
Screening for Mutations in the TBX1 Gene on Chromosome 22q11.2 in Schizophrenia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
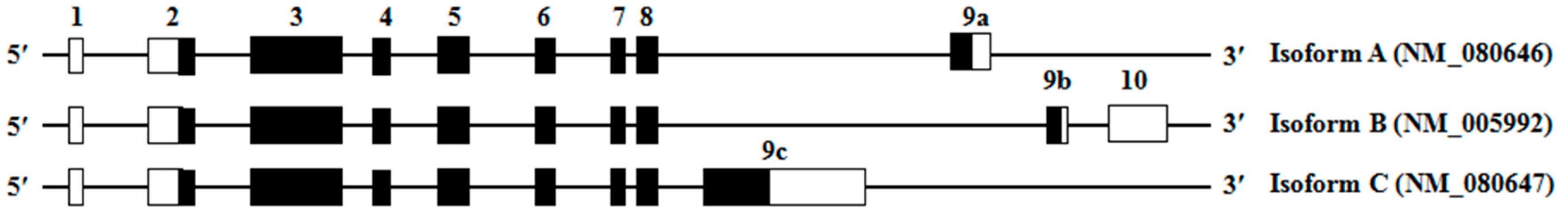
2.2. Nomenclature and Reference Sequences
2.3. PCR-Based Sequencing
2.4. In Silico Analysis
2.5. Reporter Gene Activity Assay
2.6. Statistical Analysis
3. Results
3.1. Genetic Analysis of the TBX1 Gene
3.2. Reporter Gene Activity Assay
3.3. Clinical Findings of the Patients with Rare Mutations
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Freedman, R. Schizophrenia. N. Engl. J. Med. 2003, 349, 1738–1749. [Google Scholar] [PubMed]
- Malhotra, D.; Sebat, J. Cnvs: Harbingers of a rare variant revolution in psychiatric genetics. Cell 2012, 148, 1223–1241. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, N.; Takahashi, T.; Hishimoto, A.; Izumi, T.; Boku, S.; Hiramoto, T. Copy number variation at 22q11.2: From rare variants to common mechanisms of developmental neuropsychiatric disorders. Mol. Psychiatry 2013, 18, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Szatkiewicz, J.P.; O’Dushlaine, C.; Chen, G.; Chambert, K.; Moran, J.L.; Neale, B.M.; Fromer, M.; Ruderfer, D.; Akterin, S.; Bergen, S.E.; et al. Copy number variation in schizophrenia in sweden. Mol. Psychiatry 2014, 19, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Botto, L.D.; May, K.; Fernhoff, P.M.; Correa, A.; Coleman, K.; Rasmussen, S.A.; Merritt, R.K.; O’Leary, L.A.; Wong, L.-Y.; Elixson, E.M.; et al. A population-based study of the 22q11.2 deletion: Phenotype, incidence, and contribution to major birth defects in the population. Pediatrics 2003, 112, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kobrynski, L.J.; Sullivan, K.E. Velocardiofacial syndrome, digeorge syndrome: The chromosome 22q11.2 deletion syndromes. Lancet 2007, 370, 1443–1452. [Google Scholar] [CrossRef]
- Gothelf, D.; Feinstein, C.; Thompson, T.; Gu, E.; Penniman, L.; van Stone, E.; Kwon, H.; Eliez, S.; Reiss, A.L. Risk factors for the emergence of psychotic disorders in adolescents with 22q11.2 deletion syndrome. Am. J. Psychiatry 2007, 164, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.C.; Jones, L.A.; Owen, M.J. High rates of schizophrenia in adults with velo-cardio-facial syndrome. Arch. Gen. Psychiatry 1999, 56, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Debbane, M.; Bassett, A.S.; Chow, E.W.; Fung, W.L.; van den Bree, M.; Owen, M.; Murphy, K.C.; Niarchou, M.; Kates, W.R.; et al. Psychiatric disorders from childhood to adulthood in 22q11.2 deletion syndrome: Results from the international consortium on brain and behavior in 22q11.2 deletion syndrome. Am. J. Psychiatry 2014, 171, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Monks, S.; Niarchou, M.; Davies, A.R.; Walters, J.T.; Williams, N.; Owen, M.J.; van den Bree, M.B.; Murphy, K.C. Further evidence for high rates of schizophrenia in 22q11.2 deletion syndrome. Schizophr. Res. 2014, 153, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Grozeva, D.; Conrad, D.F.; Barnes, C.P.; Hurles, M.; Owen, M.J.; O’Donovan, M.C.; Craddock, N.; Kirov, G. Independent estimation of the frequency of rare cnvs in the uk population confirms their role in schizophrenia. Schizophr. Res. 2012, 135, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Christofolini, D.M.; Bellucco, F.T.; Ota, V.K.; Belangero, S.I.; Cernach, M.C.P.; Gadelha, A.; Mari, J.; Bressan, R.A.; Smith, M.A.C.; Melaragno, M.I. Assessment of 22q11.2 copy number variations in a sample of brazilian schizophrenia patients. Schizophr. Res. 2011, 132, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Abraham, S.; Gronostajski, R.M.; Campbell, C.E. Differential DNA binding and transcription modulation by three t-box proteins, T, TBX1 and TBX2. Gene 2000, 258, 15–29. [Google Scholar] [CrossRef]
- Chieffo, C.; Garvey, N.; Gong, W.; Roe, B.; Zhang, G.; Silver, L.; Emanuel, B.S.; Budarf, M.L. Isolation and characterization of a gene from the digeorge chromosomal region homologous to the mouse TBX1 gene. Genomics 1997, 43, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Furutani, Y.; Hamada, H.; Sasaki, T.; Asakawa, S.; Minoshima, S.; Ichida, F.; Joo, K.; Kimura, M.; Imamura, S.-I.; et al. Role of TBX1 in human del22q11.2 syndrome. Lancet 2003, 362, 1366–1373. [Google Scholar] [CrossRef]
- Jerome, L.A.; Papaioannou, V.E. Digeorge syndrome phenotype in mice mutant for the T-box gene, TBX1. Nat. Genet. 2001, 27, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, E.A.; Vitelli, F.; Su, H.; Morishima, M.; Huynh, T.; Pramparo, T.; Jurecic, V.; Ogunrinu, G.; Sutherland, H.F.; Scambler, P.J.; et al. TBX1 haploinsufficiency in the digeorge syndrome region causes aortic arch defects in mice. Nature 2001, 410, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Merscher, S.; Funke, B.; Epstein, J.A.; Heyer, J.; Puech, A.; Lu, M.M.; Xavier, R.J.; Demay, M.B.; Russell, R.G.; Factor, S.; et al. TBX1 is responsible for cardiovascular defects in velo-cardio-facial/digeorge syndrome. Cell 2001, 104, 619–629. [Google Scholar] [CrossRef]
- Guna, A.; Butcher, N.J.; Bassett, A.S. Comparative mapping of the 22q11.2 deletion region and the potential of simple model organisms. J. Neurodev. Disord. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Maynard, T.M.; Haskell, G.T.; Peters, A.Z.; Sikich, L.; Lieberman, J.A.; LaMantia, A.-S. A comprehensive analysis of 22q11 gene expression in the developing and adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 14433–14438. [Google Scholar] [CrossRef] [PubMed]
- Paylor, R.; Glaser, B.; Mupo, A.; Ataliotis, P.; Spencer, C.; Sobotka, A.; Sparks, C.; Choi, C.-H.; Oghalai, J.; Curran, S.; et al. TBX1 haploinsufficiency is linked to behavioral disorders in mice and humans: Implications for 22q11 deletion syndrome. Proc. Natl. Acad. Sci. USA 2006, 103, 7729–7734. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, N.; Zhu, H.; Lee, M.; Funke, B.; Arai, M.; Itokawa, M.; Kucherlapati, R.; Morrow, B.; Sawamura, T.; Agatsuma, S. A 200-kb region of human chromosome 22q11.2 confers antipsychotic-responsive behavioral abnormalities in mice. Proc. Natl. Acad. Sci. USA 2005, 102, 19132–19137. [Google Scholar] [CrossRef] [PubMed]
- Flore, G.; Cioffi, S.; Bilio, M.; Illingworth, E. Cortical development requires mesodermal expression of TBX1, a gene haploinsufficient in 22q11.2 deletion syndrome. Cereb. Cortex 2016. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, S.; Martucciello, S.; Fulcoli, F.G.; Bilio, M.; Ferrentino, R.; Nusco, E.; Illingworth, E. TBX1 regulates brain vascularization. Hum. Mol. Genet. 2014, 23, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, T.; Kang, G.; Suzuki, G.; Satoh, Y.; Kucherlapati, R.; Watanabe, Y.; Hiroi, N. TBX1: Identification of a 22q11.2 gene as a risk factor for autism spectrum disorder in a mouse model. Hum. Mol. Genet. 2011, 20, 4775–4785. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Okabe, S.; Broin, P.O.; Nishi, A.; Ye, K.; Beckert, M.V.; Izumi, T.; Machida, A.; Kang, G.; Abe, S.; et al. Structure and function of neonatal social communication in a genetic mouse model of autism. Mol. Psychiatry 2016, 21, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, G.; Minshew, N.J.; Allen, D.N.; Seaton, B.E. High-functioning autism and schizophrenia: A comparison of an early and late onset neurodevelopmental disorder. Arch. Clin. Neuropsychol. 2002, 17, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Cichon, S.; Craddock, N.; Daly, M.; Faraone, S.V.; Gejman, P.V.; Kelsoe, J.; Lehner, T.; Levinson, D.F.; Moran, A.; Sklar, P.; et al. Genomewide association studies: History, rationale, and prospects for psychiatric disorders. Am. J. Psychiatry 2009, 166, 540–556. [Google Scholar] [PubMed]
- Craddock, N.; O’Donovan, M.C.; Owen, M.J. Phenotypic and genetic complexity of psychosis. Invited commentary on ... Schizophrenia: A common disease caused by multiple rare alleles. Br. J. Psychiatry 2007, 190, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Kenny, E.M.; Cormican, P.; Furlong, S.; Heron, E.; Kenny, G.; Fahey, C.; Kelleher, E.; Ennis, S.; Tropea, D.; Anney, R.; et al. Excess of rare novel loss-of-function variants in synaptic genes in schizophrenia and autism spectrum disorders. Mol. Psychiatry 2014, 19, 872–879. [Google Scholar] [CrossRef] [PubMed]
- McClellan, J.M.; Susser, E.; King, M.-C. Schizophrenia: A common disease caused by multiple rare alleles. Br. J. Psychiatry 2007, 190, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Den Dunnen, J.T.; Antonarakis, S.E. Mutation nomenclature. Curr. Protoc. Hum. Genet. 2003. [Google Scholar] [CrossRef]
- Funke, B.H.; Lencz, T.; Finn, C.T.; DeRosse, P.; Poznik, G.D.; Plocik, A.M.; Kane, J.; Rogus, J.; Malhotra, A.K.; Kucherlapati, R. Analysis of TBX1 variation in patients with psychotic and affective disorders. Mol. Med. 2007, 13, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Shi, Y.; Tang, W.; He, Z.; Huang, K.; Li, Z.; He, G.; Feng, G.; Li, H.; He, L. An association study between the genetic polymorphisms within TBX1 and schizophrenia in the chinese population. Neurosci. Lett. 2007, 425, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Ogata, T.; Niihori, T.; Tanaka, N.; Kawai, M.; Nagashima, T.; Funayama, R.; Nakayama, K.; Nakashima, S.; Kato, F.; Fukami, M.; et al. TBX1 mutation identified by exome sequencing in a japanese family with 22q11.2 deletion syndrome-like craniofacial features and hypocalcemia. PLoS ONE 2014, 9, e91598. [Google Scholar] [CrossRef] [PubMed]
- Girirajan, S.; Rosenfeld, J.A.; Cooper, G.M.; Antonacci, F.; Siswara, P.; Itsara, A.; Vives, L.; Walsh, T.; McCarthy, S.E.; Baker, C.; et al. A recurrent 16p12.1 microdeletion supports a two-hit model for severe developmental delay. Nat. Genet. 2010, 42, 203–209. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variant | Group | n | Genotype | HWE | p | Allele | p | |||
|---|---|---|---|---|---|---|---|---|---|---|
| c.-85G>C (rs737868) | G/G | G/C | C/C | 0.60 | G | C | 0.46 | |||
| SZ | 564 | 105(18.6%) | 269(47.7%) | 190(33.7%) | 0.57 | 479(42.5%) | 649(57.5%) | |||
| Ctrl | 546 | 89(16.3%) | 269(49.3%) | 188(34.4%) | 0.66 | 447(40.9%) | 645(59.1%) | |||
| p.Phe140= (rs41298814) | T/T | T/C | C/C | 0.40 | T | C | 0.20 | |||
| SZ | 594 | 161(27.1%) | 293(49.3%) | 140(23.6%) | 0.77 | 615(51.8%) | 573(48.2%) | |||
| Ctrl | 567 | 166(29.3%) | 285(50.3%) | 116(20.4%) | 0.75 | 617(54.4%) | 517(45.6%) | |||
| p.Leu222= (rs2301558) | C/C | C/T | T/T | 0.48 | C | T | 0.77 | |||
| SZ | 518 | 397(76.6%) | 114(22.0%) | 7(1.4%) | 0.71 | 908(87.6%) | 128(12.4%) | |||
| Ctrl | 536 | 419(78.2%) | 106(19.8%) | 11(2.0%) | 0.17 | 944(88.1%) | 128(11.9%) | |||
| p.Asn397His (rs72646967) | A/A | A/C | C/C | 0.20 | A | C | 0.07 | |||
| SZ | 529 | 151(28.5%) | 238(45.0%) | 140(26.5%) | 0.02 | 540(51.0%) | 518(49.0%) | |||
| Ctrl | 524 | 167(31.9%) | 242(46.2%) | 115(22.0%) | 0.12 | 476(55.0%) | 472(45.0%) | |||
| p.Thr350Met (rs4819522) | C/C | C/T | T/T | 0.78 | C | T | 0.64 | |||
| SZ | 653 | 502(76.9%) | 138(21.1%) | 13(0.2%) | 0.34 | 1142(87.4%) | 164(12.6%) | |||
| Ctrl | 545 | 423(77.6%) | 114(20.9%) | 8(1.5%) | 0.92 | 960(88.1%) | 130(11.9%) | |||
| c.*121G>T (rs5746826) | G/G | G/T | T/T | 0.38 | G | T | 0.16 | |||
| SZ | 650 | 110(16.9%) | 306(47.1%) | 234(36.0%) | 0.56 | 526(40.5%) | 774(59.5%) | |||
| Ctrl | 544 | 107(19.7%) | 257(47.2%) | 180(33.1%) | 0.38 | 471(43.3%) | 617(56.7%) | |||
| Variant (TBX1 Isoform) | RS Number | Frequency | Amino Acid Change | In Silico Analysis | ||||
|---|---|---|---|---|---|---|---|---|
| SZ | Ctrl | Transcription Factor a | Pmut | PolyPhen-2 | SIFT | |||
| c.-123G>C (isoform C) | None | 1/638 | 0/534 | N/A | Wild-type: AP-2, LVc | N/A | N/A | N/A |
| c.-120G>T (isoform C) | None | 0/637 | 1/534 | N/A | Wild-type: AP-2 | N/A | N/A | N/A |
| c.-84G>A (isoform C) | None | 0/564 | 1/546 | N/A | N/A | N/A | N/A | N/A |
| c.-11delC (isoform C) | rs78833362 | 1/565 | 0/546 | N/A | N/A | N/A | N/A | N/A |
| c.453T>A (isoform C) | rs778041960 | 1/594 | 0/567 | p.Asp151Glu | N/A | Neutral | Benign | Deleterious |
| c.463G>A (isoform C) | rs374011293 | 0/594 | 3/567 | p.Asp155Asn | N/A | Neutral | Possibly damaging | Deleterious |
| c.768C>T (isoform C) | rs759225333 | 0/553 | 1/548 | p.Ala256= | N/A | N/A | N/A | |
| c.770A>C (isoform C) | None | 1/553 | 0/548 | p.Glu257Ala | N/A | Neutral | Benign | Deleterious |
| c.813C>T (isoform C) | rs61730282 | 0/449 | 1/548 | p.Thr271= | N/A | N/A | N/A | |
| c.1025G>A (isoform C) | rs549715785 | 2/535 | 0/515 | p.Arg342Gln | N/A | Pathological | Benign | Deleterious |
| c.1059A>G (isoform C) | rs13054377 | 1/426 | 0/528 | pAla353= | N/A | N/A | N/A | N/A |
| c.1194C>A (isoform C) | None | 1/531 | 0/400 | p.Pro398= | N/A | N/A | N/A | N/A |
| c.*13C>T (isoform C) | rs543378212 | 0/489 | 2/525 | N/A | N/A | N/A | N/A | N/A |
| c.*108G>C (isoform C) | None | 0/500 | 1/525 | N/A | N/A | N/A | N/A | N/A |
| c.*164_*165insG (isoform C) | rs41298842 | 10/500 | 10/534 | N/A | N/A | N/A | N/A | N/A |
| c.*315C>T (isoform C) | None | 1/488 | 0/535 | N/A | N/A | N/A | N/A | N/A |
| c.*399G>A (isoform C) | None | 0/487 | 3/534 | N/A | N/A | N/A | N/A | N/A |
| c.*398-406 delTGTAGATAC (isoform C) | None | 1/488 | 1/534 | N/A | N/A | N/A | N/A | N/A |
| c.1076T>C (isoform A) | rs200361367 | 0/543 | 1/544 | p.Val359Ala | N/A | Pathological | Possibly damaging | Tolerated |
| c.1177G>A (isoform A) | None | 1/652 | 1/544 | p.Ala393Thr | N/A | Neutral | Benign | Deleterious |
| c.1185C>T (isoform A) | None | 3/652 | 1/544 | p.Asp395= | N/A | N/A | N/A | N/A |
| c.1187G>A (isoform A) | rs207477905 | 1/542 | 0/544 | p.Arg396His | N/A | Neutral | Benign | Deleterious |
| c.*24C>A (isoform A) | rs41298008 | 12/542 | 9/544 | N/A | N/A | N/A | N/A | N/A |
| c.*123G>A (isoform A) | None | 1/650 | 0/544 | N/A | N/A | N/A | N/A | N/A |
| c.*170C>T (isoform B) | None | 1/588 | 0/326 | N/A | N/A | N/A | N/A | N/A |
| Mutation | Fluc/Rluc (n = 6) | p Value |
|---|---|---|
| Wild-type | 36.96 ± 3.92 | |
| c.-123C | 18.19 ± 2.39 | <0.01 * |
| c.-120T | 36.17 ± 3.64 | =0.72 |
| pGL3-Enhancer | 12.83 ± 2.00 | |
| Wild-type | 18.02 ± 1.70 | |
| c.-84A | 43.90 ± 4.21 | <0.01 * |
| c.-11delC | 52.39 ± 8.76 | <0.01 * |
| pGL3-Enhancer | 18.50 ± 3.73 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ping, L.-Y.; Chuang, Y.-A.; Hsu, S.-H.; Tsai, H.-Y.; Cheng, M.-C. Screening for Mutations in the TBX1 Gene on Chromosome 22q11.2 in Schizophrenia. Genes 2016, 7, 102. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110102
Ping L-Y, Chuang Y-A, Hsu S-H, Tsai H-Y, Cheng M-C. Screening for Mutations in the TBX1 Gene on Chromosome 22q11.2 in Schizophrenia. Genes. 2016; 7(11):102. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110102
Chicago/Turabian StylePing, Lieh-Yung, Yang-An Chuang, Shih-Hsin Hsu, Hsin-Yao Tsai, and Min-Chih Cheng. 2016. "Screening for Mutations in the TBX1 Gene on Chromosome 22q11.2 in Schizophrenia" Genes 7, no. 11: 102. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110102