
Stationary-Phase Mutagenesis in Stressed Bacillus subtilis Cells Operates by Mfd-Dependent Mutagenic Pathways

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of Mutant Strains
2.3. Stationary-Phase Mutagenesis Assays
2.4. Analysis of Mutation Rates
2.5. Beta-Galactosidase Assays
2.6. Quantitative Real-Time PCR
3. Results
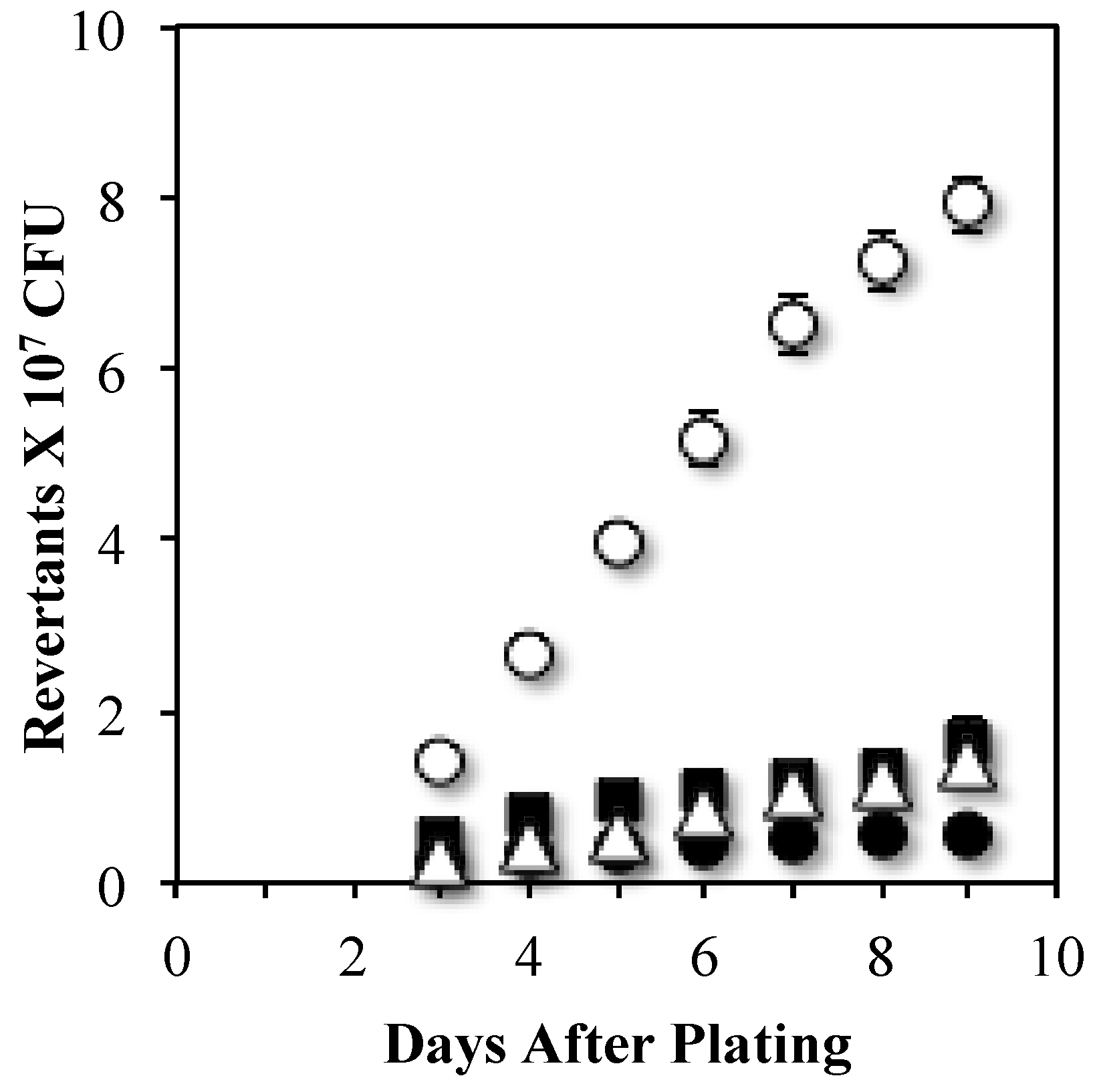
3.1. Transcription-Coupled Nucleotide Excision Repair Produces Stationary-Phase Mutations at leuC427 gene
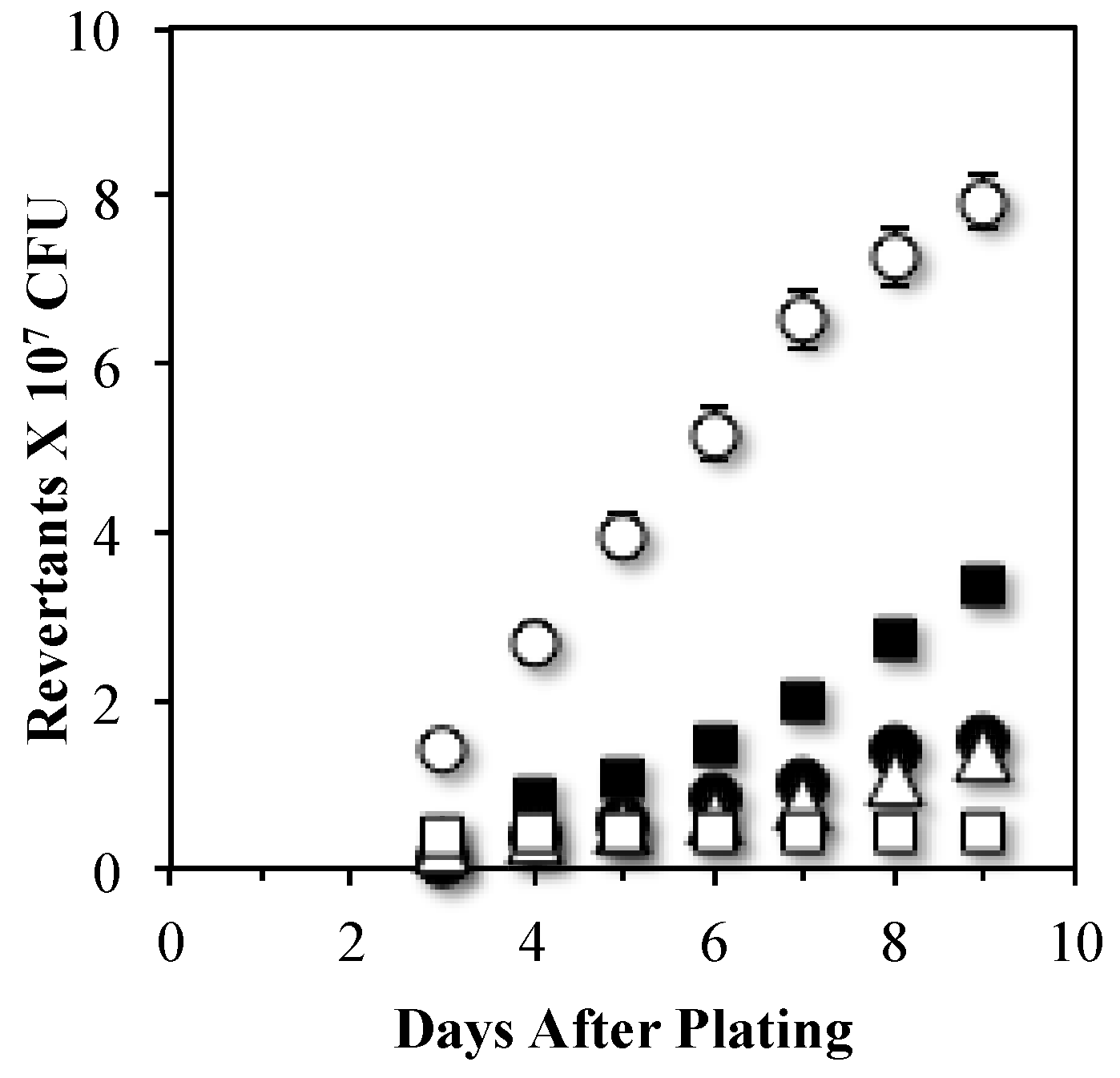
3.2. An Mfd Mutagenic Pathway Operates through MutY Activity Producing Leu+ SPM
3.3. MutYand UvrA Act in Independent Stationary-Phase Mutagenic Pathways that Require Mfd
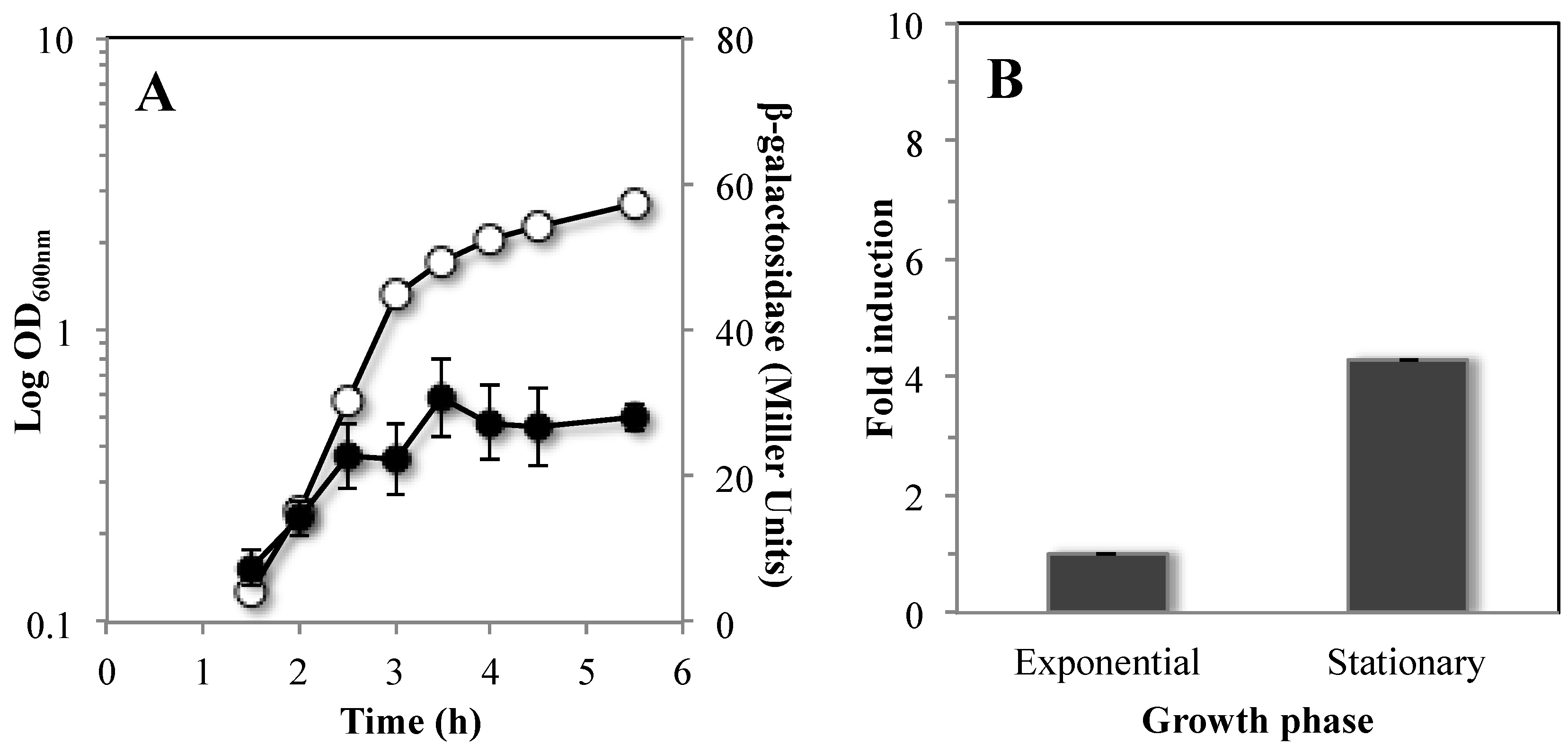
3.4. Mfd is Highly Expressed during Stationary Phase of B. subtilis
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| A, G, C | adenine, guanine and cytosine | 3-AT | 3-amino-1,2,4-triazole |
| Amp | ampicillin | AP | apurinic/apyrimidinic |
| BER | base excision repair | CFU | colony-forming unit |
| Cm | chloramphenicol | Ct | threshold cycle |
| DNA | deoxyribonucleic acid | DO | optical density |
| Em | erythromycin | GO | oxidized guanine |
| HCl | hydrochloric acid | IPTG | isopropyl-β-Dthiogalactopyranoside |
| LB | Luria-Bertani | Leu+ | leucine reversions |
| MMR | Mismatch repair | NER | nucleotide excision repair |
| NSM | non-screening medium | ONPG | ortho-notrophenyl-β-D-galactopyranoside |
| ORF | open reading frame | PAB | Penassay broth |
| PCR | Polymerase chain reaction | RNA | Ribonucleic acid |
| RNAP | ribonucleic acid polymerase | SEM | standard error of the mean |
| SM | screening medium | SMM | spizizen minimal medium |
| SMS | Spizizen minimal salts | Sp | spectinomycin |
| SPM | stationary-phase mutagenesis | T90 | 90 min after the cessation of exponential growth |
| TBAB | tryptose blood agar base | TCR | transcription-couple repair |
| Tet | tetracycline | UV | ultraviolet light |
References
- Foster, P.L. Stress-Induced Mutagenesis in Bacteria. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 373–397. [Google Scholar] [CrossRef] [PubMed]
- Galhardo, R.S.; Hastings, P.J.; Rosenberg, S.M. Mutation as a stress response and the regulation of evolvability. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 399–435. [Google Scholar] [CrossRef] [PubMed]
- Robleto, E.A.; Yasbin, R.; Ross, C.; Pedraza-Reyes, M. Stationary phase mutagenesis in Bacillus subtilis: A paradigm to study genetic diversity programs in cells under stress. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Kivisaar, M. Mechanisms of stationary-phase mutagenesis in bacteria: Mutational processes in pseudomonads. FEMS Microbiol. Lett. 2010, 312, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Robleto, E.A.; Martin, H.A.; Vallin, C.; Pedraza-Reyes, M.; Yasbin, R. Transcription-mediated mutagenic processes. In Stress-Induced Mutagenesis; Mittelman, D., Ed.; Springer: Heidelberg, Germany, 2013; p. 275. [Google Scholar]
- Robleto, E.A.; Martin, H.A.; Pedraza-Reyes, M. Mfd and transcriptional derepression cause genetic diversity in Bacillus subtilis. Front. Biosci. 2012, 4, 1246–1254. [Google Scholar] [CrossRef]
- Wimberly, H.; Shee, C.; Thornton, P.C.; Sivaramakrishnan, P.; Rosenberg, S.M.; Hastings, P.J. R-loops and nicks initiate DNA breakage and genome instability in non-growing Escherichia coli. Nat. Commun. 2013. [Google Scholar] [CrossRef]
- Witkin, E.M. Radiation-induced mutations and their repair. Science 1966, 152, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Hanawalt, P.C.; Spivak, G. Transcription-coupled DNA repair: two decades of progress and surprises. Nat. Rev. Mol. Cell Biol. 2008, 9, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Deaconescu, A.M. RNA polymerase between lesion bypass and DNA repair. Cell. Mol. Life Sci. 2013, 70, 4495–4509. [Google Scholar] [CrossRef] [PubMed]
- Deaconescu, A.M.; Artsimovitch, I.; Grigorieff, N. Interplay of DNA repair with transcription: From structures to mechanisms. Trends Biochem. Sci. 2012, 37, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Deaconescu, A.M.; Chambers, A.L.; Smith, A.J.; Nickels, B.E.; Hochschild, A.; Savery, N.J.; Darst, S.A. Structural basis for bacterial transcription-coupled DNA repair. Cell 2006, 124, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Howan, K.; Smith, A.J.; Westblade, L.F.; Joly, N.; Grange, W.; Zorman, S.; Darst, S.A.; Savery, N.J.; Strick, T.R. Initiation of transcription-coupled repair characterized at single-molecule resolution. Nature 2012, 490, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Haines, N.M.; Kim, Y.-I.T.; Smith, A.J.; Savery, N.J. Stalled transcription complexes promote DNA repair at a distance. Proc. Natl. Acad. Sci. USA 2014, 111, 4037–4042. [Google Scholar] [CrossRef] [PubMed]
- Deaconescu, A.M.; Sevostyanova, A.; Artsimovitch, I.; Grigorieff, N. Nucleotide excision repair (NER) machinery recruitment by the transcription-repair coupling factor involves unmasking of a conserved intramolecular interface. Proc. Natl. Acad. Sci. USA 2012, 109, 3353–3358. [Google Scholar] [CrossRef] [PubMed]
- Brégeon, D.; Doddridge, Z.A.; You, H.J.; Weiss, B.; Doetsch, P.W. Transcriptional mutagenesis induced by uracil and 8-oxoguanine in Escherichia coli. Mol. Cell. 2003, 12, 959–970. [Google Scholar] [CrossRef]
- Clauson, C.L.; Saxowsky, T.T.; Doetsch, P.W. Dynamic flexibility of DNA repair pathways in growth arrested Escherichia coli. DNA Repair 2010, 9, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, E.R.; Walker, G.C.; Siede, W.R.D.; Schultz, R.A.; Ellenburger, T. DNA Repair and Mutagenesis, 2nd ed.; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Barajas-Ornelas, R.C.; Ramirez-Guadiana, F.H.; Juarez-Godinez, R.; Ayala-Garcia, V.M.; Robleto, E.A.; Yasbin, R.E.; Pedraza-Reyes, M. Error-prone processing of apurinic/apyrimidinic (AP) sites by PolX underlies a novel mechanism that promotes adaptive mutagenesis in Bacillus subtilis. J. Bacteriol. 2014, 196, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Bridges, B.A. Starvation-associated mutation in Escherichia coli strains defective in transcription repair coupling factor. Mutat. Res. 1995, 329, 49–56. [Google Scholar] [CrossRef]
- Millon-Weaver, S.; Samadpour, N.A.; Moreno-Hable, D.A.; Nugent, P.; Briinacher, M.J.; Weiss, E.; Hayden, H.S.; Miller, S.I.; Liachko, I.; Merrikh, H. An underlying mechanism for the increased mutagenesis of lagging-strand genes in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2015, 112, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Pybus, C.; Pedraza-Reyes, M.; Sung, H.M.; Yasbin, R.E.; Robleto, E. Novel role of mfd: Effects on stationary-phase mutagenesis in Bacillus subtilis. J. Bacteriol. 2006, 188, 7512–7520. [Google Scholar] [CrossRef] [PubMed]
- Briley, K.; Prepiak, P.; Dias, M.J.; Hahn, J.; Dubnau, D. Maf acts downstream of ComGA to arrest cell division in competent cells of B. subtilis. Mol. Microbiol. 2011, 81, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Veening, J.W.; Murray, H.; Errington, J. A mechanism for cell cycle regulation of sporulation initiation in Bacillus subtilis. Genes Dev. 2009, 23, 1959–1970. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.D.; Sanders, G.M.; Grossman, A.D. Nutritional Control of Elongation of DNA Replication by (p)ppgpp. Cell 2007, 128, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Yasbin, R.E. Adaptive, or stationary-Phase, mutagenesis, a component of bacterial differentiation in Bacillus subtilis. J. Bacteriol. 2002, 184, 5641–5653. [Google Scholar] [CrossRef] [PubMed]
- Hastings, P.J. Adaptive amplification. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Epshtein, V.; Kamarthapu, V.; McGary, K.; Svetlov, V.; Ueberheide, B.; Proshkin, S.; Mironov, A.; Nudler, E. UvrD facilitates DNA repair by pulling RNA polymerase backwards. Nature 2014, 505, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.E.; Lewis, C.A.; Mooney, R.A.; Kohanski, M.A.; Collins, J.J.; Landick, R.; Walker, G.C. Roles for the transcription elongation factor NusA in both DNA repair and damage tolerance pathways in Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 15517–15522. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.E.; Walker, G.C. The Transcription Elongation Factor NusA Is Required for Stress-Induced Mutagenesis in Escherichia coli. Curr. Biol. 2010, 20, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debora, B.N.; Vidales, L.E.; Ramírez, R.; Ramírez, M.; Robleto, E.A.; Yasbin, R.E.; Pedraza-Reyes, M. Mismatch repair modulation of MutY activity drives Bacillus subtilis stationary-phase mutagenesis. J. Bacteriol. 2011, 193, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Hans-Ulrich, H. Molecular nutrition, food research. In Methods for General and Molecular; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; p. 791. [Google Scholar]
- Duigou, S.; Ehrlich, S.D.; Noirot, P.; Noirot-Gros, M.F. DNA polymerase I acts in translesion synthesis mediated by the Y-polymerases in Bacillus subtilis. Mol. Microbiol. 2005, 57, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Guadiana, F.H.; del Carmen Barajas-Ornelas, R.; Ayala-García, V.M.; Yasbin, R.E.; Robleto, E.A.; Pedraza-Reyes, M. Transcriptional coupling of DNA repair in sporulating Bacillus subtilis cells. Mol. Microbiol. 2013, 90, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Vagner, V.; Dervyn, E.; Ehrlich, S.D. A vector for systematic gene inactivation in Bacillus subtilis. Microbiology 1998, 144, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Spizizen, J. Transformation of biochemically deficient strain of Bacillus subtilis by deoxiribonucleate. Proc. Natl. Acad. Sci. USA 1958, 44, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Lea, D.E.; Coulson, C.A. The distribution of the numbers of mutants in bacterial populations. J. Genet. 1949, 49, 264–285. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-reyes, M.; Yasbin, R.E. Contribution of the mismatch DNA repair system to the generation of stationary-phase-induced mutants of Bacillus subtilis. J. Bacteriol. 2004, 186, 6485–6491. [Google Scholar] [CrossRef] [PubMed]
- Rosche, W.A.; Foster, P.L. Determining mutation rates in bacterial populations. Methods 2000, 20, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Setlow, P. Sporulation, germination and outgrowth. In Molecular Biological Methods for Bacillus, 2nd ed.; Harwood, C.R., Cutting, S.M., Eds.; John Wiley & Sons: Sussex, UK, 1990. [Google Scholar]
- Fukushima, S.; Itaya, M.; Kato, H.; Ogasawara, N.; Yoshikawa, H. Reassessment of the in vivo functions of DNA polymerase I and RNase H in bacterial cell growth. J. Bacteriol. 2007, 189, 8575–8583. [Google Scholar] [CrossRef] [PubMed]
- Ollington, J.F.; Haldenwang, W.G.; Huynh, T.V.; Losick, R. Developmentally regulated transcription in a cloned segment of the Bacillus subtilis chromosome. J. Bacteriol. 1981, 147, 432–442. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vidales, L.E.; Cárdenas, L.C.; Robleto, E.A.; Yasbin, R.E.; Pedraza-Reyes, M. Defects in the error prevention oxidized guanine system potentiate stationary-phase mutagenesis in Bacillus subtilis. J. Bacteriol. 2009, 191, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Pybus, C.; Pedraza-Reyes, M.; Ross, C.A.; Martin, H.; Ona, K.; Yasbin, R.E.; Robleto, E.A. Transcription-associated mutation in Bacillus subtilis cells under stress. J. Bacteriol. 2010, 192, 3321–3328. [Google Scholar] [CrossRef] [PubMed]
- Branum, M.E.; Reardon, J.T.; Sancar, A. DNA repair excision nuclease attacks undamaged DNA: A potential source of spontaneous mutations. J. Biol. Chem. 2001, 276, 25421–25426. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Yoshiyama, K.; Maki, H. Spontaneous mutagenesis associated with nucleotide excision repair in Escherichia coli. Genes Cells 2008, 13, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Duigou, S.; Ehrlich, S.D.; Noirot, P.; Noirot-Gros, M.F. Distinctive genetic features exhibited by the Y-family DNA polymarases in Bacillus subtilis. Mol. Microbiol. 2004, 54, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Pope, M.A.; Porello, S.L.; David, S.S. Escherichia coli apurinic-apyrimidinic endonucleases enhance the turnover of the adenine glycosylase MutY with G:A substrates. J. Biol. Chem. 2002, 277, 22605–22615. [Google Scholar] [CrossRef] [PubMed]
- Porello, S.L.; Leyes, A.E.; David, S.S. Single-turnover and pre-steady-state kinetics of the reaction of the adenine glycosylase MutY with mismatch-containing DNA substrates. Biochemistry 1998, 37, 14756–14764. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Sahin, O.; Barton, Y.W.; Zhang, Q. Key role of Mfd in the development of fluoroquinolone resistance in Campylobacter jejuni. PLoS Pathog. 2008. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.N.; Rodin, A.S.; Juhasz, A.; Holmquist, G.P. Cancerous hyper-mutagenesis in p53 genes is possibly associated with transcriptional bypass of DNA lesions. Mutat. Res. 2002, 510, 153–168. [Google Scholar] [CrossRef]
- Doetsch, P.W. Translesion synthesis by RNA polymerases: occurrence and biological implications for transcriptional mutagenesis. Mutat. Res. 2002, 510, 131–140. [Google Scholar] [CrossRef]
- Holmquist, G.P. Cell-selfish modes of evolution and mutations directed after transcriptional bypass. Mutat. Res. 2002, 510, 141–152. [Google Scholar] [CrossRef]
- Lee, G.H.; Jeong, J.Y.; Chung, J.W.; Nam, W.H.; Lee, S.M.; Pak, J.H.; Choi, K.D.; Song, H.J.; Jung, H.Y.; Kim, J.H. The Helicobacter pylori Mfd protein is important for antibiotic resistance and DNA repair. Diagn. Microbiol. Infect. Dis. 2009, 65, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, T.D.; Flett, K.B.; Yelin, I.; Martin, T.R.; McAdam, A.J.; Priebe, G.P.; Kishony, R. Genetic variation of a bacterial pathogen within individuals with cystic fibrosis provides a record of selective pressures. Nat. Genet. 2014, 46, 82–87. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant genotype | Reference or source |
| YB955 | hisC952 metB5 leuC427 xin-1 SpSENS | [26] |
| YB9801 | YB955 mfd::tet, Tet r | [22] |
| YB9900 | YB955 uvrA::em, Em r | a BSU35160 → YB955 |
| JJS164 | TF8a ΔpolA::sp, Sp r | [33] |
| AMP100 | YB955 ΔpolA::sp, Sp r | a JJS164 → YB955 |
| AMP101 | YB955 ΔpolA::sp, mfd::tet, Sp r Tet r | a JJS164 → YB9801 |
| BSU35160 | 168 uvrA::em, Em r | Bacillus Genetic Stock Center (BKE Knock-out collection) |
| HAM300 | YB955 uvrA::emmfd::tet, Em r Tet r | a YB9900 → YB9801 |
| PERM598 | 168 mutY::sp, Sp r | [31] |
| PERM687 | YB955 uvrA::sp, Sp r | Peter Setlow |
| PERM704 | YB955 mutY::sp, Sp r | [31] |
| PERM899 | YB955 mutY:: sp amyE::Phs-mutY, Cm r | [31] |
| PERM818 | YB955 mfd::tet mutY::sp, Tet r Sp r | a PERM598 → YB9801 |
| PERM995 | YB955 mfd::tet mutY::sp amyE::Phs-mutY, Cm r | b pPERM852 → PERM818 |
| PERM1029 | YB955 mutY::em, Em r | b pPERM979 → YB955 |
| PERM1041 | YB955 mfd::tet mutY::em, Tet r Em r | b pPERM979 → YB9801 |
| PERM1042 | YB955 mfd::tet mutY::em amyE::Phs-mfd, Tet r Em r Spr | b pPERM1041 → PERM1041 |
| PERM1046 | YB955 mfd::tet mutY::em amyE::Phs, Tet r Em r Sp r | b pdrE amyE → PERM1041 |
| PERM1352 | YB955 mutY::em uvrA::sp, Em r Sp r | a PERM1029 → PERM687 |
| PERM1353 | YB955 mfd::tet mutY::em uvrA::150, Tet r Em r | a PERM1029 → PERM1352 |
| PERM1123 | 168 pMUTIN4::mfd-lacZ, Em r. | [34] |
| Plasmids | Description | Reference or source |
| pHyperspank | Integrative vector, Sp r | David Rudner |
| pDR244 | cre, temperature-sensitive replication Amp r Sp r | Bacillus Genetic Stock Center (BKE Knock-out collection) |
| pPERM852 | pHyperspank-mutY Integrative vector with mutY control by IPTG, Amp r Cm r | [31] |
| pPERM979 | pMUTIN4::mutY, Amp r Em r | This study; [35] |
| pPERM1043 | pHyperspank-mfd Integrative vector with mfd under the control of IPTG, Amp r Sp r | This study |
| pPERM1084 | pBT-mutY, Cm r | This study |
| pPERM1072 | pTRG-mfd, Tet r | This study |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Marroquín, M.; Martin, H.A.; Pepper, A.; Girard, M.E.; Kidman, A.A.; Vallin, C.; Yasbin, R.E.; Pedraza-Reyes, M.; Robleto, E.A. Stationary-Phase Mutagenesis in Stressed Bacillus subtilis Cells Operates by Mfd-Dependent Mutagenic Pathways. Genes 2016, 7, 33. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070033
Gómez-Marroquín M, Martin HA, Pepper A, Girard ME, Kidman AA, Vallin C, Yasbin RE, Pedraza-Reyes M, Robleto EA. Stationary-Phase Mutagenesis in Stressed Bacillus subtilis Cells Operates by Mfd-Dependent Mutagenic Pathways. Genes. 2016; 7(7):33. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070033
Chicago/Turabian StyleGómez-Marroquín, Martha, Holly A. Martin, Amber Pepper, Mary E. Girard, Amanda A. Kidman, Carmen Vallin, Ronald E. Yasbin, Mario Pedraza-Reyes, and Eduardo A. Robleto. 2016. "Stationary-Phase Mutagenesis in Stressed Bacillus subtilis Cells Operates by Mfd-Dependent Mutagenic Pathways" Genes 7, no. 7: 33. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070033