
Characterization of Thermotolerant Chitinases Encoded by a Brevibacillus laterosporus Strain Isolated from a Suburban Wetland


Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Collection and Chitinolytic Bacteria Isolation
2.2. Preparation of Colloidal Chitin
2.3. Fermentation and Chitinase Activity Assay
2.4. Bacterial Total DNA Extraction and 16S rRNA Gene Sequencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence a | Product Length (in bp) | |
|---|---|---|
| 27F | aga gtt tga tcc tgg ctc | — |
| 1492R | ggt tac ctt gtt acg act | |
| chiA cloning | atg aaa aga ttt ttc ntc atg gc | 1950 |
| cta tga ggt tcc tgt aat kgg tg | ||
| chiC cloning | atg tat caa cac att cct act g | 2574 |
| tta ctt atc agt aac cgc ata t | ||
| ChiA expression | gcc gag ctc gag aac agg agc aac tcc tc | 1875 |
| ccg ctc gag cta tga ggt tcc tgt aat kgg tg | ||
| ChiC expression | gcc gag ctc tcg ycg atc tgc tca gct g | 2460 |
| ccg ctc gag tta ctt atc agt aac cgc ata ttt ttt g |
2.5. Heterologous Expression of Chitinase from B. lateroporus M64
2.6. Purification of Chitinases and Their Characterization
2.7. Nucleotide Sequence Accession Numbers
3. Results
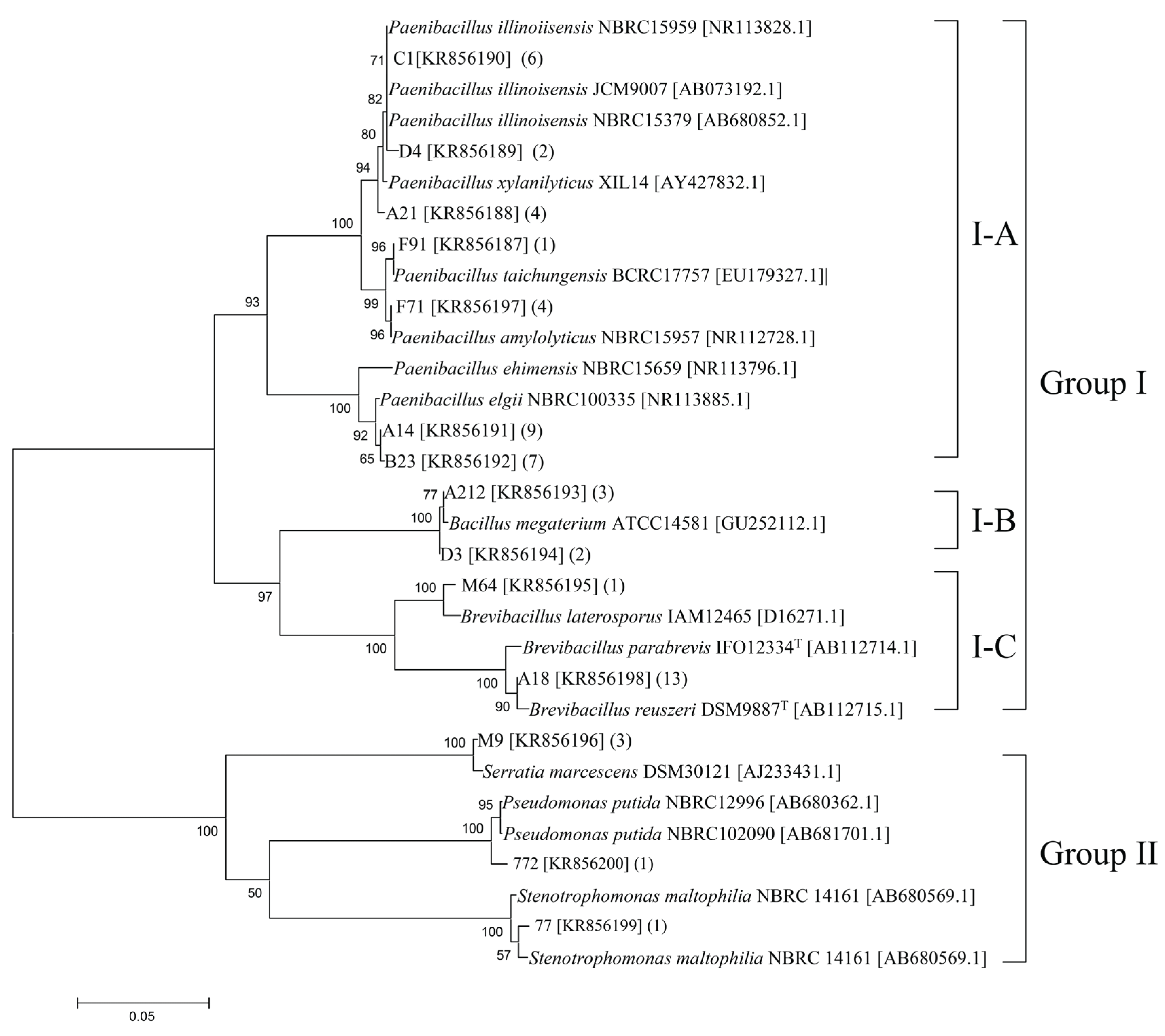
3.1. Culturable Chitinolytic Bacteria Isolated from Wetland Samples and Phylogenetic Classification

3.2. Chitinolytic Activities
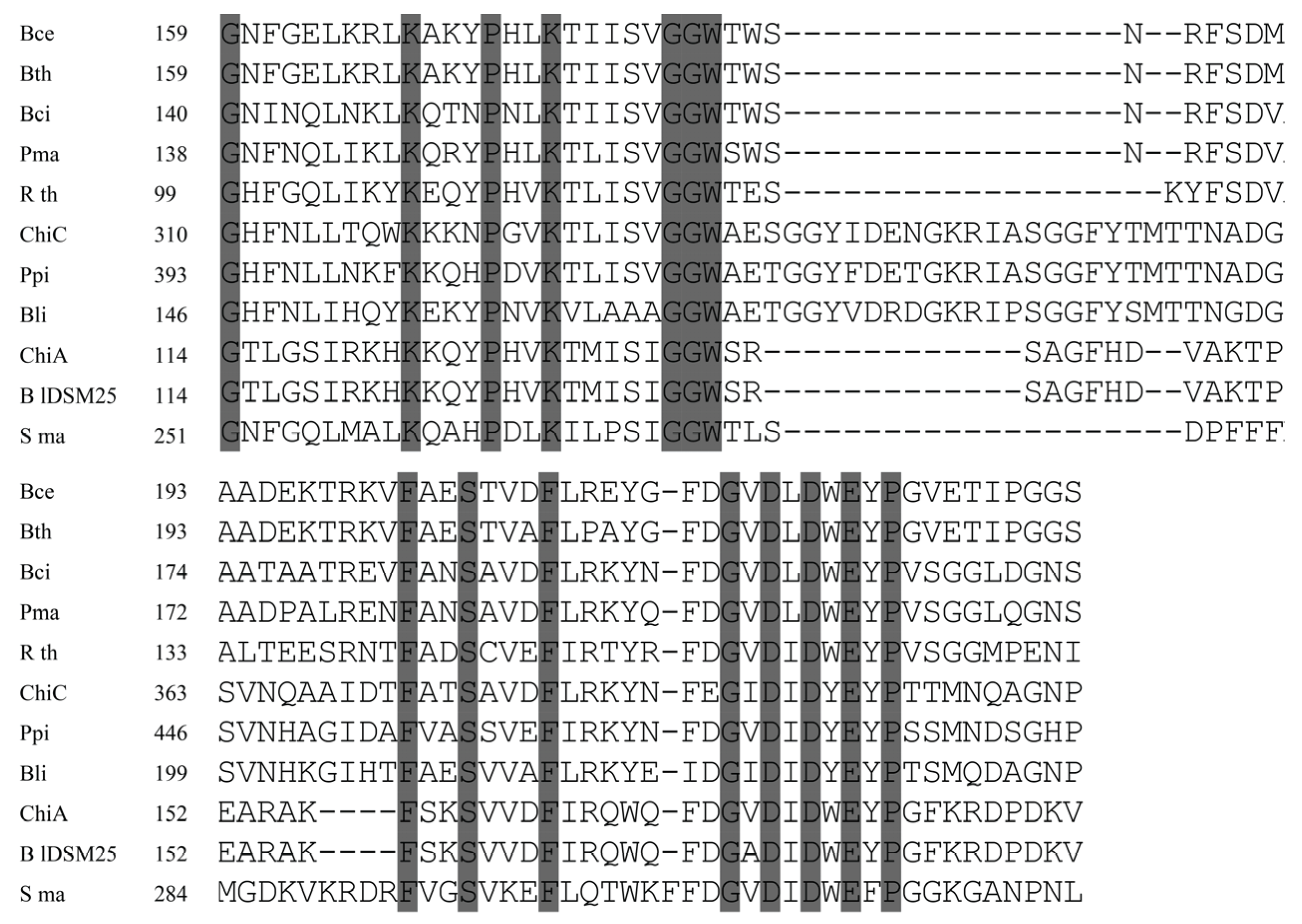
3.3. Cloning and Molecular Analysis of the chiA and chiC Genes from B. laterosporus M64


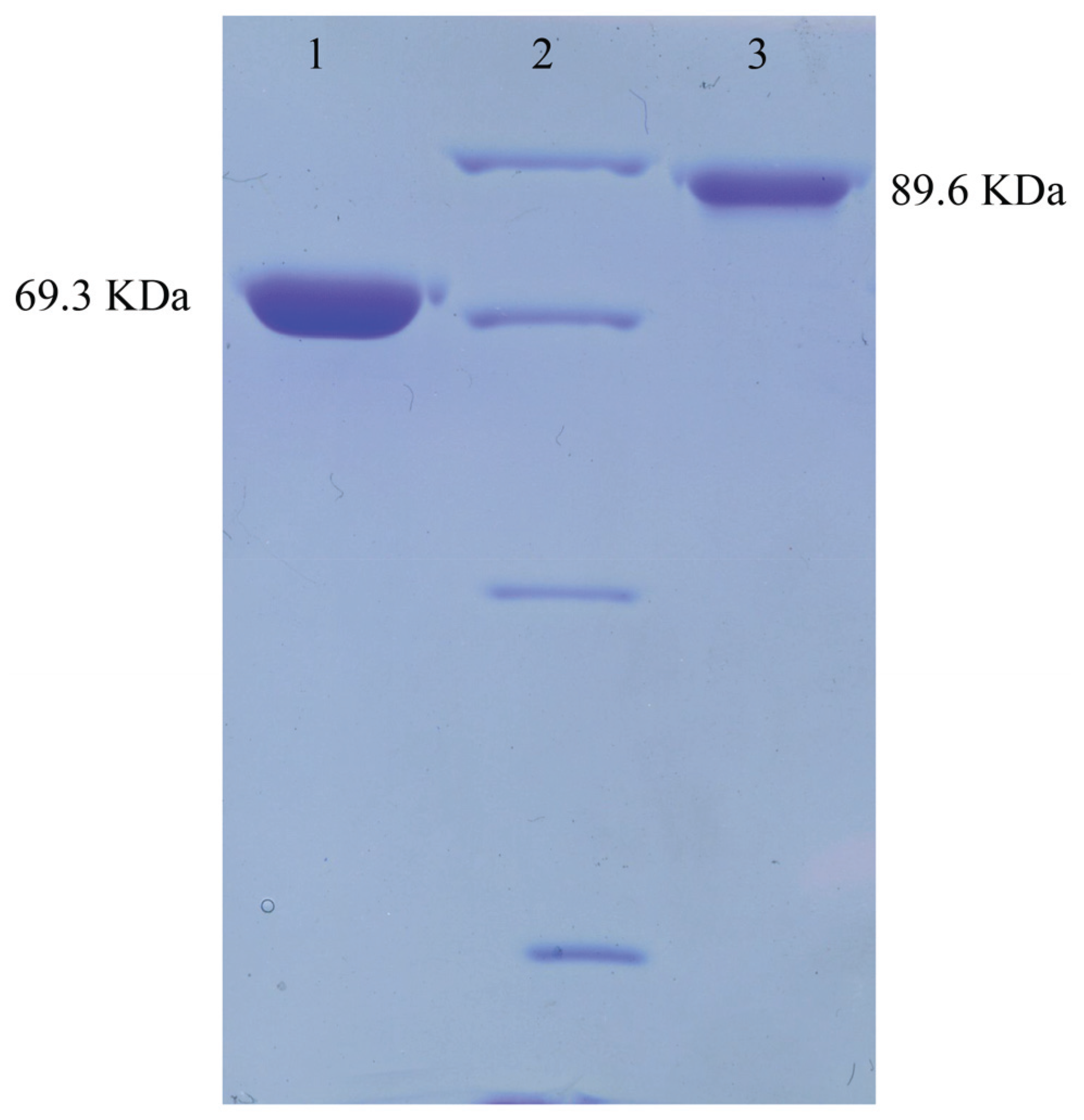
3.4. Purification of ChiA and ChiC

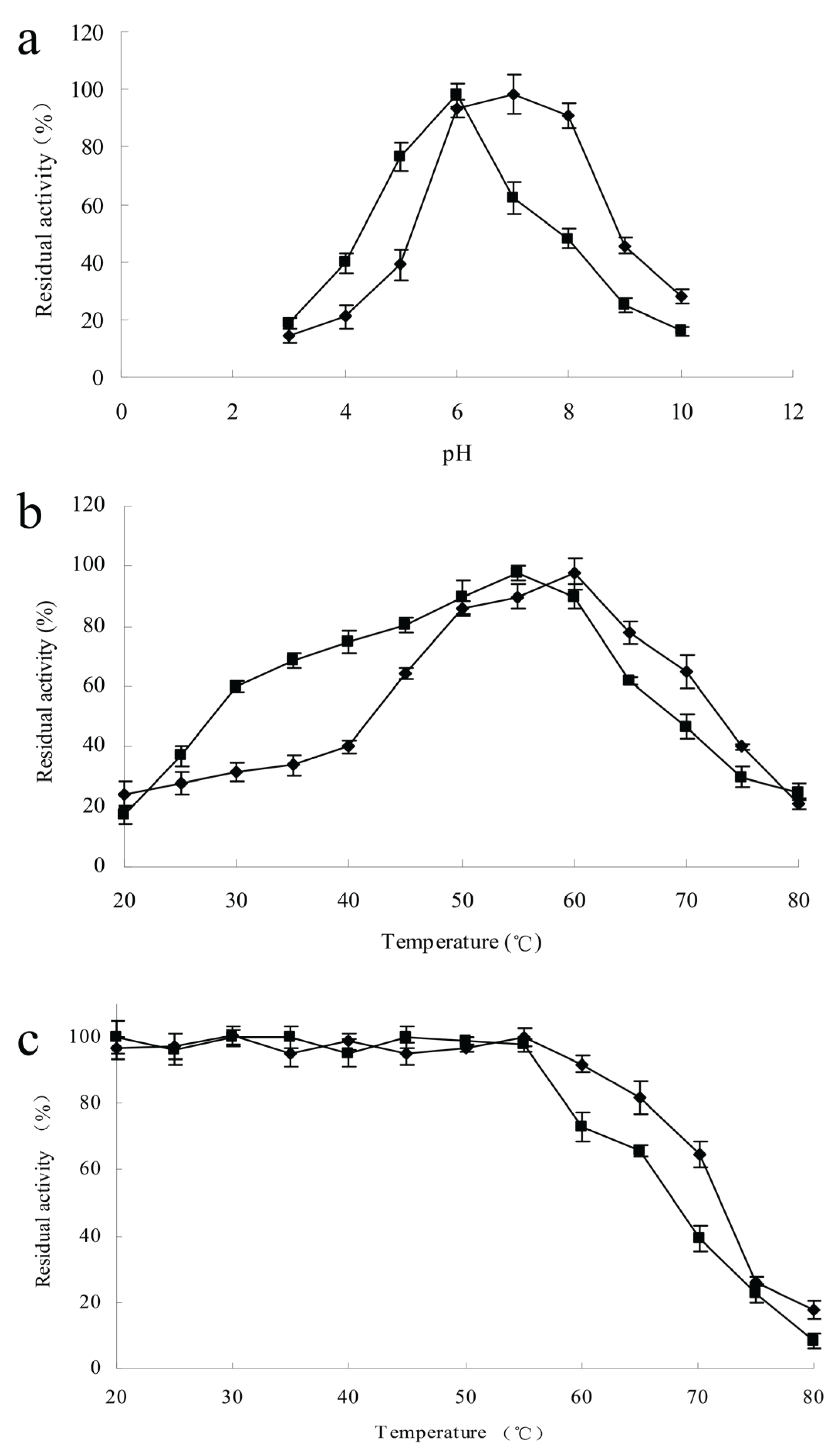
3.5. Effects of Temperature and pH on ChiA and ChiC Activities

3.6. Effects of Metal Ions on the Activities of Purified Enzymes
| Metal Ion | Percent Relative Activity (in Percentage) | |
|---|---|---|
| ChiA | ChiC | |
| Mg2+ | 98.83 ± 1.7 | 101.17 ± 1.8 |
| K+ | 103.75 ± 2.1 | 97.82 ± 1.1 |
| Ca2+ | 113.85 ± 0.9 | 115.90 ± 1.4 |
| Fe2+ | 105.38 ± 2.6 | 109.82 ± 1.9 |
| Fe3+ | 93.13 ± 1.1 | 96.65 ± 3.1 |
| Cu2+ | 81.86 ± 3.8 | 66.79 ± 1.8 |
| Hg2+ | 21.50 ± 3.0 | 13.34 ± 2.5 |
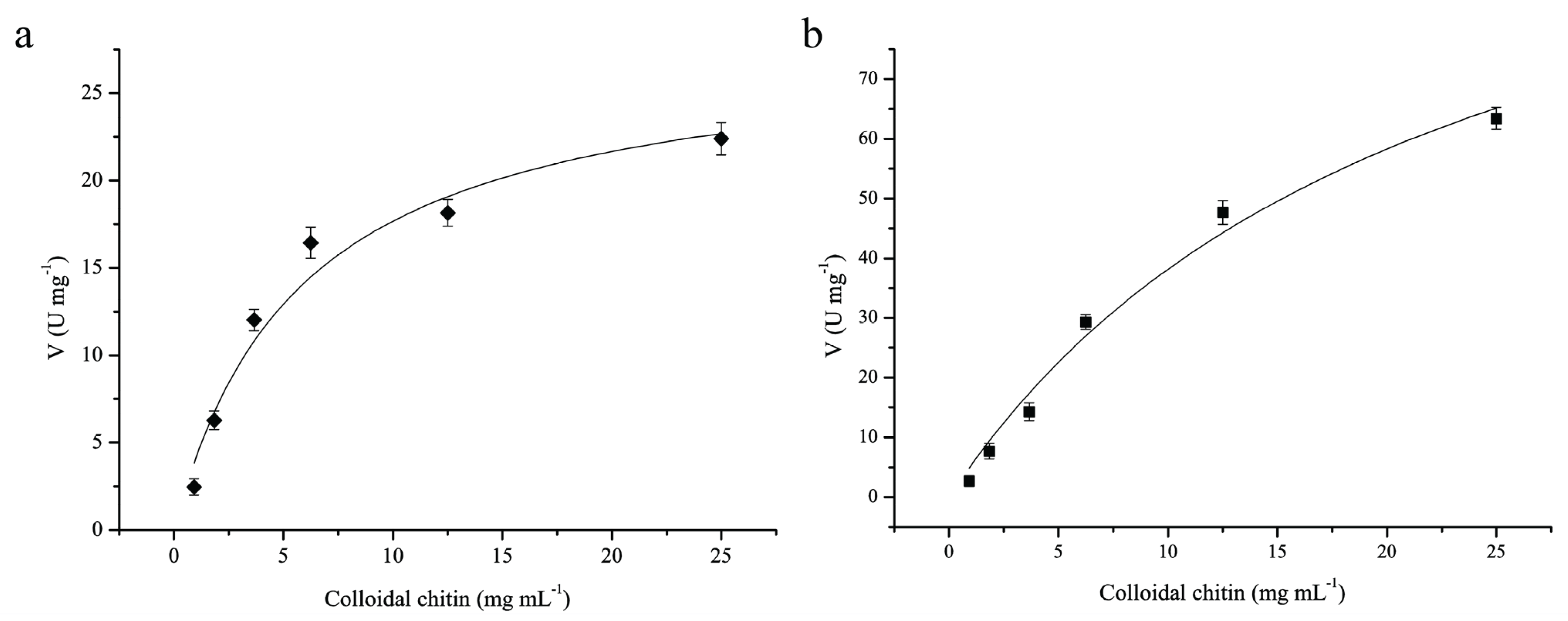
3.7. Kinetic Parameters of ChiA and ChiC

4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mereta, S.T.; Boets, P.; Bayih, A.A.; Malu, A.; Ephrem, Z.; Sisay, A.; Endale, H.; Yitbarek, M.; Jemal, A.; De Meester, L.; et al. Analysis of environmental factors determining the abundance and diversity of macroinvertebrate taxa in natural wetlands of southwest Ethiopia. Ecol. Inform. 2012, 7, 52–61. [Google Scholar] [CrossRef]
- Adrangi, S.; Faramarzi, M.A. From bacteria to human: A journey into the world of chitinase. Biotechnol. Adv. 2013, 31, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Gutknecht, J.L.M.; Goodman, R.M.; Balser, T.C. Linking soil process and microbial ecology in freshwater wetland ecosystem. Plant Soil 2006, 289, 17–34. [Google Scholar] [CrossRef]
- Manjeet, K.; Purushotham, P.; Neeraja, C.; Podile, A.R. Bacterial chitin binding proteins show differential substrate binding and synergy with chitinase. Microbiol. Res. 2013, 168, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Oyanagi, W.; Suzuki, K.; Ohnishi, K.; Tanaka, H. Structure of the gene encoding chitinase D of Bacillus circulans WL-12 and possible homology of the enzyme to other prokaryotic chitinase and class III plant chitinase. J. Bacteriol. 1992, 174, 408–414. [Google Scholar] [PubMed]
- Miyashita, K.; Fuji, T.; Sawada, Y. Molecular cloning and characterization of chitinase gene from Streptomyces lividans. J. Gen. Microbiol. 1991, 137, 2065–2072. [Google Scholar] [CrossRef]
- Ueda, M.; Fujiwara, A.; Kawaguchi, T.; Arai, M. Purification and some properties of six chitinases from Aeromonas sp. No. 10S-24. Biosci. Biotechnol. Biochem. 1995, 59, 2162–2164. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Taiyoji, M.; Sugawara, N.; Nikaidou, N.; Henrissat, B.; Watanabe, T. The third chitinase gene (chiC) of Serratia marcescens 2170 and the relationship of its product to other bacterial chitinase. Biochem. J. 1999, 343, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Sugawara, N.; Suzuki, M.; Uchiyama, T.; Katouno, F.; Nikaidou, N. Chitinase A, B, and C1 of Serratia marcescens 2170 produced by recombinant Escherichia coli: Enzymatic properties and synergim on chitin degradation. Biosci. Biotechnol. Biochem. 2002, 66, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, N.; Tewari, R.; Hoondal, G.S. Biotechnological aspects of chitinolytic enzymes: A review. Appl. Microbiol. Biotechnol. 2006, 71, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Hoster, F.; Schmitz, J.E.; Daniel, R. Enrichment of chitinolytic microorganisms: Isolation and characterization of a chitinase exhibiting antifungal activity against phytopathogenic fungi from a novel Streptomyces strain. Appl. Microbiol. Biotechnol. 2005, 66, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Yu, H.; Cheng, J.; Miao, L. Community composition and cellulase activity of cellulolytic bacteria from forest soils planted with broad-leaved deciduous and evergreen trees. Appl. Microbiol. Biotechnol. 2014, 98, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.K.; Selitrennikoff, C.P. Plant and bacterial chitinases differ in antifungal activity. J. Gen. Microbiol. 1988, 134, 169–176. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrisalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudlry, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijine, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, L.; Eijsink, V.G.; Meadow, R.; Gåseidnes, S. A novel strain of Brevibacillus laterosporus produces chitinases that contribute to its biocontrol potential. Appl. Microbiol. Biotechnol. 2013, 97, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Laribi-Habchi, H.; Bouanane-Darenfed, A.; Drouiche, N.; Pauss, A.; Mameri, N. Purification, characterization, and molecular cloning of an extracellular chitinase from Bacillus licheniformis strain LHH100 isolated wastewater samples in Algeria. Int. J. Biol. Macromol. 2015, 72, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kobori, K.; Miyashita, K.; Fuji, T.; Sakai, H.; Uchida, M.; Tanaka, H. Identification of glutamic acid 204 and aspartic acid 200 in chitinase A1 of Bacillus circulans WL-12 as essential residues for chitinase activity. J. Biol. Chem. 1993, 268, 18567–18572. [Google Scholar] [PubMed]
- Da Silva, A.F.; GarcÍa-Fraga, G.; LÓpez-Seijas, J.; Sieiro, C. Characterization and optimization of heterologous expression in Escherichia coli of the chitinase encoded by the chiA gene of Bacillus halodurans C-125. Process Biochem. 2014, 49, 1622–1629. [Google Scholar] [CrossRef]
- Jee, J.G.; Ikegami, T.; Hashimoto, M.; Kawabata, T.; Ikeguchi, M.; Watanabe, T.; Shirakawa, M. Solution structure of the fibronectin type Ⅲ domain from Bacillus circulans WL-12 chitinase A1. J. Biol. Chem. 2002, 277, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Peterbauer, C.; Hayes, C.K.; Harman, G.E. Synergistic interaction between fungal cell wall degrading enzymes and different antifungal compounds enhances inhibition of spore germination. Microbiology 1994, 140, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Pardo-lópez, L.; Muñoz-Garay, C.; Porta, H.; Rodríguez-Almazán, C.; Soberón, M.; Bravo, A. Strategies to improve the insecticidal activity of Cry toxins from Bacillus thuringiensis. Peptides 2009, 30, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Someya, N.; Nakajima, K.; Hirayae, T.; Hibi, T.; Akutsu, K. Synergistic antifungal activity of chitinolytic enzymes and prodigiosin produced by biocontrol bacterium, Serratia marcescens strain B2, against gray mold pathogen, Botrytis cinerea. J. Gen. Plant Pathol. 2001, 67, 312–317. [Google Scholar] [CrossRef]
- Felse, P.A.; Panda, T. Regulation and cloning of microbial chitinase gene. Appl. Microbiol. Biotechnol. 1999, 51, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Das, S.N.; Sarma, P.V.S.R.N.; Neeraja, C.; Malati, N.; Podile, A.R. Members of Gammaproteobacteria and Bacilli represent the culturable diversity of chitinolytic bacteria in chitin-enriched soils. World J. Microbiol. Biotechnol. 2010, 26, 1875–1881. [Google Scholar] [CrossRef]
- Someya, N.; Ikeda, S.; Morohoshi, T.; Tsujimoto, M.N.; Yoshida, T.; Sawada, H. Diversity of culturable chitinolytic bacteria from rhizospheres of agronomic plants in Japan. Microbes Environ. 2011, 26, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ruiu, L. Brevibacillus laterosporus, a pathogen of invertebrates and a broad-spectrum antimicrobial species. Insects 2013, 4, 476–492. [Google Scholar] [CrossRef] [PubMed]
- Terahara, T.; Ikeda, S.; Noritake, C.; Minamisawa, K.; Ando, K.; Tsuneda, S. Molecular diversity of bacterial chitinase in arable soils and the effects of environmental factors on the chitinolytic bacterial community. Soil Biol. Biochem. 2009, 41, 473–480. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Tan, H.; Chi, N.; Zhang, Q.; Du, Y.; Yin, H. Characterization of a chitinase from Pseudoalteromonas sp. DL-6, a marine psychrophilic bacterium. Int. J. Biol. Macromol. 2014, 70, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Greene, L.H. Sequence and structural analysis of the chitinase insertion domain reveals two conserved motifs involved in chitin-binding. PLoS One 2010, 5, e8654. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.A.; Nguyen, T.H.; Nguyen, T.T.; Peterbauer, C.K.; Mathiesen, G.; Haltrich, D. Chitinase from Bacillus licheniformis DSM13: Expression in Lactobacillus plantarum WCFS1 and biochemical characterization. Protein Expr. Purif. 2012, 81, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Kudan, S.; Pichyanqkura, R. Purification and characterization of thermostable chitinase from Bacillus licheniformis SK-1. Appl. Biochem. Biotechnol. 2009, 157, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Karthik, N.; Binod, P.; Pandev, A. Purification and characterization of an acidic and antifungal chitinase produced by a Streptomyces sp. Bioresour. Technol. 2015, 188, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hsu, M.; Jiang, S. Purification and characterization of an exo-N,N'-diacetylchitobiohydrolase-like enzyme from Cellulomonas flavigena NTOU1. Enzyme Microb. Techol. 1997, 20, 191–197. [Google Scholar] [CrossRef]
- Pradeep, G.C.; Choi, Y.H.; Choi, Y.S.; Suh, S.E.; Seong, J.H.; Cho, S.S.; Bae, M.S.; Yoo, J.C. An extremely alkaline novel chitinase from Streptomyces sp. CS495. Process Biochem. 2014, 49, 223–229. [Google Scholar] [CrossRef]
- Suma, K.; Podile, A.R. Chitinase A from Stenotrophmonas maltophilia shows transglycosylation and antifungal activities. Bioresour. Technol. 2013, 133, 213–220. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Cheng, D.; Miao, L. Characterization of Thermotolerant Chitinases Encoded by a Brevibacillus laterosporus Strain Isolated from a Suburban Wetland. Genes 2015, 6, 1268-1282. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041268
Liu P, Cheng D, Miao L. Characterization of Thermotolerant Chitinases Encoded by a Brevibacillus laterosporus Strain Isolated from a Suburban Wetland. Genes. 2015; 6(4):1268-1282. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041268
Chicago/Turabian StyleLiu, Pulin, Deyong Cheng, and Lihong Miao. 2015. "Characterization of Thermotolerant Chitinases Encoded by a Brevibacillus laterosporus Strain Isolated from a Suburban Wetland" Genes 6, no. 4: 1268-1282. https://0-doi-org.brum.beds.ac.uk/10.3390/genes6041268