Bioinformatics Analysis of MAPKKK Family Genes in Medicago truncatula


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the MAPKKK Gene Family in Medicago truncatula
2.2. Phylogenetic Analysis of the MtMAPKKK Genes in M. truncatula
2.3. Chromosomal Location and Gene Duplication of MtMAPKKK Genes
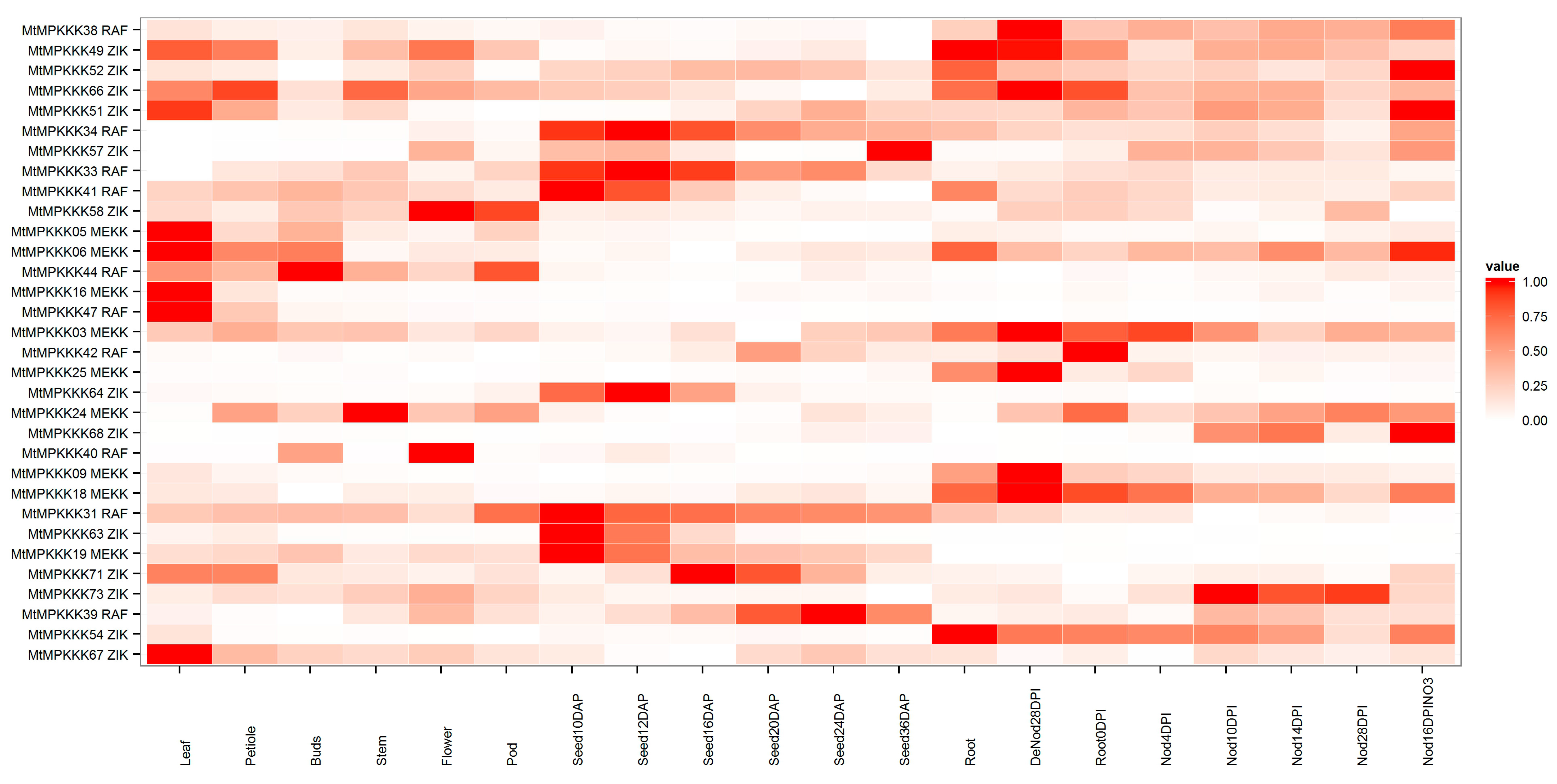
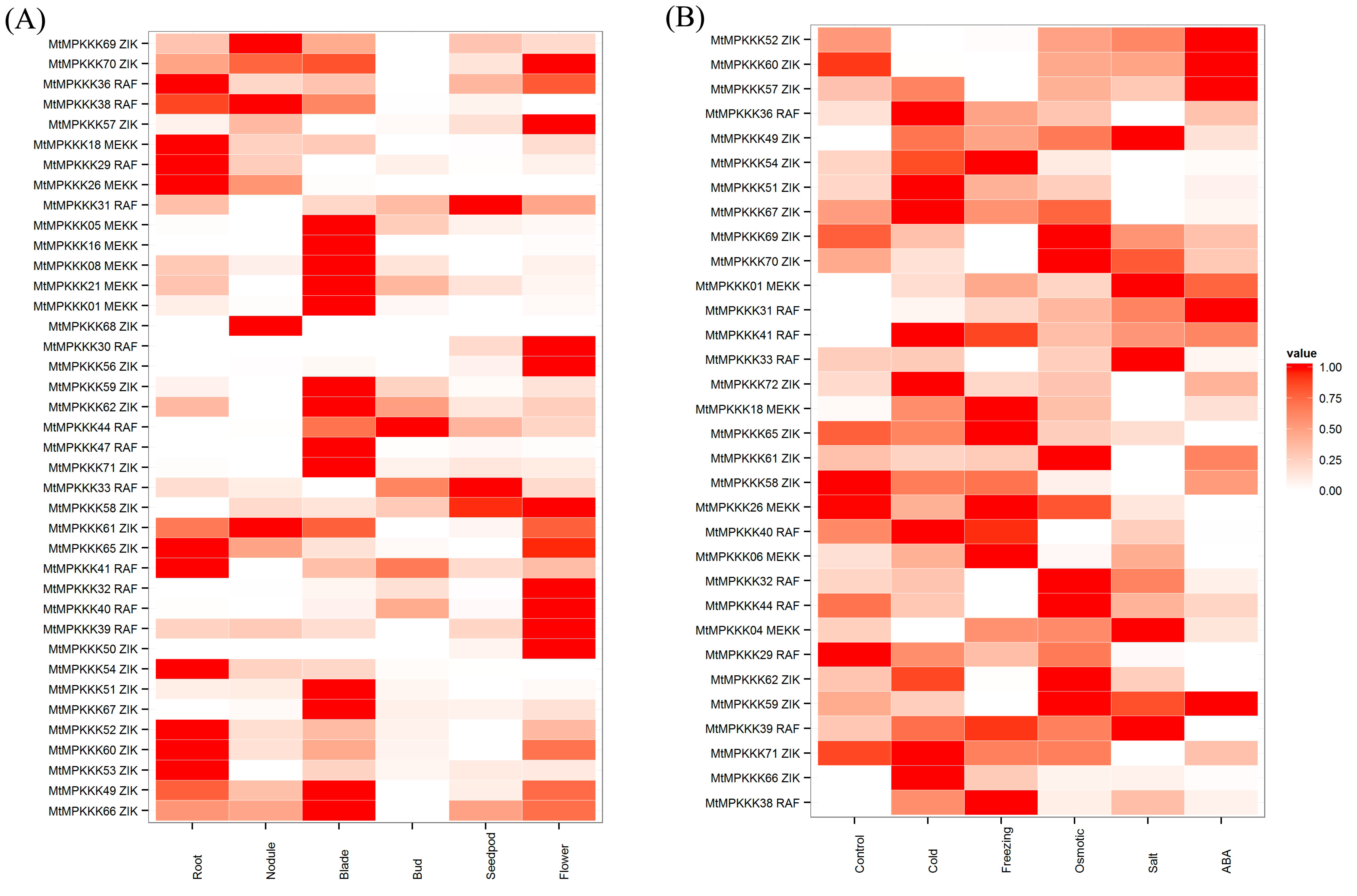
2.4. Expression Analysis of MtMAPKKK Genes in Growth and Development
2.5. Expression Analysis of MtMAPKKK Genes’ Response to Abiotic Stress
3. Results
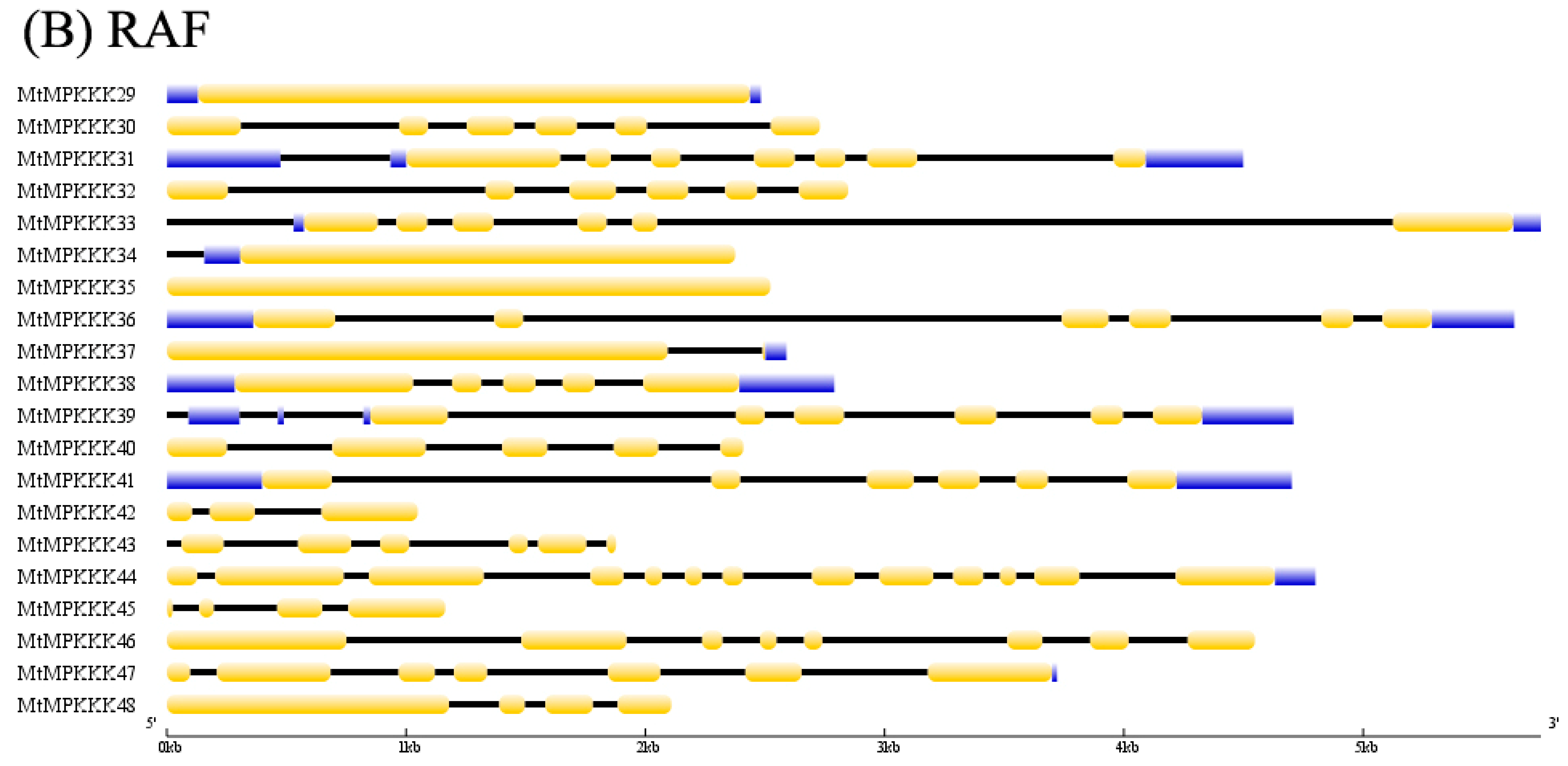
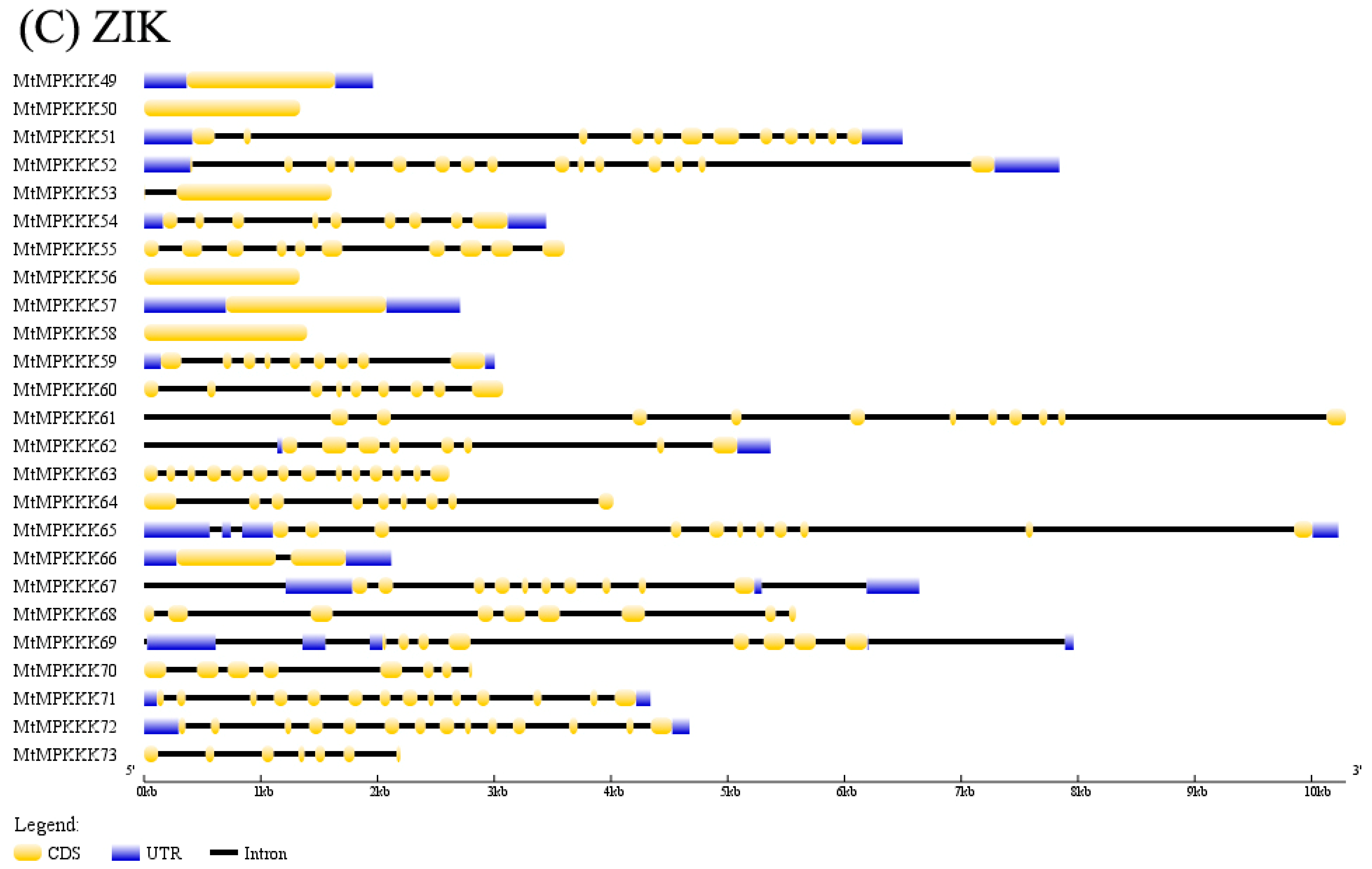
3.1. Identification and Characterization of MAPKKK Family in M. truncatula
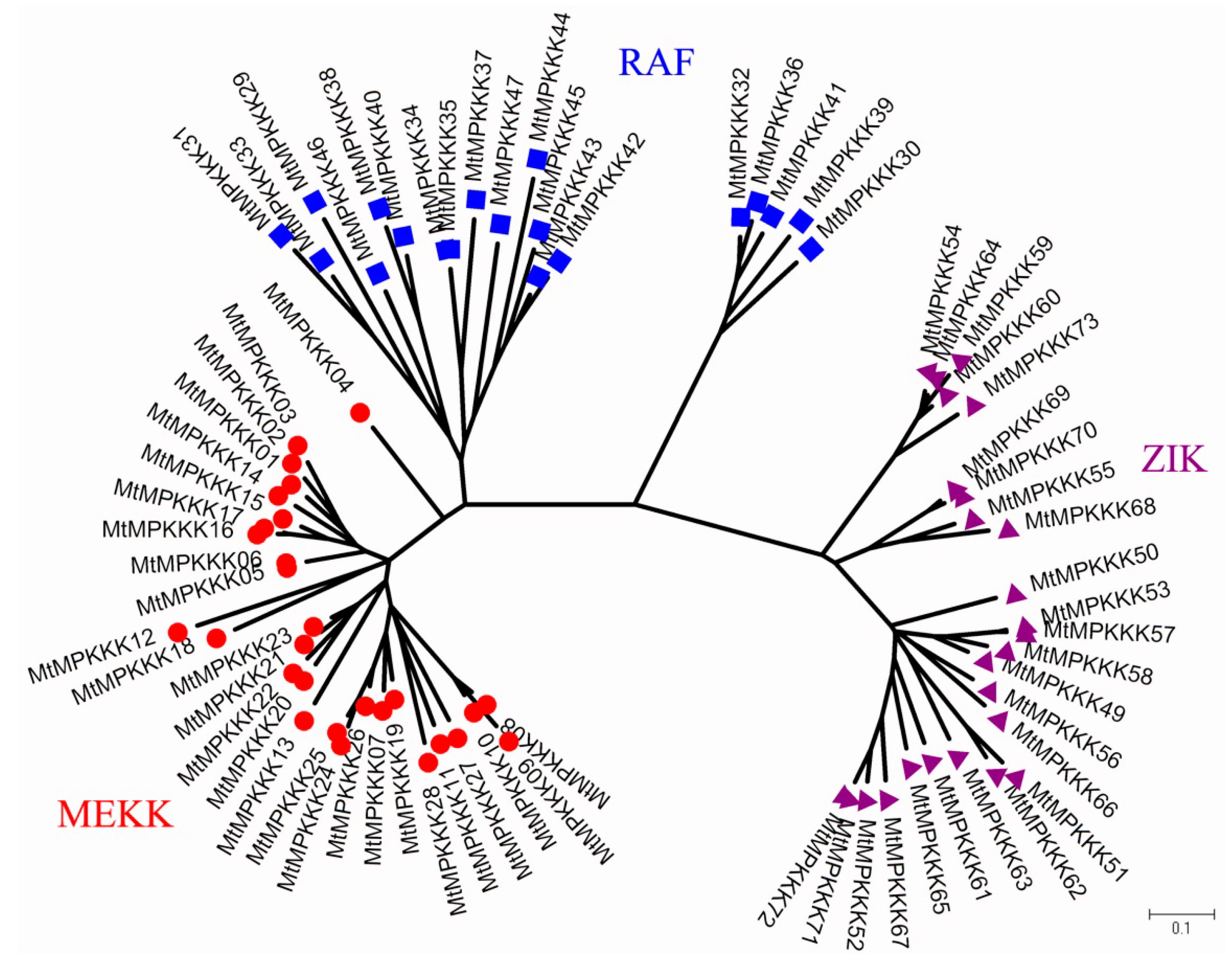
3.2. Phylogenetic Analysis of MtMAPKKK Genes
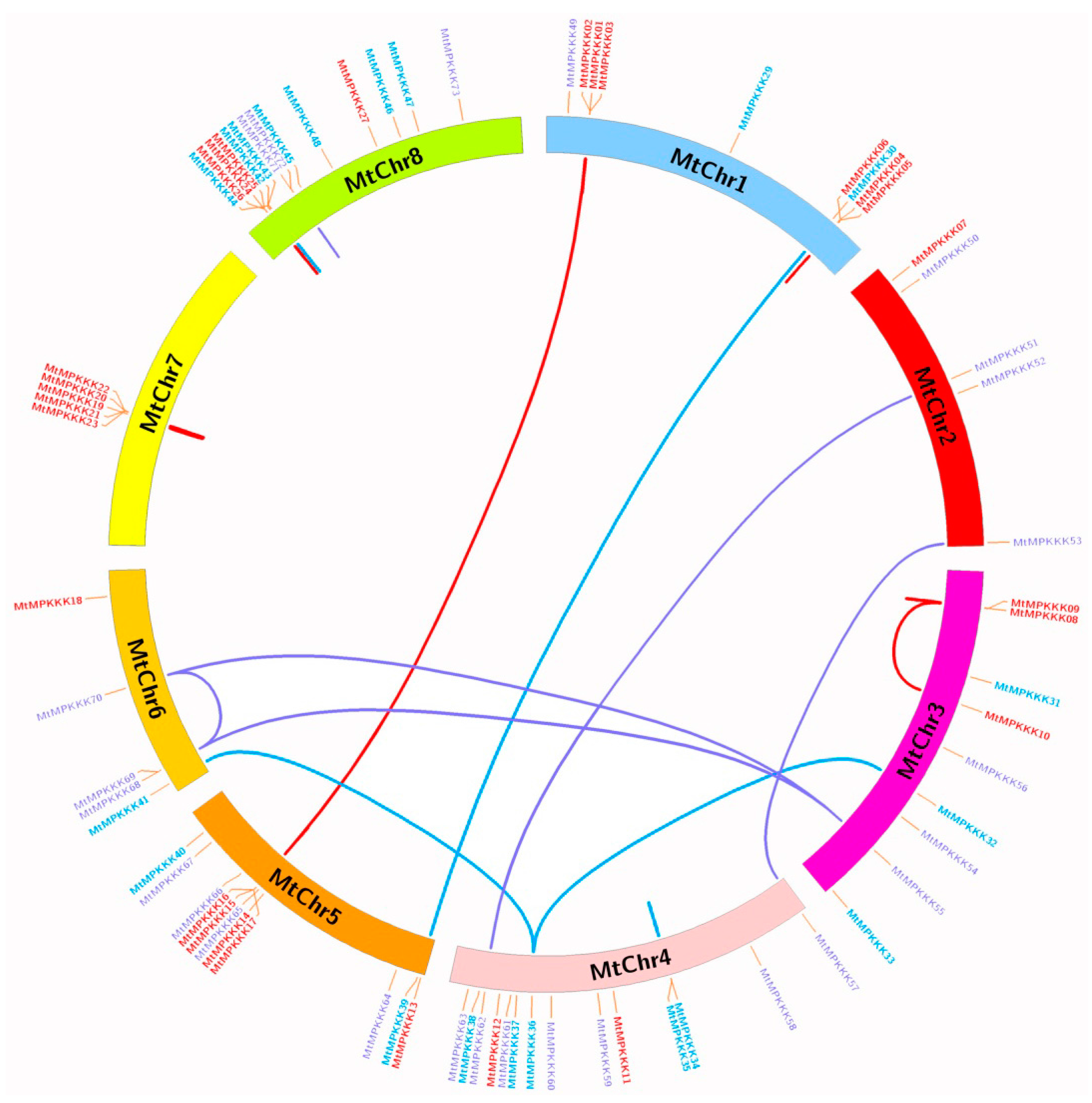
3.3. Chromosomal Location and Duplication Analysis of MtMAPKKK Genes
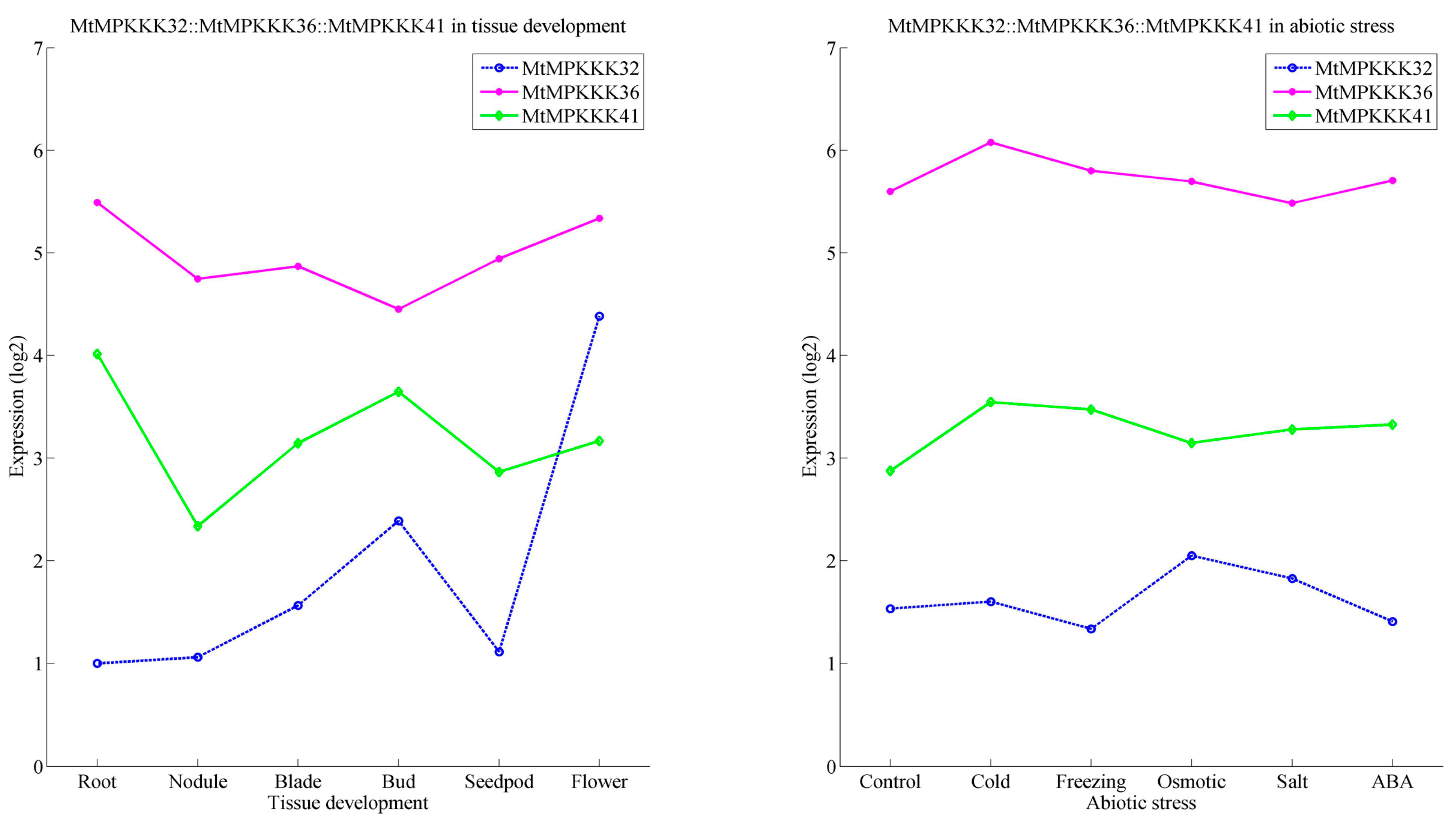
3.4. In Silico Expression Analysis of MtMAPKKK Genes Involved in Growth and Development
3.5. Expression Analysis of MtMAPKKK Genes in Response to Abiotic Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MAPKKK | Mitogen-activated protein kinase kinase kinase |
| MtGEA | Medicago truncatula Gene Expression Atlas |
| NCBI | National Center for Biotechnology Information |
| ROS | reactive oxygen species |
| SRA | short read archive |
| MtChr | M. truncatula chromosomes |
References
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Doczi, R.; Okresz, L.; Romero, A.E.; Paccanaro, A.; Bogre, L. Exploring the evolutionary path of plant MAPK networks. Trends Plant Sci. 2012, 17, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.; Hirt, H.; Wilson, C.; et al. Mitogen-activated protein kinase cascades in plants a new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar]
- Asai, T. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Caunt, C.J.; Keyse, S.M. Dual-specificity MAP kinase phosphatases (MKPs). FEBS J. 2013, 280, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Arora, P.K.; Mohanta, N.; Parida, P.; Bae, H. Identification of new members of the MAPK gene family in plants shows diverse conserved domains and novel activation loop variants. BMC Genomics 2015. [Google Scholar] [CrossRef] [PubMed]
- Krysan, P.J. An Arabidopsis mitogen-activated protein kinase kinase kinase gene family encodes essential positive regulators of cytokinesis. Plant Cell 2002, 14, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Musielak, T.J.; Bayer, M. YODA signalling in the early Arabidopsis embryo. Biochem. Soc. Trans. 2014, 42, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Pitzschke, A.; Schikora, A.; Hirt, H. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol. 2009, 12, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Qiu, L.; Wen, C.K. Possible modulation of Arabidopsis ETR1 N-terminal signaling by CTR1. Plant Signal. Behav. 2012, 7, 1243–1245. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Christiansen, K.M.; Innes, R.W. Regulation of plant disease resistance, stress responses, cell death, and ethylene signaling in Arabidopsis by the EDR1 protein kinase. Plant Physiol. 2005, 138, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Tena, G.; Asai, T.; Chiu, W.L.; Sheen, J. Plant mitogen-activated protein kinase signaling cascades. Curr. Opin. Plant Biol. 2001, 4, 392–400. [Google Scholar] [CrossRef]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A Raf-like MAPKKK gene DSM1 mediates drought resistance through reactive oxygen species scavenging in rice. Plant Physiol. 2010, 152, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Melech-Bonfil, S.; Sessa, G. Tomato MAPKKKε is a positive regulator of cell-death signaling networks associated with plant immunity. Plant J. 2010, 64, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Qu, N.; Gao, M.; Zhang, Z.; Ding, X.; Yang, F.; Li, Y.; Dong, O.X.; Chen, S.; Li, X.; et al. The MEKK1-MKK1/MKK2-MPK4 kinase cascade negatively regulates immunity mediated by a mitogen-activated protein kinase kinase kinase in Arabidopsis. Plant Cell 2012, 24, 2225–2236. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.P.; Richa, T.; Kumar, K.; Raghuram, B.; Sinha, A.K. In silico analysis reveals 75 members of mitogen-activated protein kinase kinase kinase gene family in rice. DNA Res. 2010, 17, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Lv, W.; Zhang, D.; Jiang, S.; Zhang, S.; Li, D. Genome-wide identification and analysis of expression profiles of maize mitogen-activated protein kinase kinase kinase. PLoS ONE 2013, 8, e57714. [Google Scholar] [CrossRef] [PubMed]
- Neupane, A.; Nepal, M.P.; Piya, S.; Subramanian, S.; Rohila, J.S.; Reese, R.N.; Benson, B.V. Identification, nomenclature, and evolutionary relationships of mitogen-activated protein kinase (MAPK) genes in soybean. Evol. Bioinform. 2013, 9, 363–386. [Google Scholar]
- Young, N.D.; Debelle, F.; Oldroyd, G.E.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.; Gouzy, J.; Schoof, H.; et al. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Neupane, A.; Nepal, M.P.; Piya, S.; Benson, B.V.; MacArthur, K.J. Evolutionary history of mitogen-activated protein kinase (MAPK) genes in Lotus, Medicago, and Phaseolus. Plant Signal. Behav. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pan, C.; Wang, Y.; Ye, L.; Wu, J.; Chen, L.; Zou, T.; Lu, G. Genome-wide identification of MAPK, MAPKK, and MAPKKK gene families and transcriptional profiling analysis during development and stress response in cucumber. BMC Genomics 2015. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Hu, W.; Tan, S.L.; Wang, M.; Ma, Z.B.; Zhou, S.Y. Genome-wide identification and analysis of MAPK and MAPKK gene families in Brachypodium distachyon. PLoS ONE 2012, 7, e46744. [Google Scholar] [CrossRef] [PubMed]
- Janitza, P.; Ullrich, K.K.; Quint, M. Toward a comprehensive phylogenetic reconstruction of the evolutionary history of mitogen-activated protein kinases in the plant kingdom. Front. Plant Sci. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinformatics 2003. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2013, 41, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Zhang, J.; Song, L.; Guo, C. Genome-wide analysis of the AP2/ERF superfamily genes and their responses to abiotic stress in Medicago truncatula. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Benedito, V.A.; Torres-Jerez, I.; Murray, J.D.; Andriankaja, A.; Allen, S.; Kakar, K.; Wandrey, M.; Verdier, J.; Zuber, H.; Ott, T.; et al. A gene expression atlas of the model legume Medicago truncatula. Plant J. 2008, 55, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Traprell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Callis, J.; Fromm, M.; Walbot, V. Introns increase gene expression in cultured maize cells. Genes Dev. 1987, 1, 1183–1200. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lovato, A.; Polverari, A.; Wang, M.; Liang, Y.H.; Ma, Y.C.; Cheng, Z.M. Genome-wide identification and analysis of mitogen activated protein kinase kinase kinase gene family in grapevine (Vitis vinifera). BMC Plant Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, J.; Pan, C.; Guan, X.; Wang, Y.; Liu, S.; He, Y.; Chen, J.; Chen, L.; Lu, G. Genome-wide identification of MAPKK and MAPKKK gene families in tomato and transcriptional profiling analysis during development and stress response. PLoS ONE 2014, 9, e103032. [Google Scholar] [CrossRef] [PubMed]
- Hamel, L.P.; Nicole, M.C.; Sritubtim, S.; Morency, M.J.; Ellis, M.; Ehlting, J.; Beaudoin, N.; Barbazuk, B.; Klessig, D.; Lee, J.; et al. Ancient signals: comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006, 11, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Menges, M.; Dóczi, R.; Ökrész, L.; Morandini, P.; Mizzi, L.; Soloviev, M.; Bögre, L. Comprehensive gene expression atlas for the Arabidopsis MAP kinase signalling pathways. New Phytol. 2008, 179, 643–662. [Google Scholar] [CrossRef] [PubMed]
- O'Brien, J.A.; Benkova, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Opdenakker, K.; Remans, T.; Vangronsveld, J.; Cuypers, A. Mitogen-activated protein (MAP) kinases in plant metal stress: Regulation and responses in comparison to other biotic and abiotic stresses. Mol. Sci. 2012, 13, 7828–7853. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Locus | Chromosome Location | AA | Introns | Family Group |
|---|---|---|---|---|---|
| MtMPKKK01 | Medtr1g021610 | chr1:6496817-6500636 | 671 | 6 | MEKK |
| MtMPKKK02 | Medtr1g021630 | chr1:6502386-6507333 | 677 | 6 | MEKK |
| MtMPKKK03 | Medtr1g021635 | chr1:6514070-6517643 | 675 | 6 | MEKK |
| MtMPKKK04 | Medtr1g105615 | chr1:47604242-47607699 | 507 | 4 | MEKK |
| MtMPKKK05 | Medtr1g105800 | chr1:47742832-47746207 | 654 | 6 | MEKK |
| MtMPKKK06 | Medtr1g105820 | chr1:47757298-47760553 | 672 | 6 | MEKK |
| MtMPKKK07 | Medtr2g011280 | chr2:2723023-2726806 | 842 | 7 | MEKK |
| MtMPKKK08 | Medtr3g019420 | chr3:5452378-5457705 | 822 | 6 | MEKK |
| MtMPKKK09 | Medtr3g019530 | chr3:5500322-5504353 | 779 | 5 | MEKK |
| MtMPKKK10 | Medtr3g051420 | chr3:20341370-20344887 | 781 | 5 | MEKK |
| MtMPKKK11 | Medtr4g081730 | chr4:31738243-31741501 | 667 | 6 | MEKK |
| MtMPKKK12 | Medtr4g117800 | chr4:48882742-48885495 | 492 | 7 | MEKK |
| MtMPKKK13 | Medtr5g005520 | chr5:482150-485215 | 671 | 6 | MEKK |
| MtMPKKK14 | Medtr5g065130 | chr5:27367690-27371987 | 666 | 6 | MEKK |
| MtMPKKK15 | Medtr5g068250 | chr5:28854252-28857008 | 554 | 7 | MEKK |
| MtMPKKK16 | Medtr5g068260 | chr5:28859095-28862135 | 667 | 7 | MEKK |
| MtMPKKK17 | Medtr5g068290 | chr5:28874322-28875866 | 359 | 4 | MEKK |
| MtMPKKK18 | Medtr6g083980 | chr6:31351315-31355795 | 276 | 5 | MEKK |
| MtMPKKK19 | Medtr7g056420 | chr7:19934346-19939785 | 1107 | 11 | MEKK |
| MtMPKKK20 | Medtr7g056617 | chr7:20229172-20231645 | 668 | 4 | MEKK |
| MtMPKKK21 | Medtr7g056647 | chr7:20269305-20274777 | 688 | 8 | MEKK |
| MtMPKKK22 | Medtr7g056657 | chr7:20284636-20285367 | 157 | 2 | MEKK |
| MtMPKKK23 | Medtr7g056680 | chr7:20300556-20304825 | 660 | 8 | MEKK |
| MtMPKKK24 | Medtr8g013560 | chr8:4119594-4124466 | 829 | 6 | MEKK |
| MtMPKKK25 | Medtr8g013580 | chr8:4129624-4135154 | 830 | 7 | MEKK |
| MtMPKKK26 | Medtr8g013620 | chr8:4158808-4164403 | 785 | 5 | MEKK |
| MtMPKKK27 | Medtr8g465580 | chr8:23390859-23393309 | 653 | 4 | MEKK |
| MtMPKKK28 | Medtr0090s0020 | scaffold0090:7455-10869 | 727 | 6 | MEKK |
| MtMPKKK29 | Medtr1g064560 | chr1:28404485-28406967 | 769 | 0 | RAF |
| MtMPKKK30 | Medtr1g103270 | chr1:46733948-46736677 | 383 | 5 | RAF |
| MtMPKKK31 | Medtr3g047890 | chr3:15980547-15985045 | 506 | 7 | RAF |
| MtMPKKK32 | Medtr3g078110 | chr3:35189304-35192151 | 364 | 5 | RAF |
| MtMPKKK33 | Medtr3g116590 | chr3:54529585-54535327 | 447 | 5 | RAF |
| MtMPKKK34 | Medtr4g061833 | chr4:22900571-22904005 | 690 | 0 | RAF |
| MtMPKKK35 | Medtr4g061930 | chr4:22952545-22955067 | 841 | 0 | RAF |
| MtMPKKK36 | Medtr4g106980 | chr4:43970220-43975851 | 393 | 5 | RAF |
| MtMPKKK37 | Medtr4g111925 | chr4:46269757-46272346 | 702 | 1 | RAF |
| MtMPKKK38 | Medtr4g125260 | chr4:51937267-51940056 | 515 | 4 | RAF |
| MtMPKKK39 | Medtr5g006560 | chr5:981991-986717 | 391 | 7 | RAF |
| MtMPKKK40 | Medtr5g092120 | chr5:40224346-40226755 | 373 | 4 | RAF |
| MtMPKKK41 | Medtr6g007603 | chr6:1663170-1667871 | 376 | 5 | RAF |
| MtMPKKK42 | Medtr8g014740 | chr8:4708849-4709897 | 233 | 2 | RAF |
| MtMPKKK43 | Medtr8g014860 | chr8:4751582-4753459 | 283 | 5 | RAF |
| MtMPKKK44 | Medtr8g015340 | chr8:4995169-4999969 | 908 | 12 | RAF |
| MtMPKKK45 | Medtr8g028115 | chr8:10454560-10455723 | 228 | 3 | RAF |
| MtMPKKK46 | Medtr8g064690 | chr8:27154367-27158914 | 673 | 7 | RAF |
| MtMPKKK47 | Medtr8g070910 | chr8:30050035-30053755 | 614 | 6 | RAF |
| MtMPKKK48 | Medtr8g442290 | chr8:15952474-15954582 | 571 | 3 | RAF |
| MtMPKKK49 | Medtr1g013700 | chr1:3292908-3294869 | 424 | 0 | ZIK |
| MtMPKKK50 | Medtr2g016340 | chr2:4982022-4983359 | 446 | 0 | ZIK |
| MtMPKKK51 | Medtr2g045470 | chr2:19958506-19965003 | 466 | 11 | ZIK |
| MtMPKKK52 | Medtr2g049790 | chr2:22365714-22373554 | 461 | 14 | ZIK |
| MtMPKKK53 | Medtr2g105010 | chr2:45267272-45268878 | 445 | 1 | ZIK |
| MtMPKKK54 | Medtr3g086940 | chr3:39410214-39413658 | 348 | 8 | ZIK |
| MtMPKKK55 | Medtr3g099920 | chr3:45863019-45866621 | 493 | 9 | ZIK |
| MtMPKKK56 | Medtr3g466480 | chr3:27311509-27312840 | 444 | 0 | ZIK |
| MtMPKKK57 | Medtr4g006970 | chr4:879328-882035 | 458 | 0 | ZIK |
| MtMPKKK58 | Medtr4g029020 | chr4:9994106-9995503 | 466 | 0 | ZIK |
| MtMPKKK59 | Medtr4g086855 | chr4:34076831-34079835 | 364 | 8 | ZIK |
| MtMPKKK60 | Medtr4g099240 | chr4:41124392-41127469 | 338 | 8 | ZIK |
| MtMPKKK61 | Medtr4g114670 | chr4:47183905-47194199 | 382 | 10 | ZIK |
| MtMPKKK62 | Medtr4g123940 | chr4:51095933-51101333 | 352 | 7 | ZIK |
| MtMPKKK63 | Medtr4g128820 | chr4:53613425-53616040 | 453 | 13 | ZIK |
| MtMPKKK64 | Medtr5g013550 | chr5:4318757-4322778 | 339 | 8 | ZIK |
| MtMPKKK65 | Medtr5g067150 | chr5:28353988-28364218 | 374 | 12 | ZIK |
| MtMPKKK66 | Medtr5g075100 | chr5:31902979-31905097 | 441 | 1 | ZIK |
| MtMPKKK67 | Medtr5g088350 | chr5:38344231-38350871 | 339 | 10 | ZIK |
| MtMPKKK68 | Medtr6g012980 | chr6:4042934-4048518 | 432 | 8 | ZIK |
| MtMPKKK69 | Medtr6g012990 | chr6:4049178-4057281 | 362 | 10 | ZIK |
| MtMPKKK70 | Medtr6g048250 | chr6:17403678-17408073 | 360 | 7 | ZIK |
| MtMPKKK71 | Medtr8g024590 | chr8:9075528-9079866 | 436 | 13 | ZIK |
| MtMPKKK72 | Medtr8g024600 | chr8:9083996-9088668 | 437 | 13 | ZIK |
| MtMPKKK73 | Medtr8g088740 | chr8:36861632-36863829 | 188 | 6 | ZIK |
| Species | MAPKKK | Total number of MAPKKKs | ||
|---|---|---|---|---|
| MEKK | RAF | ZIK | ||
| Arabidopsis | 21 | 48 | 11 | 80 |
| tomato | 33 | 40 | 16 | 89 |
| rice | 22 | 43 | 10 | 75 |
| maize | 22 | 46 | 6 | 74 |
| soybean | 34 | 92 | 24 | 150 |
| M. truncatula | 28 | 20 | 25 | 73 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Xu, H.; Liu, Y.; Song, L.; Guo, C.; Shu, Y. Bioinformatics Analysis of MAPKKK Family Genes in Medicago truncatula. Genes 2016, 7, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040013
Li W, Xu H, Liu Y, Song L, Guo C, Shu Y. Bioinformatics Analysis of MAPKKK Family Genes in Medicago truncatula. Genes. 2016; 7(4):13. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040013
Chicago/Turabian StyleLi, Wei, Hanyun Xu, Ying Liu, Lili Song, Changhong Guo, and Yongjun Shu. 2016. "Bioinformatics Analysis of MAPKKK Family Genes in Medicago truncatula" Genes 7, no. 4: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040013