
Prokaryotic Expression and Serodiagnostic Potential of Glyceraldehyde-3-Phosphate Dehydrogenase and Thioredoxin Peroxidase from Baylisascaris schroederi

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasites and Animals
2.2. Serum and Secondary Antibody
2.3. Ethics Statement
2.4. RNA Extraction and Amplification of Baylisascaris schroederi Thioredoxin Peroxidaseand Glyceraldehyde-3-Phosphate Dehydrogenase
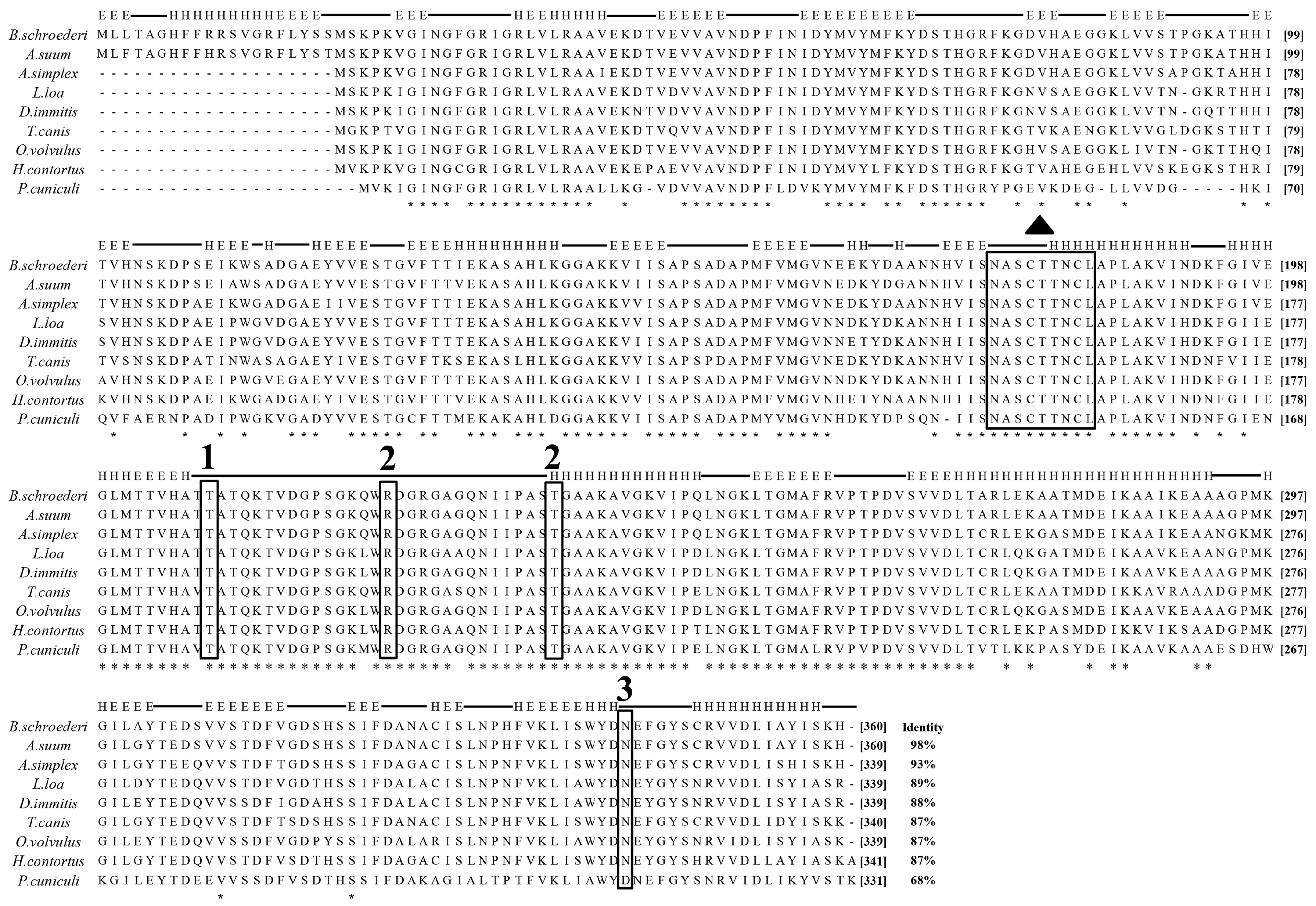
2.5. Sequence and Bioinformatic Analysis
2.6. Expression and Purification of Baylisascaris schroederi Thioredoxin Peroxidase and Glyceraldehyde-3-Phosphate Dehydrogenase
2.7. Preparation of Polyclonal Antibodies
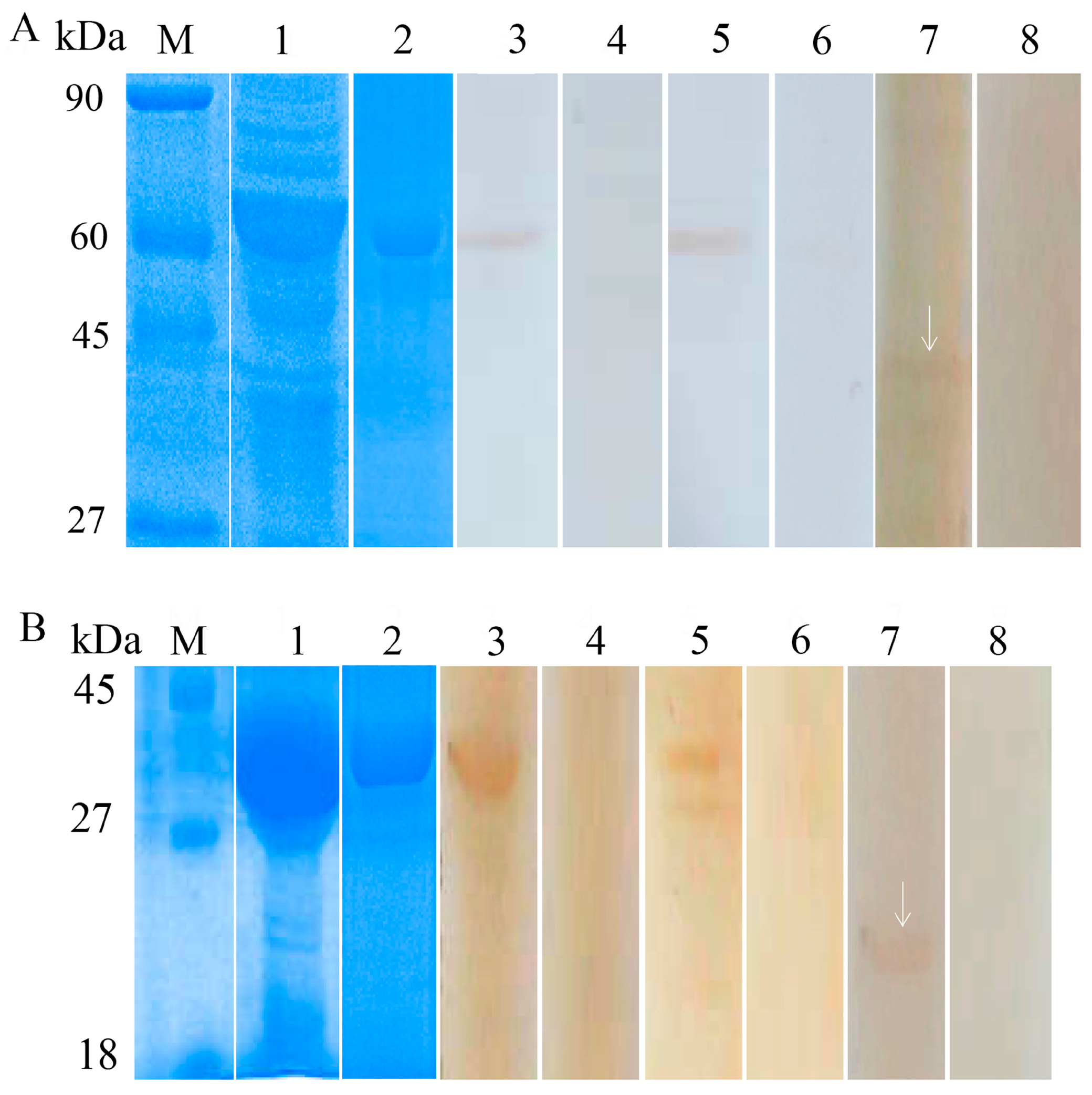
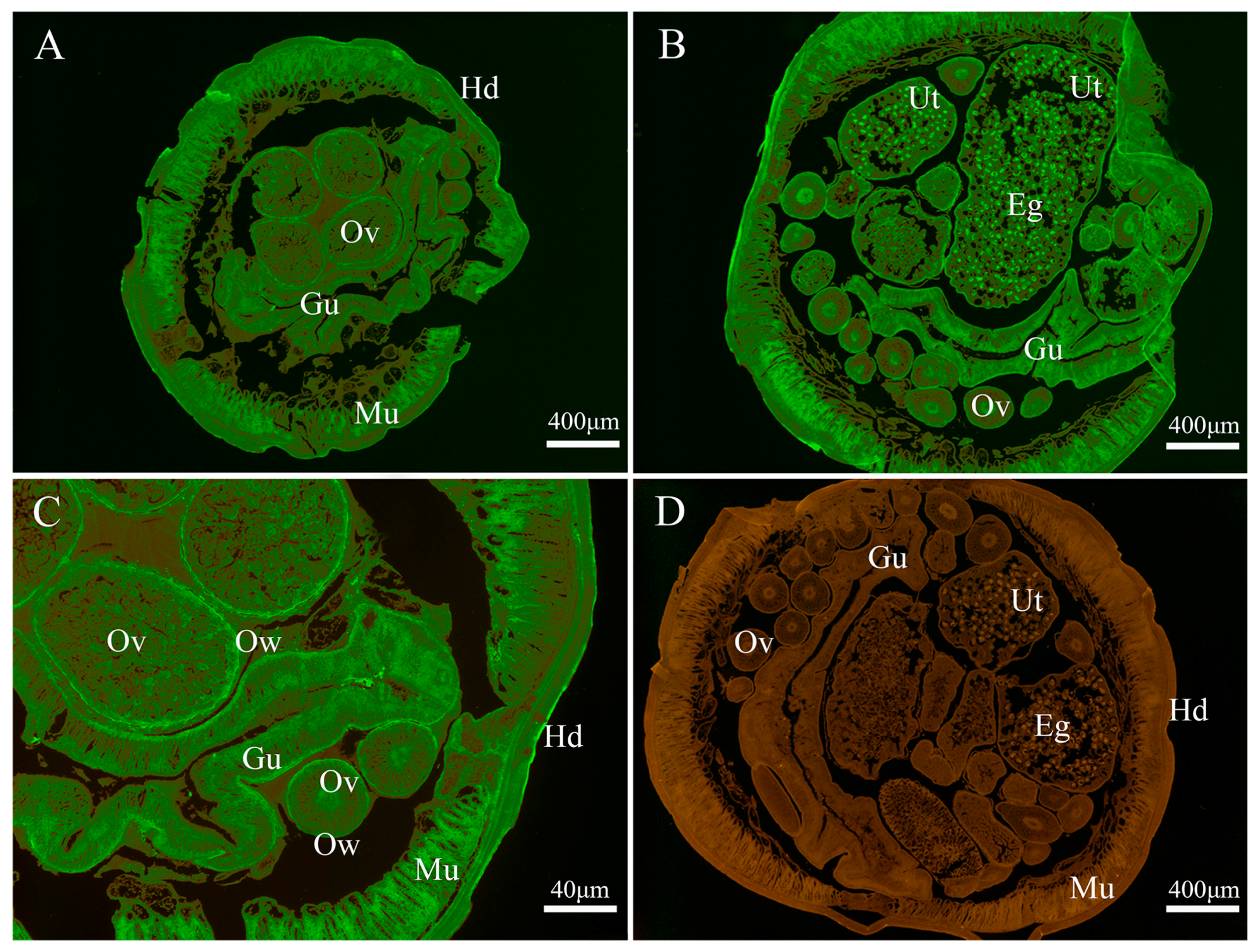
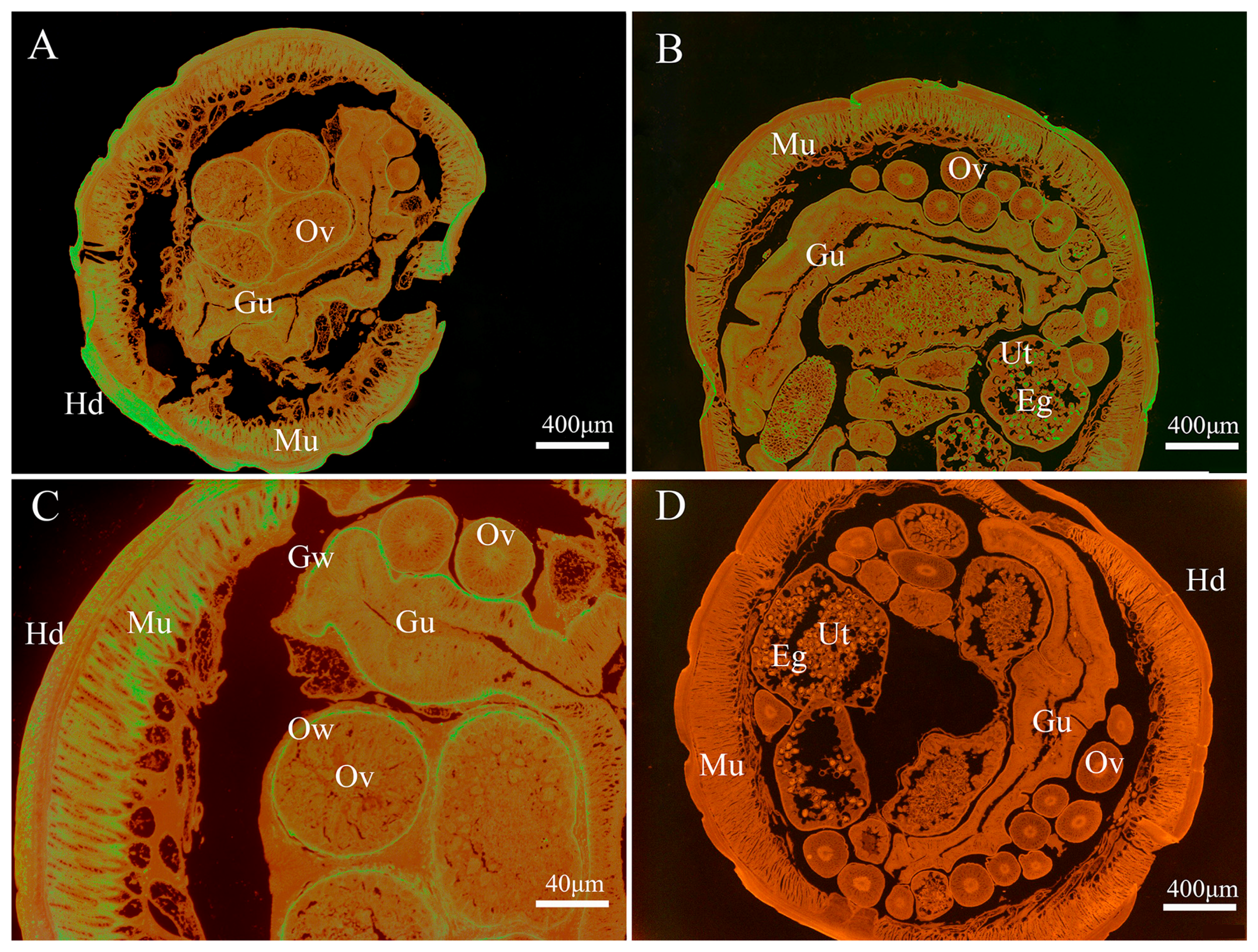
2.8. Western Blot Analysis and Immunolocalization
2.9. Development of Indirect ELISA
2.10. Antibody Titer Detection of Mouse Serum
2.11. Repeatability and Reproducibility of the Indirect ELISA
2.12. Sensitivity and Specificity Analysis of the ELISA
2.13. Statistical Analysis
3. Results
3.1. Molecular and Characterization of Baylisascaris schroederi Thioredoxin Peroxidase and Glyceraldehyde-3-Phosphate Dehydrogenase
3.2. Expression and Identification of the Recombinant Proteins
3.3. Immunolocalization of Endogenous Baylisascaris schroederi Thioredoxin Peroxidase and Glyceraldehyde-3-Phosphate Dehydrogenase
3.4. Establishment of Indirect ELISA
3.5. Detection of the Antibody Titers of Mice Infected with Baylisascaris schroederi
3.6. Repeatability and Reproducibility of the Indirect ELISA
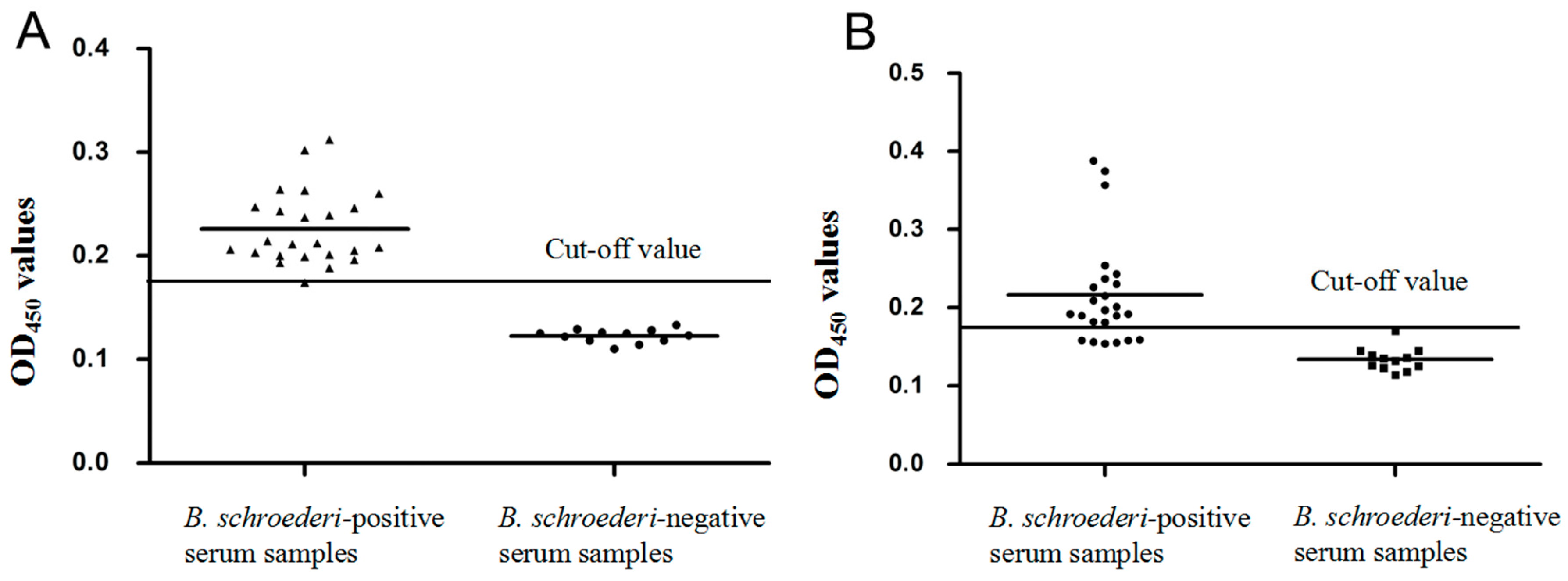
3.7. Specificity and Sensitivity of the Indirect ELISA
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, J.S.; Daszak, P.; Huang, H.L.; Yang, G.Y.; Kilpatrick, A.M.; Zhang, S. Parasite Threat to Panda Conservation. EcoHealth 2008, 5, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; He, G.Z.; Yang, G.Y.; Fei, Y.X.; Zhang, Z.H.; Wang, C.D.; Yang, Z.; Lan, J.C.; Luo, L.; Liu, L. Cloning, expression and evaluation of the efficacy of a recombinant Baylisascaris schroederi Bs-Ag3 antigen in mice. Vaccine 2008, 26, 6919–6924. [Google Scholar] [CrossRef] [PubMed]
- Beaver, P.C. The nature of visceral larva migrans. J. Parasitol. 1969, 55, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Li, D.S.; He, Y.; Wu, H.L.; Wang, C.D.; Li, C.W.; Lan, J.C.; Cheng, Z.Q.; Xie, Y.; Han, H.Y.; Yang, G.Y.; Wang, C.D. Prevalence of Helminths in Captive Giant Pandas. J. Econ. Anim. 2014, 18, 214–216. [Google Scholar]
- Wu, J.; Hu, H. Ascaridosis of giant panda (Ailuropoda melanoleuca). Chin. J. Wildl. 1985, 5, 42–43. [Google Scholar]
- Loeffler, I.K.; Montali, R.J.; Rideout, B.A. Diseases and Pathology of Giant Pandas. In Giant Pandas: Biology, Veterinary Medicine and Management; Cambridge University Press: Cambridge, UK, 2006; pp. 377–409. [Google Scholar]
- Qiu, X.; Mainka, S.A. Review of Mortality of the Giant Panda (Ailuropoda melanoleuca). J. Zoo Wildl. Med. 1993, 24, 425–429. [Google Scholar]
- Zhou, X.; Yu, H.; Wang, N.; Xie, Y.; Liang, Y.N.; Li, D.S.; Wang, C.D.; Chen, S.J.; Yan, Y.B.; Gu, X.B.; et al. Molecular diagnosis of Baylisascaris schroederi infections in giant panda (Ailuropoda melanoleuca) feces using PCR. J. Wildl. Dis. 2015, 49, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.P.; Yie, S.M.; Yue, B.S.; Zhou, J.L.; An, R.X.; Yang, J.D.; Chen, W.L.; Wang, C.D.; Zhang, L.; Shen, F.J.; et al. Determination of Baylisascaris schroederi Infection in Wild Giant Pandas by an Accurate and Sensitive PCR/CE-SSCP Method. PLoS ONE 2012, 7, e41995. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Li, D.S.; Zhou, X.; Xie, Y.; Liang, Y.N.; Wang, C.D.; Yu, H.; Chen, S.J.; Yan, Y.B.; Gu, X.B.; et al. A sensitive and specific PCR assay for the detection of Baylisascaris schroederi eggs in giant panda feces. Parasitol. Int. 2013, 62, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A.; Schröder, E.; Robin Harris, J.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Lim, Y.S.; Cha, M.K.; Kim, H.K.; Uhm, T.B.; Park, J.W.; Kim, K.; Kim, I.H. Removals of Hydrogen Peroxide and Hydroxyl Radical by Thiol-Specific Antioxidant Protein as a Possible Role in Vivo. Biochem. Biophys. Res. Commun. 1993, 192, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C.; Röllinghoff, M.; Diefenbach, A. Reactive oxygen and reactive nitrogen intermediates in innate and specific immunity. Curr. Opin. Immunol. 2000, 12, 64–76. [Google Scholar] [CrossRef]
- McEligot, A.J.; Yang, S.; Meyskens, F.L., Jr. Redox regulation by intrinsic species and extrinsic nutrients in normal and cancer cells. Annu. Rev. Nutr. 2005, 25, 261–295. [Google Scholar] [CrossRef] [PubMed]
- Angeles, J.M.; Goto, Y.; Kirinoki, M.; Asada, M.; Leonardo, L.R.; Rivera, P.T.; Villacorte, E.A.; Inoue, N.; Chigusa, Y.; Kawazu, S. Utilization of ELISA Using Thioredoxin Peroxidase-1 and Tandem Repeat Proteins for Diagnosis of Schistosoma japonicum Infection among Water Buffaloes. PLoS Negl. Trop. Dis. 2012, 6, e1800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Rogniaux, H.; Huang, W.; Chauvin, A.; Moreau, E. Analysis of thioredoxin peroxidase as a promising antigen for diagnosis of Fasciola gigantica infection A preliminary study. Parasitol. Int. 2010, 60, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Asahi, H.; Botkin, D.J.; Stadecker, M.J. Schistosome infection stimulates host CD4 (+) T helper cell and B-cell responses against a novel egg antigen, thioredoxin peroxidase. Infect. Immun. 2001, 69, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Sirover, M.A. New insights into an old protein: The functional diversity of mammalian glyceraldehyde-3-phosphate dehydrogenase. Biochim. Biophys. Acta 1999, 1432, 159. [Google Scholar] [CrossRef]
- Viscogliosi, E.; Müller, M. Phylogenetic Relationships of the Glycolytic Enzyme, Glyceraldehyde-3-Phosphate Dehydrogenase, from Parabasalid Flagellates. J. Mol. Evol. 1998, 47, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, J.L.; Sirover, M.A. Subcellular localization of human glyceraldehyde-3-phosphate dehydrogenase is independent of its glycolytic function. Biochim. Biophys. Acta 2003, 1622, 50–56. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, F.; Yu, C.X.; Xiao, D.; Song, L.J.; Yin, X.R.; Shen, S.; Hua, W.Q.; Zhang, J.Z.; Zhang, H.F.; et al. Identification of proteins inducing short-lived antibody responses from excreted/secretory products of Schistosoma japonicum adult worms by immunoproteomic analysis. J. Proteom. 2013, 87, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cui, S.J.; Hu, W.; Feng, Z.; Wang, Z.Q.; Han, Z.G. Excretory/secretory proteome of the adult developmental stage of human blood fluke, Schistosoma japonicum. Mol. Cell. Proteom. 2009, 8, 1236–1251. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, F.; Yu, C.X.; Xiao, D.; Song, L.J.; Yin, X.R.; Shen, S.; Hua, W.Q.; Zhang, J.Z.; Zhang, H.F.; et al. The value of diagnosing schistosomiasis by detecting IgG antibodies against glyceraldehyde-3-phosphate dehydrogenase of Schistosoma japonicum. J. Pathog. Biol. 2015, 10, 976–979. [Google Scholar]
- Zhang, H.; Zhang, Z.L. Diagnosis and treatment of giant panda ascarids. Gansu Anim. Vet. Sci. 2002, 32, 25–26. [Google Scholar]
- Xie, Y.; Xu, J.; Zhou, X.; Gu, X.B.; Yang, G.Y. The genome and transcriptome of the giant panda roundworm Baylisascaris schroederi. (unpublished).
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, S.; Yan, Y.; Zhang, Z.; Li, D.; Yu, H.; Wang, C.; Nong, X.; Zhou, X.; Gu, X.; et al. Potential of recombinant inorganic pyrophosphatase antigen as a new vaccine candidate against Baylisascaris schroederi in mice. Vet. Res. 2013, 44, 90. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Niu, D.S.; Zhang, X.Y.; Chen, M.L.; Cui, H.; Wei, W.J.; Wen, B.H.; Chen, X.R. Recombinant 56-Kilodalton major outer membrane protein antigen of Orientia tsutsugamushi Shanxi and its antigenicity. Infect. Immun. 2003, 71, 4772–4779. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, L.; Yang, Y.; Gu, X.; Wang, Y.; Lai, W.; Peng, X.; Yang, G. Expression, tissue localization and serodiagnostic potential ofTaenia multiceps acidic ribosomal protein P2. Parasites Vectors 2015, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Liddell, S.; Knox, D.P. Extracellular and cytoplasmic Cu/Zn superoxide dismutases from Haemonchus contortus. Parasitology 1998, 116, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wei, T.; Liu, L.N.; Zhang, X.; Qi, X.; Zhang, Z.F.; Wang, Z.Q. Molecular characterization of a Spirometra mansoni antigenic polypeptide gene encoding a 28.7 kDa protein. Parasitol. Res. 2014, 113, 3511. [Google Scholar] [CrossRef] [PubMed]
- Erlwein, O.; Robinson, M.J.; Kaye, S.; Wills, G.; Izui, S.; Wessely, S.; Weber, J.; Cleare, A.; Collier, D.; McClure, M.O. Investigation into the Presence of and Serological Response to XMRV in CFS Patients. PLoS ONE 2012, 6, e17592. [Google Scholar] [CrossRef] [PubMed]
- Breathnach, R.; Chambon, P. Organization and Expression of Eucaryotic Split Genes Coding for Proteins. Annu. Rev. Biochem. 1981, 50, 349–383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.; Wu, H.; Gu, X.; Hu, Y.; Wei, F. The parasites of giant pandas: individual-based measurement in wild animals. J. Wildl. Dis. 2011, 47, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Jolodar, A.; Fischer, P.; Bergmann, S.; Büttner, D.W.; Hammerschmidt, S.; Brattig, N.W. Molecular cloning of an alpha-enolase from the human filarial parasite Onchocerca volvulus that binds human plasminogen. Biochim. Biophys. Acta 2003, 1627, 111–120. [Google Scholar] [CrossRef]
- Hou, Q.L.; Wang, H.; Zhang, Z.Z.; Cao, W.Y.; Zhang, F.C.; Zhang, W.B. Immunolocalization of the antioxidant protein TPx of Echinococcus granulosus. Chin. J. Cell. Mol. Immunol. 2007, 23, 998. [Google Scholar]
- Yang, G.Y.; Zhang, Z.H. Parasitic Diseases of Wildlife; Beijing Science Press: Beijing, China, 2013; pp. 526–538. [Google Scholar]
- Montali, R.J.; Bartz, C.R.; Teare, J.A.; Allen, J.T.; Appel, M.J.; Bush, M. Clinical trials with canine distemper vaccines in exotic carnivores. J. Am. Vet. Med. Assoc. 1983, 183, 1163–1167. [Google Scholar] [PubMed]
- Wang, C.D.; Yang, S.T.; Wu, K.B.; Gao, Y.W.; Zhang, Z.H.; Luo, L.; Wang, W.Y.; Wang, T.C.; Yan, Y.B.; Hu, J.; et al. Serological evaluation of the efficacy of the multivalent canine distemper attenuated live vaccines on giant pandas (Ailuropoda melanoleuca). Acta Theriol. Sin. 2008, 28, 212–216. [Google Scholar]
- Cherpillod, P.; Tipold, A.; Griot-Wenk, M.; Cardozo, C.; Schmid, I.; Fatzer, R.; Schobesberger, M.; Zurbriggen, R.; Bruckner, L.; Roch, F.; et al. Immune protection in dogs inoculated by CDV DNA vaccine. Chin. J. Vet. Sci. 2007, 32, 37–39. [Google Scholar]
- Chen, W.; Peng, S.M.; Chen, X.J.; Huang, M.; Cai, Q.H. Comparison of the Immune Effects of Several Canine Distemper Vaccines on Red Panda. Guangdong J. Anim. Vet. Sci. 2007, 32, 37–39. [Google Scholar]
- Vlaminck, J.; Nejsum, P.; Vangroenweghe, F.; Thamsborg, S.M.; Vercruysse, J.; Geldhof, P. Evaluation of a serodiagnostic test using Ascaris suum haemoglobin for the detection of roundworm infections in pig populations. Vet. Parasitol. 2012, 189, 267. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Sun, Y.; Gu, X.; Xie, Y.; Lai, W.; Jing, B.; Peng, X.; Yang, G. Prokaryotic Expression and Serodiagnostic Potential of Glyceraldehyde-3-Phosphate Dehydrogenase and Thioredoxin Peroxidase from Baylisascaris schroederi. Genes 2017, 8, 293. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110293
Li Y, Sun Y, Gu X, Xie Y, Lai W, Jing B, Peng X, Yang G. Prokaryotic Expression and Serodiagnostic Potential of Glyceraldehyde-3-Phosphate Dehydrogenase and Thioredoxin Peroxidase from Baylisascaris schroederi. Genes. 2017; 8(11):293. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110293
Chicago/Turabian StyleLi, Yu, Ying Sun, Xiaobin Gu, Yue Xie, Weiming Lai, Bo Jing, Xuerong Peng, and Guangyou Yang. 2017. "Prokaryotic Expression and Serodiagnostic Potential of Glyceraldehyde-3-Phosphate Dehydrogenase and Thioredoxin Peroxidase from Baylisascaris schroederi" Genes 8, no. 11: 293. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110293