
Metabolic Genes within Cyanophage Genomes: Implications for Diversity and Evolution

Abstract
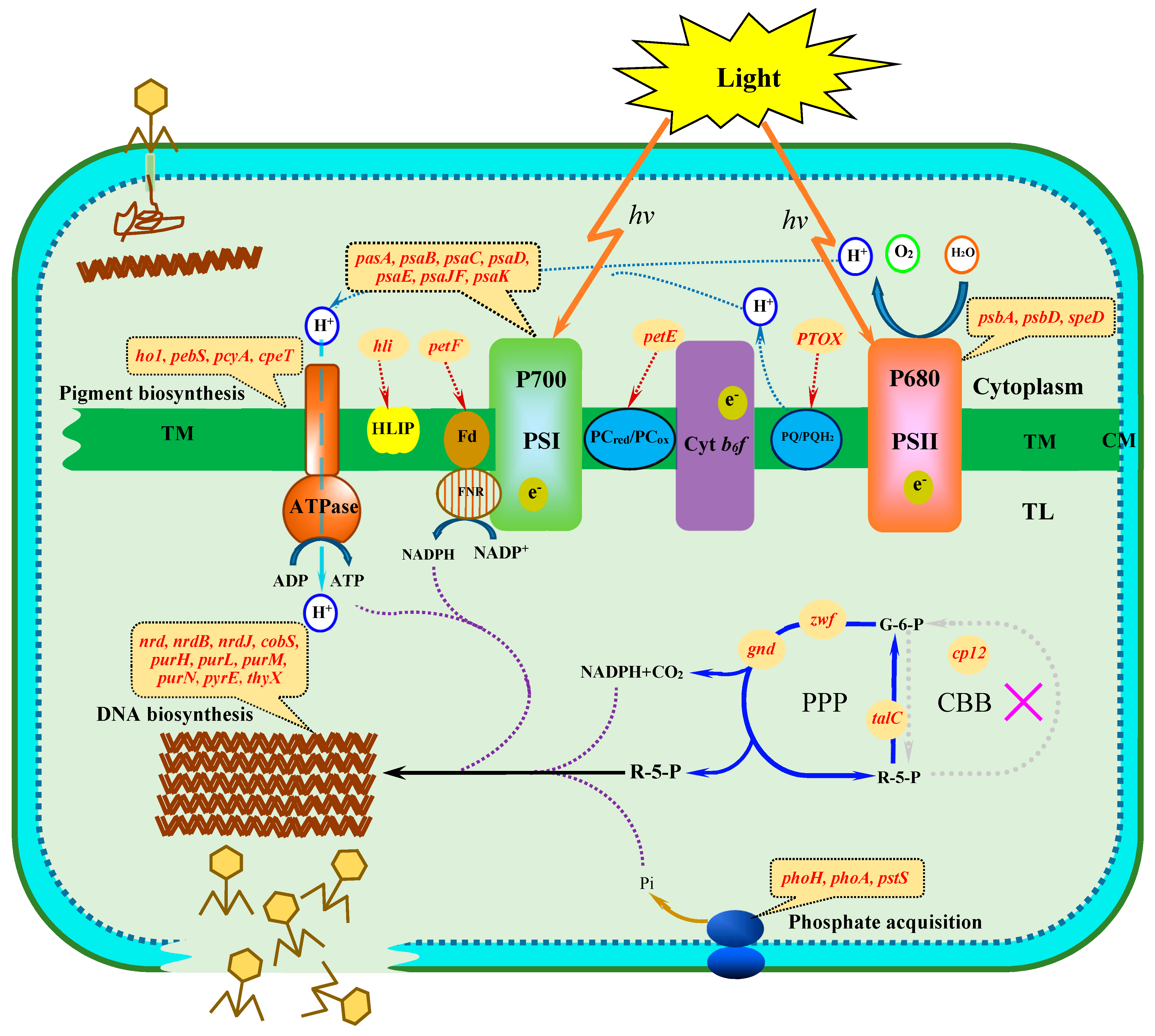
:1. Introduction
2. Origin of Host-Derived Metabolic Genes
3. Physiological Functions of Host-Derived Metabolic Genes
3.1. Photosynthetic Membrane
3.2. Phycobilisome Degradation
3.3. Carbon Metabolism
3.4. Phosphate Acquisition
3.5. DNA Biosynthesis
4. Signature Markers for Cyanophage Evolution
5. Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Drewes, F.; Peter, H.; Sommarugaa, R. Are viruses important in the plankton of highly turbid glacier-fed lakes? Sci. Rep. 2016, 6, 24608. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, J.A. Marine viruses: Biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Hambly, E.; Suttle, C.A. The viriosphere, diversity, and genetic exchange within phage communities. Curr. Opin. Microbiol. 2005, 8, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A. Marine viruses-major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Buckling, A.; Rainey, P.B. Antagonistic coevolution between a bacterium and a bacteriophage. Proc. Biol. Sci. 2002, 269, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Marston, M.F.; Pierciey, F.J.; Shepard, A.; Gearin, G.; Qi, J.; Yandava, C.; Schuster, S.C.; Henn, M.R.; Martiny, J.B. Rapid diversification of co-evolving marine Synechococcus and a virus. Proc. Natl. Acad. Sci. USA 2012, 109, 4544–4549. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Riemann, L.; Marston, M.F.; Middelboe, M. Antagonistic co-evolution of marine planktonic viruses and their hosts. Ann. Rev. Mar. Sci. 2014, 6, 393–414. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Yoshida, T.; Kashima, A.; Takashima, Y.; Hosoda, N.; Nagasaki, K.; Hiroishi, S. Ecological dynamics of the toxic bloom-forming cyanobacterium Microcystis aeruginosa and its cyanophages in freshwater. Appl. Environ. Microbiol. 2008, 74, 3269–3273. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Jaskulska, A.; Pawełczyk, J.; Gągała, I.; Serwecińska, L.; Dziadek, J. Cyanophages infection of Microcystis bloom in lowland dam reservoir of Sulejów, Poland. Microb. Ecol. 2016, 71, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Lu, J. Genomic sequence and evolution of marine cyanophage P60: A new insight on lytic and lysogenic phages. Appl. Environ. Microbiol. 2002, 68, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Mann, N.H.; Clokie, M.R.; Millard, A.; Cook, A.; Wilson, W.H.; Wheatley, P.J.; Letarov, A.; Krisch, H.M. The genome of S-PM2, a “photosynthetic” T4-type bacteriophage that infects marine Synechococcus strains. J. Bacteriol. 2005, 187, 3188–3200. [Google Scholar] [CrossRef] [PubMed]
- Pope, W.H.; Weigele, P.R.; Chang, J.; Pedulla, M.L.; Ford, M.E.; Houtz, J.M.; Jiang, W.; Chiu, W.; Hatfull, G.F.; Hendrix, R.W.; et al. Genome sequence, structural proteins, and capsid organization of the cyanophage Syn5: A ‘horned’ bacteriophage of marine Synechococcus. J. Mol. Biol. 2007, 368, 966–981. [Google Scholar] [CrossRef] [PubMed]
- Weigele, P.R.; Pope, W.H.; Pedulla, M.L.; Houtz, J.M.; Smith, A.L.; Conway, J.F.; King, J.; Hatfull, G.F.; Lawrence, J.G.; Hendrix, R.W. Genomic and structural analysis of Syn9, a cyanophage infecting marine Prochlorococcus and Synechococcus. Environ. Microbiol. 2007, 9, 1675–1695. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Krastins, B.; Hughes, J.L.; Kelly, L.; Chase, M.; Sarracino, D.; Chisholm, S.W. The genome and structural proteome of an ocean cyanobacterial siphovirus: A new window into the cyanobacterial ‘mobilome’. Environ. Microbiol. 2009, 11, 2935–2951. [Google Scholar] [CrossRef] [PubMed]
- Proctor, L.M.; Fuhrman, J.A. Viral mortality of marine bacteria and cyanobacteria. Nature 1990, 343, 60–62. [Google Scholar] [CrossRef]
- Suttle, C.A. Cyanophages and their role in the ecology of cyanobacteria. In The Ecology of Cyanobacteria. Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 563–589. [Google Scholar]
- Suttle, C.A.; Chan, A.M.; Cottrell, M.T. Infection of phytoplankton by viruses and reduction of primary productivity. Nature 1990, 347, 467–469. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chan, A.M. Marine cyanophages infecting oceanic and coastal strains of Synechococcus: Abundance, morphology, cross-infectivity and growth characteristics. Mar. Ecol. Prog. Ser. 1993, 92, 99–109. [Google Scholar] [CrossRef]
- Mühling, M.; Fuller, N.J.; Millard, A.; Somerfield, P.J.; Marie, D.; Wilson, W.H.; Scanlan, D.J.; Post, A.F.; Joint, I.; Mann, N.H. Genetic diversity of marine Synechococcus and co-occurring cyanophage communities: Evidence for viral control of phytoplankton. Environ. Microbiol. 2005, 7, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wommack, K.E.; Chen, F. Abundance and distribution of Synechococcus spp. and cyanophages in the Chesapeake Bay. Appl. Environ. Microbiol. 2011, 77, 7459–7468. [Google Scholar] [CrossRef] [PubMed]
- Matteson, A.R.; Rowe, J.M.; Ponsero, A.J.; Pimentel, T.M.; Boyd, P.W.; Wilhelm, S.W. High abundances of cyanomyoviruses in marine ecosystems demonstrate ecological relevance. FEMS Microbiol. Ecol. 2013, 84, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; Fuller, N.J.; Joint, I.R.; Mann, N.H. Analysis of cyanophage diversity and population structure in a south-north transect of the Atlantic ocean. Bull. Inst. Océanogr. Monaco 1999, 19, 209–216. [Google Scholar]
- Dorigo, U.; Jacquet, S.; Humbert, J.F. Cyanophage diversity, inferred from g20 gene analyses, in the largest natural lake in France, Lake Bourget. Appl. Environ. Microbiol. 2004, 70, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Marston, M.F.; Sallee, J.L. Genetic diversity and temporal variation in the cyanophage community infecting marine Synechococcus species in Rhode Island’s coastal waters. Appl. Environ. Microbiol. 2003, 69, 4639–4647. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Millard, A.D.; Mehta, J.Y.; Mann, N.H. Virus isolation studies suggest short-term variations in abundance in natural cyanophage populations of the Indian Ocean. J. Mar. Biol. Assoc. UK 2006, 86, 499–505. [Google Scholar] [CrossRef]
- Millard, A.D.; Mann, N.H. A temporal and spatial investigation of cyanophage abundance in the Gulf of Aqaba, Red Sea. J. Mar. Biol. Assoc. UK 2006, 86, 507–515. [Google Scholar] [CrossRef]
- Ortmann, A.C.; Lawrence, J.E.; Suttle, C.A. Lysogeny and lytic viral production during a bloom of the cyanobacterium Synechococcus spp. Microb. Ecol. 2002, 43, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; Joint, I.R.; Carr, N.G.; Mann, N.H. Isolation and Molecular Characterization of Five Marine Cyanophages Propagated on Synechococcus sp. Strain WH7803. Appl. Environ. Microbiol. 1993, 59, 3736–3743. [Google Scholar] [PubMed]
- Tucker, S.; Pollard, P. Identification of cyanophage Ma-LBP and infection of the cyanobacterium Microcystis aeruginosa from an Australian subtropical lake by the virus. Appl. Environ. Microbiol. 2005, 71, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Takashima, Y.; Tomaru, Y.; Shirai, Y.; Takao, Y.; Hiroishi, S.; Nagasaki, K. Isolation and characterization of a cyanophage infecting the toxic cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 2006, 72, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.B.; Yuan, X.P.; Li, R.H.; Zhang, Q.Y. Isolation of a novel cyanophage infectious to the filamentous cyanobacterium Planktothrix agardhii (Cyanophyceae) from Lake Donghu, China. Aquat. Microb. Ecol. 2009, 54, 163–170. [Google Scholar] [CrossRef]
- Yoshida, T.; Nagasaki, K.; Takashima, Y.; Shirai, Y.; Tomaru, Y.; Takao, Y.; Sakamoto, S.; Hiroishi, S.; Ogat, H. Ma-LMM01 Infecting Toxic Microcystis aeruginosa Illuminates Diverse Cyanophage Genome Strategies. J. Bacteriol. 2008, 190, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.B.; Gui, J.F.; Zhang, Q.Y. A novel cyanophage with a cyanobacterial nonbleaching protein A gene in the genome. J. Virol. 2012, 86, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, M.; Kong, S.; Gao, Y.; An, C. Cyanophage Pf-WMP4, a T7-like phage infecting the freshwater cyanobacterium Phormidium foveolarum: Complete genome sequence and DNA translocation. Virology 2007, 366, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kong, S.; Shi, M.; Fu, L.; Gao, Y.; An, C. Genomic analysis of freshwater cyanophage Pf-WMP3 Infecting cyanobacterium Phormidium foveolarum: The conserved elements for a phage. Microb. Ecol. 2008, 56, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Ou, T.; Gao, X.C.; Li, S.H.; Zhang, Q.Y. Genome analysis and gene nblA identification of Microcystis aeruginosa myovirus (MaMV-DC) reveal the evidence for horizontal gene transfer events between cyanomyovirus and host. J. Gen. Virol. 2015, 96, 3681–3697. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.D.; Zwirglmaier, K.; Downey, M.J.; Mann, N.H.; Scanlan, D.J. Comparative genomics of marine cyanomyoviruses reveals the widespread occurrence of Synechococcus host genes localized to a hyperplastic region: Implications for mechanisms of cyanophage evolution. Environ. Microbiol. 2009, 11, 2370–2387. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Waterbury, J.B.; Chisholm, S.W. Cyanophages infecting the oceanic cyanobacterium Prochlorococcus. Nature 2003, 424, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Coleman, M.L.; Weigele, P.; Rohwer, F.; Chisholm, S.W. Three Prochlorococcus Cyanophage Genomes: Signature Features and Ecological Interpretations. PLoS Biol. 2005, 3, e144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.B.; Huang, K.H.; Ignacio-Espinoza, J.C.; Berlin, A.M.; Kelly, L.; Weigele, P.R.; DeFrancesco, A.S.; Kern, S.E.; Thompson, L.R.; Young, S.; et al. Genomic analysis of oceanic cyanobacterial myoviruses compared with T4-like myoviruses from diverse hosts and environments. Environ. Microbiol. 2010, 12, 3035–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitbart, M.; Thompson, L.R.; Suttle, C.A.; Sullivan, M.B. Exploring the vast diversity of marine viruses. Oceanography 2007, 20, 135–139. [Google Scholar] [CrossRef]
- Sharon, I.; Alperovitch, A.; Rohwer, F.; Haynes, M.; Glaser, F.; Atamna-Ismaeel, N.; Pinter, R.Y.; Partensky, F.; Koonin, E.V.; Wolf, Y.I.; et al. Photosystem I gene cassettes are present in marine virus genomes. Nature 2009, 461, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Puxty, R.J.; Perez-Sepulveda, B.; Rihtman, B.; Evans, D.J.; Millard, A.D.; Scanlan, D.J. Spontaneous deletion of an “ORFanage” region facilitates host adaptation in a “photosynthetic” cyanophage. PLoS ONE 2015, 10, e0132642. [Google Scholar] [CrossRef] [PubMed]
- Alperovitch-Lavy, A.; Sharon, I.; Rohwer, F.; Aro, E.M.; Glaser, F.; Milo, R.; Nelson, N.; Béjà, O. Reconstructing a puzzle: Existence of cyanophages containing both photosystem-I and photosystem-II gene suites inferred from oceanic metagenomic datasets. Environ. Microbiol. 2011, 13, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Mann, N.H.; Cook, A.; Millard, A.; Bailey, S.; Clokie, M. Marine ecosystems: Bacterial photosynthesis genes in a virus. Nature 2003, 424, 741. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.; Clokie, M.R.J.; Shub, D.A.; Mann, N.H. Genetic organization of the psbAD region in phages infecting marine Synechococcus strains. Proc. Natl. Acad. Sci. USA 2004, 101, 11007–11012. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Sullivan, M.B.; Johnson, Z.I.; Tolonen, A.C.; Rohwer, F.; Chisholm, S.W. Transfer of photosynthesis genes to and from Prochlorococcus viruses. Proc. Natl. Acad. Sci. USA 2004, 101, 11013–11018. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Jaffe, J.D.; Johnson, Z.I.; Church, G.M.; Chisholm, S.W. Photosynthesis genes in marine viruses yield proteins during host infection. Nature 2005, 438, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Shan, J.; Bailey, S.; Jia, Y.; Krisch, H.M. Transcription of a ‘photosynthetic’ T4-type phage during infection of a marine cyanobacterium. Environ. Microbiol. 2006, 8, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Puxty, R.J.; Millard, A.D.; Evans, D.J.; Scanlan, D.J. Viruses inhibit CO2 fixation in the most abundant phototrophs on earth. Curr. Biol. 2016, 26, 1585–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, I.; Tzahor, S.; Williamson, S.; Shmoish, M.; Man-Aharonovich, D.; Rusch, D.B.; Yooseph, S.; Zeidner, G.; Golden, S.S.; Mackey, S.R.; et al. Viral photosynthetic reaction center genes and transcripts in the marine environment. ISME J. 2007, 1, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.R.; Zeng, Q.L.; Kelly, L.; Huang, K.H.; Singer, A.U.; Stubbe, J.; Chisholm, S.W. Phage auxiliary metabolic genes and the redirection of cyanobacterial host carbon metabolism. Proc. Natl. Acad. Sci. USA 2011, 108, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, B.L.; U’Ren, J.M. Viral metabolic reprogramming in marine ecosystems. Curr. Opin. Microbiol. 2016, 31, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Deveau, H.; Barrangou, R.; Garneau, J.E.; Labonte, J.; Fremaux, C.; Boyaval, P.; Romero, D.A.; Horvath, P.; Moineau, S. Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J. Bacteriol. 2007, 190, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Lindell, D.; Lee, J.A.; Thompson, L.R.; Bielawski, J.P.; Chisholm, S.W. Prevalence and evolution of core photosystem II genes in marine cyanobacterial viruses and their hosts. PLoS Biol. 2006, 4, 1344–1357. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.J.; Rusch, D.B.; Yooseph, S.; Halpern, A.L.; Heidelberg, K.B.; Glass, J.I. The Sorcerer II Global Ocean Sampling Expedition: Metagenomic characterization of viruses within aquatic microbial samples. PLoS ONE 2008, 3, e1456. [Google Scholar] [CrossRef] [PubMed]
- Zeidner, G.; Bielawski, J.P.; Shmoish, M.; Scanlan, D.J.; Sabehi, G.; Béjà, O. Potential photosynthesis gene recombination between between Prochlorococcus and Synechococcus via viral intermediates. Environ. Microbiol. 2005, 7, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Ignacio-Espinoza, J.C.; Sullivan, M.B. Phylogenomics of T4 cyanophages: Lateral gene transfer in the ‘core’ and origins of host genes. Environ. Microbiol. 2012, 14, 2113–2126. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.D.; Gierga, G.; Clokie, M.R.J.; Evans, D.J.; Hess, W.R.; Scanlan, D.J. An antisense RNA in a lytic cyanophage links psbA to a gene encoding a homing endonuclease. ISME J. 2010, 4, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Marston, M.F.; Amrich, C.G. Recombination and microdiversity in coastal marine cyanophages. Environ. Microbiol. 2009, 11, 2893–2903. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Jaffe, J.D.; Coleman, M.L.; Futschik, M.E.; Axmann, I.M.; Rector, T.; Kettler, G.; Sullivan, M.B.; Steen, R.; Hess, W.R.; et al. Genome-wide expression dynamics of a marine virus and host reveal features of co-evolution. Nature 2007, 449, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Todar, K. Bacteriophage. In Todar’s Online Textbook of Bacteriology; Madison, Wisconsin; Available online: http://textbookofbacteriology.net/phage.html (accessed on 30 August 2016).
- Zeng, Q.; Chisholm, S.W. Marine viruses exploit their host’s two-component regulatory system in response to resource limitation. Curr. Biol. 2012, 22, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Zhang, S.; Jiao, N.; Chen, F. Comparative genomic and phylogenomic analyses reveal a conserved core genome shared by estuarine and oceanic cyanopodoviruses. PLoS ONE 2015, 10, e0142962. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jing, R.; Liu, J.; Yu, Z.; Jin, J.; Liu, X.; Wang, X.; Wang, G. Narrow distribution of cyanophage psbA genes observed in two paddy waters of Northeast China by an incubation experiment. Virol. Sin. 2016, 31, 199–191. [Google Scholar] [CrossRef] [PubMed]
- Adir, N.; Zer, H.; Shochat, S.; Ohad, I. Photoinhibition—A historical perspective. Photosynth. Res. 2003, 76, 343–370. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed]
- Bograh, A.; Gingras, Y.; Tajmir-Riahi, H.A.; Carpentier, R. The effects of spermine and spermidine on the structure of photosystem II proteins in relation to inhibition of electron transport. FEBS Lett. 1997, 402, 41–44. [Google Scholar] [CrossRef]
- Puxty, R.J.; Millard, A.D.; Evans, D.J.; Scanlan, D.J. Shedding new light on viral photosynthesis. Photosynth. Res. 2015, 126, 71–97. [Google Scholar] [CrossRef] [PubMed]
- Sharon, I.; Battchikova, N.; Aro, E.M.; Giglione, C.; Meinnel, T.; Glaser, F.; Pinter, R.Y.; Breitbart, M.; Rohwer, F.; Béjà, O. Comparative metagenomics of microbial traits within oceanic viral communities. ISME J. 2011, 5, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, M. Plastid terminal oxidase and its biological significance. Planta 2004, 218, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.; Millard, A.D.; Mann, N.H. T4 genes in the marine ecosystem: Studies of the T4-like cyanophages and their role in marine ecology. Virol. J. 2010, 7, 291. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.W.; Reijerse, E.J.; Lubitz, W.; Hofmann, E.; Frankenberg-Dinkel, N. Radical mechanism of cyanophage phycoerythrobilin synthase (PebS). Biochem. J. 2011, 433, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Dammeyer, T.; Bagby, S.C.; Sullivan, M.B.; Chisholm, S.W.; Frankenberg-Dinkel, N. Efficient phage-mediated pigment biosynthesis in oceanic cyanobacteria. Curr. Biol. 2008, 18, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Zinser, E.R.; Lindell, D.; Johnson, Z.I.; Futschik, M.E.; Steglich, C.; Coleman, M.L.; Wright, M.A.; Rector, T.; Steen, R.; McNulty, N.; et al. Choreography of the transcriptome, photophysiology, and cell cycle of a minimal photoautotroph, Prochlorococcus. PLoS ONE 2009, 4, e5135. [Google Scholar] [CrossRef] [PubMed]
- Tamoi, M.; Miyazaki, T.; Fukamizo, T.; Shigeoka, S. The Calvin cycle in cyanobacteria is regulated by CP12 via the NAD(H)/NADP(H) ratio under light/dark conditions. Plant J. 2005, 42, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Martiny, A.C.; Coleman, M.L.; Chisholm, S.W. Phosphate acquisition genes in Prochlorococcus ecotypes: Evidence for genome-wide adaptation. Proc. Natl. Acad. Sci. USA 2006, 103, 12552–12557. [Google Scholar] [CrossRef] [PubMed]
- Tetu, S.G.; Brahamsha, B.; Johnson, D.A.; Tai, V.; Phillippy, K.; Palenik, B.; Paulsen, I.T. Microarray analysis of phosphate regulation in the marine cyanobacterium Synechococcus sp. WH8102. ISME J. 2009, 3, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Ding, H.; Huang, K.H.; Osburne, M.S.; Chisholm, S.W. Genetic diversity in cultured and wild marine cyanomyoviruses reveals phosphorus stress as a strong selective agent. ISME J. 2013, 7, 1827–1841. [Google Scholar] [CrossRef] [PubMed]
- Martiny, A.C.; Huang, Y.; Li, W. Occurrence of phosphate acquisition genes in Prochlorococcus cells from different ocean regions. Environ. Microbiol. 2009, 11, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Ding, H.; Zeng, Q. Transcriptomic response during phage infection of a marine cyanobacterium under phosphorus-limited conditions. Environ. Microbiol. 2016, 18, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Dreher, T.W.; Brown, N.; Bozarth, C.S.; Schwartz, A.D.; Riscoe, E.; Thrash, C.; Bennett, S.E.; Tzeng, S.C.; Maier, C.S. A freshwater cyanophage whose genome indicates close relationships to photosynthetic marine cyanomyophages. Environ. Microbiol. 2011, 13, 1858–1874. [Google Scholar] [CrossRef] [PubMed]
- Kazakov, A.E.; Vassieva, O.; Gelfand, M.S.; Osterman, A.; Overbeek, R. Bioinformatics classification and functional analysis of PhoH homologs. In Silico Biol. 2003, 3, 3–15. [Google Scholar] [PubMed]
- Gleason, F.K.; Olszewski, N.E. Isolation of the gene for the B12-dependent ribonucleotide reductase from Anabaena sp. strain PCC 7120 and expression in Escherichia coli. J. Bacteriol. 2002, 184, 6544–6550. [Google Scholar] [CrossRef] [PubMed]
- Hanson, E.; Mathews, C.K. Allosteric effectors are required for subunit association in T4 phage ribonucleotide reductase. J. Biol. Chem. 1994, 269, 30999–31005. [Google Scholar] [PubMed]
- Helliwell, K.E.; Lawrence, A.D.; Holzer, A.; Kudahl, U.J.; Sasso, S.; Kräutler, B.; Scanlan, D.J.; Warren, M.J.; Smith, A.G. Cyanobacteria and eukaryotic algae use different chemical variants of vitamin B12. Curr. Biol. 2016, 26, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Fuller, N.J.; Wilson, W.H.; Joint, I.R.; Mann, N.H. Occurrence of a sequence in marine cyanophages similar to that of T4 g20 and its application to PCR-based detection and quantification techniques. Appl. Environ. Microbiol. 1998, 64, 2051–2060. [Google Scholar] [PubMed]
- Zhong, Y.; Chen, F.; Wilhelm, S.W.; Poorvin, L.; Hodson, R.E. Phylogenetic diversity of marine cyanophage isolates and natural virus communities as revealed by sequences of viral capsid assembly protein gene g20. Appl. Environ. Microbiol. 2002, 68, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Short, C.M.; Suttle, C.A. Nearly identical bacteriophage structural gene sequences are widely distributed in both marine and freshwater environments. Appl. Environ. Microbiol. 2005, 71, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Butina, T.V.; Potapov, S.A.; Belykh, O.I.; Belikov, S.I. Genetic diversity of cyanophages of the Myoviridae family as a constituent of the associated community of the Baikal sponge Lubomirskia baicalensis. Genetika 2015, 51, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Liu, J.; Yu, Z.; Liu, X.; Wang, G. Phylogenetic distribution of the capsid assembly protein gene (g20) of cyanophages in paddy floodwaters in Northeast China. PLoS ONE 2014, 9, e88634. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.C.; Goddard, V.J.; Davy, J.; Schroeder, D.C.; Adams, D.G.; Wilson, W.H. Identification of a diagnostic marker to detect freshwater cyanophages of filamentous cyanobacteria. Appl. Environ. Microbiol. 2006, 72, 5713–5719. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Yoshida, T.; Yoshida, M.; Shirai, Y.; Tomaru, Y.; Takao, Y.; Hiroishi, S.; Nagasaki, K. Development and application of quantitative detection of cyanophages phylogenetically related to cyanophage Ma-LMM01 infecting Microcystis aeruginosa in fresh water. Microbes Environ. 2007, 22, 207–213. [Google Scholar] [CrossRef]
- Chen, F.; Wang, K.; Huang, S.; Cai, H.; Zhao, M.; Jiao, N.; Wommack, K.E. Diverse and dynamic populations of cyanobacterial podoviruses in the Chesapeake Bay unveiled through DNA polymerase gene sequences. Environ. Microbiol. 2009, 11, 2884–2892. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wilhelm, S.W.; Jiao, N.; Chen, F. Ubiquitous cyanobacterial podoviruses in the global oceans unveiled through viral DNA polymerase gene sequences. ISME J 2010, 4, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, E.M.; Cowan, D.A. Using signature genes as tools to assess environmental viral ecology and diversity. Appl. Environ. Microbiol. 2014, 80, 4470–4480. [Google Scholar] [CrossRef] [PubMed]
- Zeidner, G.; Preston, C.M.; Delong, E.F.; Massana, R.; Post, A.F.; Scanlan, D.J.; Beja, O. Molecular diversity among marine picophytoplankton as revealed by psbA analyses. Environ. Microbiol. 2003, 5, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, F. Prevalence of highly host-specific cyanophages in the estuarine environment. Environ. Microbiol. 2008, 10, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Chénard, C.; Suttle, C.A. Phylogenetic diversity of sequences of cyanophage photosynthetic gene psbA in marine and freshwaters. Appl. Environ. Microbiol. 2008, 74, 5317–5324. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Murase, J.; Asakawa, S.; Kimura, M. Novel cyanophage photosynthetic gene psbA in the floodwater of a Japanese rice field. FEMS Microbiol. Ecol. 2009, 70, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Jacquet, S. Prevalence of viral photosynthetic and capsid protein genes from cyanophages in two large and deep perialpine lakes. Appl. Environ. Microbiol. 2013, 79, 7169–7178. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Marianovsky, I.; Glaser, G. MazG—A regulator of programmed cell death in Escherichia coli. Mol. Microbiol. 2006, 59, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Ramisetty, B.C.; Raj, S.; Ghosh, D. Escherichia coli MazEF toxin-antitoxin system does not mediate programmed cell death. J. Basic Microbiol. 2016, 56, 1–5. [Google Scholar]
- Bryan, M.J.; Burroughs, N.J.; Spence, E.M.; Clokie, M.R.J.; Mann, N.H.; Bryan, S.J. Evidence for the intense exchange of mazG in marine cyanophages by horizontal gene transfer. PLoS ONE 2008, 3, e2048. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, D.B.; Crosti, G.; Dwivedi, B.; McDaniel, L.D.; Varsani, A.; Suttle, C.A.; Weinbauer, M.G.; Sandaa, R.A.; Breitbart, M. Development of phoH as a novel signature gene for assessing marine phage diversity. Appl. Environ. Microbiol. 2011, 77, 7730–7739. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, D.B.; Parsons, R.J.; Beyene, D.; Salamon, P.; Breitbart, M. Deep sequencing of the viral phoH gene reveals temporal variation, depth-specific composition, and persistent dominance of the same viral phoH genes in the Sargasso Sea. Peer J. 2015, 3, e997. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Allen, L.Z.; Palenik, B. Diversity and genome dynamics of marine cyanophages using metagenomic analyses. Environ. Microbiol. Rep. 2014, 6, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Hevroni, G.; Enav, H.; Rohwer, F.; Béjà, O. Diversity of viral photosystem-I psaA genes. ISME J. 2015, 9, 1892–1898. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Signature Gene | Function | Primer Sequence | References |
|---|---|---|---|
| psbA | Photosynthesis protein D1 | psbA-F: 5′-GTNGAYATHGAYGGNATHMGNGARCC-3′ psbA-R: 5′-GGRAARTTRTGNGCRTTNCKYTCRTGCAT-3′ | [100] |
| Pro-psbA-F: 5′-AACATCATYTCWGGTGCWGT-3′ Pro-psbA-R: 5′-TCGTGCATTACTTCCATACC-3′ | [58] | ||
| psbA-93F: 5′-TAYCCNATYTGGGAAGC-3′ psbA-341R: 5′-TCRAGDGGGAARTTRTG-3′ | [109] | ||
| psbD | Photosynthesis protein D2 | psbD-26Fa: 5′-TTYGTNTTYRTNGGNTGGAGYGG-3′ psbD-26Fb: 5′-TTYGTNTTYRTNGGNTGGTCNGG-3′ | [58] |
| psbD-54Fa: 5′-GTNACNAGYTGGTAYACNCAYGG-3′ psbD-54Fb: 5′-GTNACNTCNTGGTAYACNCAYGG-3′ | [58] | ||
| psbD-308Ra: 5′-YTCYTGNGANACRAARTCRTANGC-3′ psbD-308Rb: 5′-YTCYTGRCTNACRAARTCRTANGC-3′ | [58] | ||
| psbD-F: 5′-GGNTTYATGCTNMGNCARTT-3′ psbD-R: 5′-CKRTTNGCNGTVAYCAT-3′ | [27] | ||
| cobS | Putative porphyrin biosynthetic protein | cobS-For: 5′-BACYGTWTGGCACAAYGG-3′ cobS-Rev: 5′-CTTRGTNTCMTCATCRAARCG-3′ | [63] |
| mazG | Nucleoside triphosphate pyrophosphohydrolase | mazG-For: 5′-CTTCTTACTGCTGSYGTTGG-3′ mazG-Rev: 5′-TTATCKGTCRTCKRCWGATT-3′ | [104] |
| phoH | Putative phosphate protein | vPhoHf: 5′-TGCRGGWACAGGTAARACAT-3′ vPhoHr: 5′-TCRCCRCAGAAAAYMATTTT-3′ | [106] |
| phoH-For: 5′-GARATYGGDTTCYTDCCTGG-3′ phoH-Rev: 5′-ACWARWCCAGADCKWACRATRTC-3′ | [63] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, E.-B.; Huang, Y.; Ning, D. Metabolic Genes within Cyanophage Genomes: Implications for Diversity and Evolution. Genes 2016, 7, 80. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100080
Gao E-B, Huang Y, Ning D. Metabolic Genes within Cyanophage Genomes: Implications for Diversity and Evolution. Genes. 2016; 7(10):80. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100080
Chicago/Turabian StyleGao, E-Bin, Youhua Huang, and Degang Ning. 2016. "Metabolic Genes within Cyanophage Genomes: Implications for Diversity and Evolution" Genes 7, no. 10: 80. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7100080