | A3 | B3 | C3 | |||||
| Merino | 68 | 50.0 | 48.5 | 0.0 | 1.5 | 55.2 | 5.2 | 9.6 | 30.2 | 66.9 | 11.8 | 21.3 | |||
| Romney | 71 | 52.8 | 41.6 | 4.9 | 0.7 | 76.8 | 0.0 | 4.9 | 18.3 | 49.3 | 1.4 | 49.3 | |||
| Suffolk | 42 | 53.6 | 46.4 | 0.0 | 0.0 | 75.0 | 1.2 | 0.0 | 23.8 | 35.7 | 0.0 | 64.3 | |||
| Texel | 22 | 56.8 | 43.2 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 65.9 | 6.8 | 27.3 | |||
| Corriedale | 41 | 39.0 | 57.3 | 0.0 | 3.7 | 75.6 | 2.4 | 6.1 | 15.9 | 28.1 | 3.7 | 68.3 | |||
| Perendale | 14 | 57.1 | 39.3 | 0.0 | 3.6 | 89.3 | 0.0 | 0.0 | 10.7 | 42.9 | 0.0 | 57.1 | |||
| Dorper | 39 | 76.9 | 23.1 | 0.0 | 0.0 | 56.4 | 1.3 | 38.5 | 3.9 | 53.9 | 5.1 | 41.0 | |||
| Dorset Down | 19 | 81.6 | 18.4 | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 34.2 | 0.0 | 65.8 | |||
| Overall | 316 | 55.7 | 42.1 | 1.1 | 1.1 | 72.78 | 1.7 | 8.7 | 16.8 | 49.1 | 4.4 | 46.5 | |||
3.4. Haplotypes Spanning Region-1 to Region-3
| Haplotype | Frequency | Number (n) |
|---|---|---|
| A1-C3 | 30.1% | 139 |
| A1-A3 | 26.4% | 123 |
| B1-A3 | 23.2% | 108 |
| B1-C3 | 14.9% | 69 |
| B1-B3 | 3.3% | 15 |
| C1-A3 | 1.1% | 5 |
| A1-B3 | 0.7% | 3 |
| C1-B3 | 0.2% | 1 |
| C1-C3 | 0.2% | 1 |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yokota, T.; Meka, C.S.R.; Medina, K.L.; Igarashi, H.; Comp, P.C.; Takahashi, M.; Nishida, M.; Oritani, K.; Miyagawa, J.-I.; Funahashi, T.; et al. Paracrine regulation of fat cell formation in bone marrow cultures via adiponectin and prostaglandins. J. Clin. Invest. 2002, 109, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Morsci, N.S.; Schnabel, R.D.; Taylor, J.F. Association analysis of adiponectin and somatostatin polymorphisms on BTA1 with growth and carcass traits in Angus cattle. Anim. Genet. 2006, 37, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, Y.H.; Jiang, H.; Xiao, S.Q.; Wang, S.; Ma, Q.; Sun, G.J.; Li, F.J.; Deng, Q.; Dai, L.S.; et al. Detection of differentially expressed genes in the longissimus dorsi of Northeastern Indigenous and Large White pigs. Genet. Mol. Res. 2011, 10, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Arita, Y.; Yamagata, K.; Matsukawa, Y.; Okutomi, K.; Horie, M.; Shimomura, I.; Hotta, K.; Kuriyama, H.; Kihara, S.; et al. Genomic structure and mutations in adipose-specific gene, adiponectin. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, W.-C.; Jean, P.L., St.; Mitchell, B.D.; Pollin, T.I.; Knowler, W.C.; Ehm, M.G.; Bell, C.J.; Sakul, H.; Wagner, M.J.; Burns, D.K.; et al. Genome-wide and fine-mapping linkage studies of type 2 diabetes and glucose traits in the Old Order Amish: Evidence for a new diabetes locus on chromosome 14q11 and confirmation of a locus on chromosome 1q21-q24. Diabetes 2003, 52, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Wang, M.; Zhong, D.; Shi, D.; Ma, L.; Tong, N.; Zhang, Z. ADIPOQ polymorphisms are associated with type 2 diabetes mellitus: A meta‐analysis study. Diabetes Metab. Res. Rev. 2013, 29, 532–545. [Google Scholar] [PubMed]
- Yang, Y.; Zhang, F.; Ding, R.; Skrip, L.; Wang, Y.; Lei, H.; Hu, D. ADIPOQ gene polymorphisms and cancer risk: A meta-analysis. Cytokine 2013, 61, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Ramya, K.; Ayyappa, K.A.; Ghosh, S.; Mohan, V.; Radha, V. Genetic association of ADIPOQ gene variants with type 2 diabetes, obesity and serum adiponectin levels in south Indian population. Gene 2013, 532, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jiang, H.; Kuang, J.; Hou, X.; Feng, Y.; Su, Z. Genetic variations in ADIPOQ gene are associated with chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e50848. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Du, Y.; Zhang, C.; Shi, X.; Chen, D.; Sun, J.; Jin, Q.; Lan, X.; Chen, H. Polymorphism in a microsatellite of the acrp30 gene and its association with growth traits in goats. Biochem. Genet. 2011, 49, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.H.; Xiong, Y.Z.; Deng, C.Y.; Jiang, S.W.; Zuo, B.; Zheng, R.; Li, F.E.; Lei, M.G. Association of the AG polymorphism in porcine adiponectin gene with fat deposition and carcass traits. Asian-Australas. J. Anim. Sci. 2006, 19, 779–783. [Google Scholar] [CrossRef]
- Houde, A.A.; Murphy, B.D.; Mathieu, O.; Bordignon, V.; Palin, M.F. Characterization of swine adiponectin and adiponectin receptor polymorphisms and their association with reproductive traits. Anim. Genet. 2008, 39, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hickford, J.G.H.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G.H. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, S.; Liu, X.; Wang, J.; Luo, Y.; Hickford, J.G.H. Haplotyping using a combination of polymerase chain reaction-single-strand conformational polymorphism analysis and haplotype-specific PCR amplification. Anal. Biochem. 2014, 466, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ye, E.; Si, G.; Chen, L.; Cai, L.; Ye, C.; Zhang, C.; Lu, X. Adiponectin gene polymorphism rs2241766 T/G is associated with response to pioglitazone treatment in type 2 diabetic patients from Southern China. PLoS ONE 2014, 9, e112480. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.F. Biomarkers of adiponectin: Plasma protein variation and genomic DNA polymorphisms. Biomarker Insights 2009, 4, 123–133. [Google Scholar] [PubMed]
- Barth, N.; Langmann, T.; Schölmerich, J.; Schmitz, G.; Schäffler, A. Identification of regulatory elements in the human adipose most abundant gene transcript-1 (apM-1) promoter: Role of SP1/SP3 and TNF-α as regulatory pathways. Diabetologia 2002, 45, 1425–1433. [Google Scholar] [PubMed]
- Zhang, D.; Ma, J.; Brismar, K.; Efendic, S.; Gu, H.F. A single nucleotide polymorphism alters the sequence of SP1 binding site in the adiponectin promoter region and is associated with diabetic nephropathy among type 1 diabetic patients in the Genetics of Kidneys in Diabetes Study. J. Diabetes Complications 2009, 23, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.F.; Long, K.Z.; Hsu, C.-C.; al Mamun, A.; Chiu, Y.-F.; Tu, H.-P.; Chen, P.-S.; Jhang, H.-R.; Hwang, S.-J.; Huang, M.-C. Adiponectin gene (ADIPOQ) polymorphisms correlate with the progression of nephropathy in Taiwanese male patients with type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Jennen, D.G.J.; Tholen, E.; Juengst, H.; Kleinwächter, T.; Hölker, M.; Tesfaye, D.; Ün, G.; Schreinemachers, H.-J.; Murani, E.; et al. A genome scan reveals QTL for growth, fatness, leanness and meat quality in a Duroc‐Pietrain resource population. Anim. Genet. 2007, 38, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Ren, J.; Yang, K.; Ma, J.; Zhang, Z.; Huang, L. Quantitative trait loci for fatty acid composition in longissimus dorsi and abdominal fat: Results from a White Duroc × Erhualian intercross F2 population. Anim. Genet. 2009, 40, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, J.; Flisikowska, T.; Schnieke, A.; Kind, A.; Szydlowski, M.; Switonski, M.; Flisikowski, K. Polymorphisms in the promoter region of the adiponectin (ADIPOQ) gene are presumably associated with transcription level and carcass traits in pigs. Anim. Genet. 2013, 44, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, M.; Li, C.; Xu, Y.; Sun, J.; Lei, C.; Lan, X.; Zhang, C.; Chen, H. Identification and genetic effect of a variable duplication in the promoter region of the cattle ADIPOQ gene. Anim. Genet. 2014, 45, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Kim, D.J.; Lee, J.K.; Lee, C.Y. Genetic relationship between carcass traits and carcass price of Korean cattle. Asian-Australas. J. Anim. Sci. 2010, 23, 848–854. [Google Scholar] [CrossRef]
- Shin, S.; Chung, E. Novel SNPs in the bovine ADIPOQ and PPARGC1A genes are associated with carcass traits in Hanwoo (Korean cattle). Mol. Biol. Rep. 2013, 40, 4651–4660. [Google Scholar] [CrossRef] [PubMed]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012. [Google Scholar] [CrossRef] [PubMed]
- Gaunitz, F.; Heise, K.; Gebhardt, R. A silencer element in the first intron of the glutamine synthetase gene represses induction by glucocorticoids. Mol. Endocrinol. 2004, 18, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Liu, Y.; Liu, Y.; Fang, F.; Wang, X.; Li, B. Association of four insulin resistance genes with type 2 diabetes mellitus and hypertension in the Chinese Han population. Mol. Biol. Rep. 2014, 41, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.Y.; Liu, J.B.; Zhu, J.L.; Liu, T.G.; Zhang, L.Z.; Zhang, Y.; Lei, C.Z.; Chen, H. Identification of a novel mutation within the goat adiponectin gene and its effect on body weight in Chinese indigenous breeds. Biochem. Genet. 2012, 50, 94–102. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Q.-M.; Zhou, H.-T.; Hu, J.; Luo, Y.-Z.; Hickford, J.G.H. Haplotypes and Sequence Variation in the Ovine Adiponectin Gene (ADIPOQ). Genes 2015, 6, 1230-1241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041230
An Q-M, Zhou H-T, Hu J, Luo Y-Z, Hickford JGH. Haplotypes and Sequence Variation in the Ovine Adiponectin Gene (ADIPOQ). Genes. 2015; 6(4):1230-1241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041230
Chicago/Turabian StyleAn, Qing-Ming, Hui-Tong Zhou, Jiang Hu, Yu-Zhu Luo, and Jon G. H. Hickford. 2015. "Haplotypes and Sequence Variation in the Ovine Adiponectin Gene (ADIPOQ)" Genes 6, no. 4: 1230-1241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041230