
The Stearoyl-CoA Desaturase-1 (Desat1) in Drosophila cooperated with Myc to Induce Autophagy and Growth, a Potential New Link to Tumor Survival

 , and
, and
Abstract
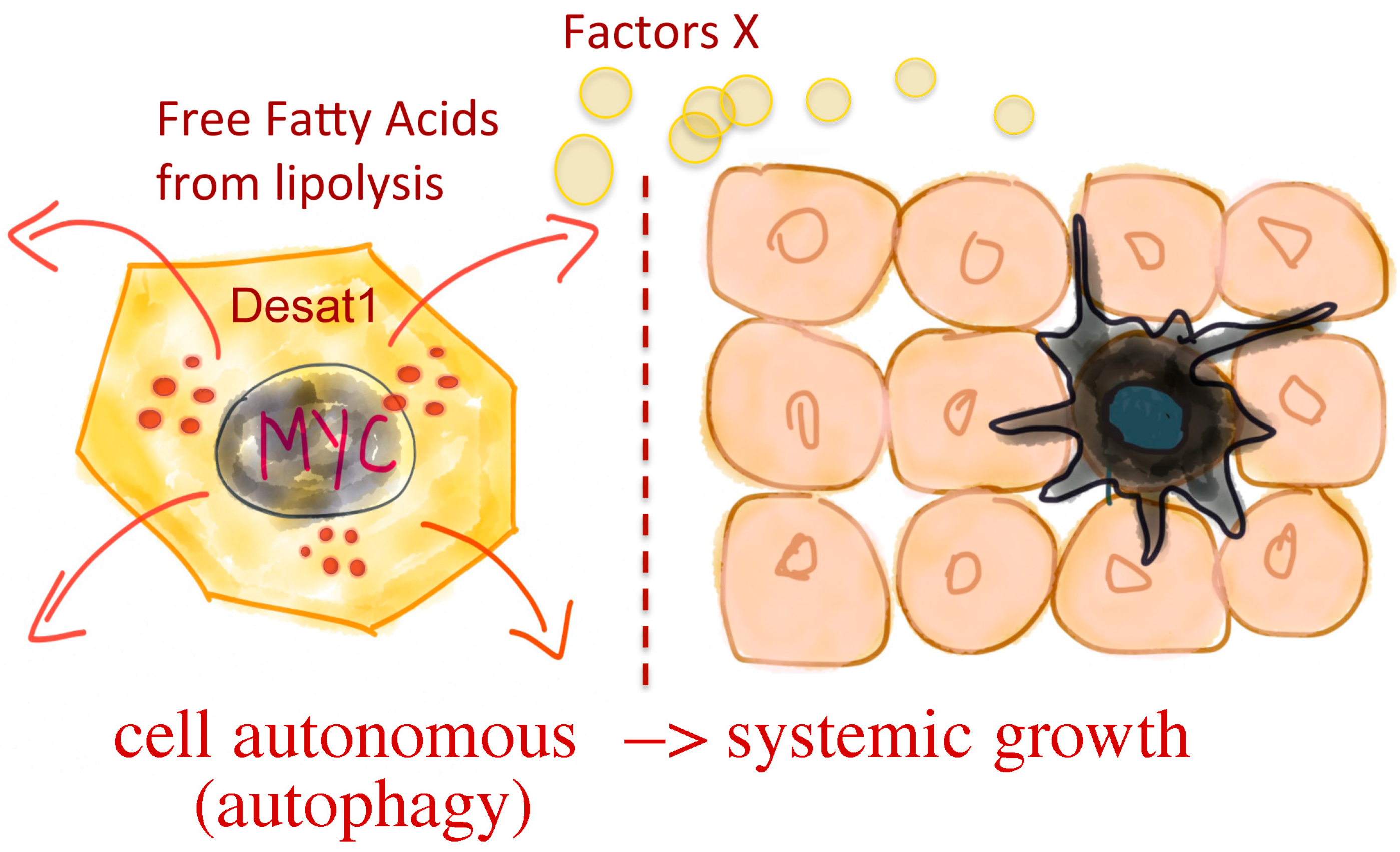
:1. Introduction
2. Materials and Methods
2.1. Fly Husbandry
2.2. Proteomics
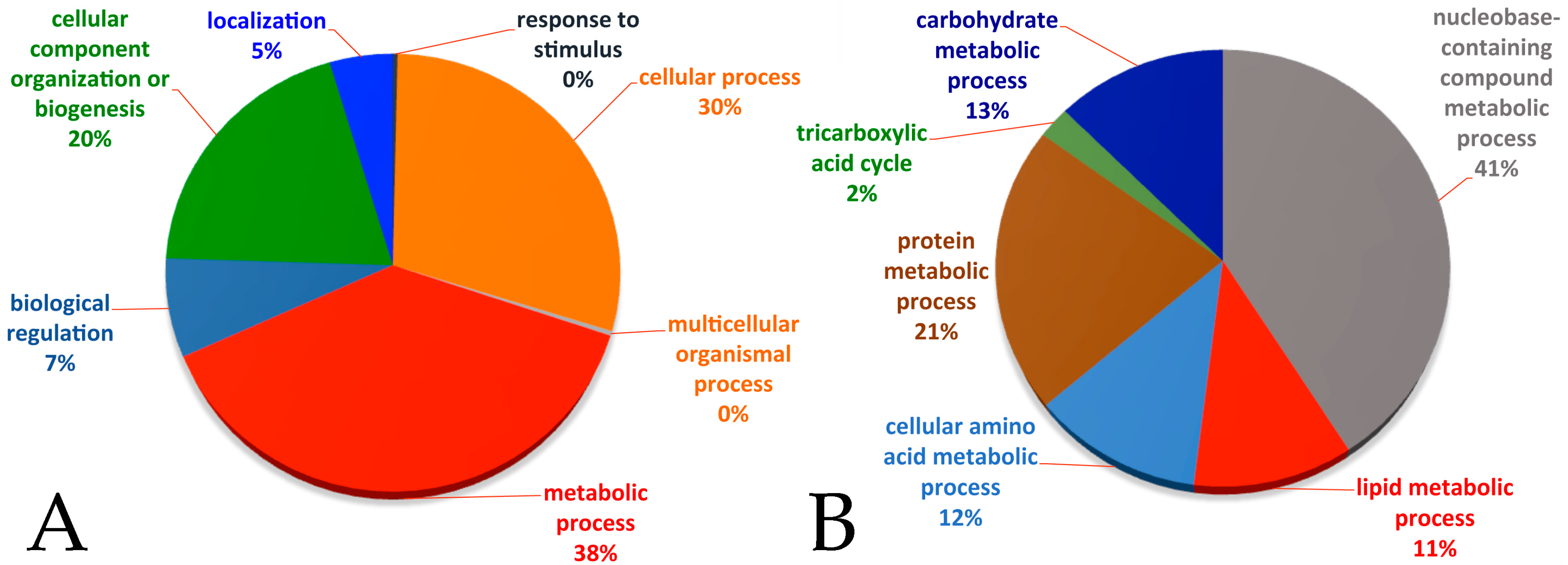
2.3. Gene Ontology Analysis
2.4. Generation of Inducible Flip-Out Clones and Clonal Analysis
2.5. Starvation Assay
2.6. Western Blot
2.7. Lysotracker Staining
2.8. Paraffin Embedding and Immunostaining of Human Samples
3. Results
3.1. Proteomic Profile from Fat Bodies Expressing Myc
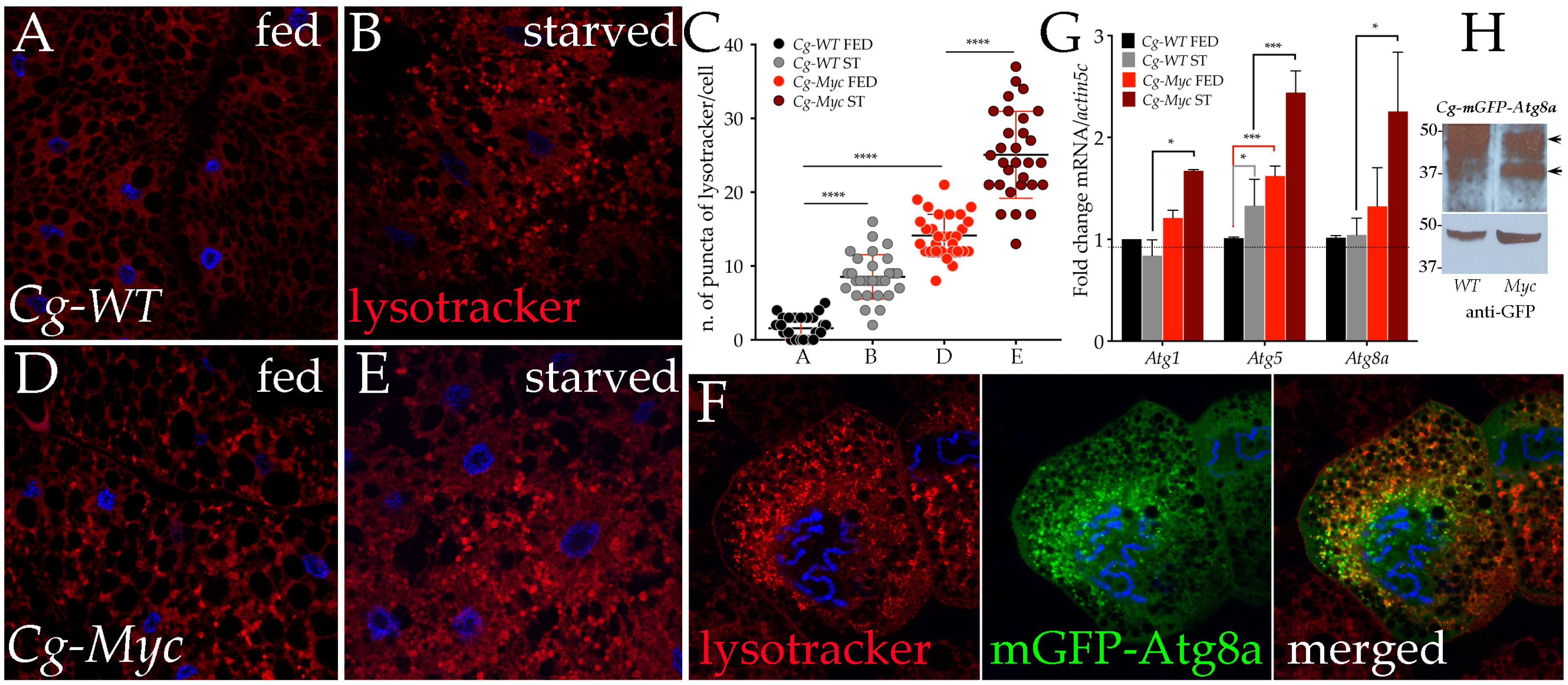
3.2. Myc Increases the Expression of Autophagic Genes and Induces Autophagy in the Fat Body from Animals Under Starvation
3.3. Physiological Levels of Desat1 Are Necessary for Myc-Induced Autophagy
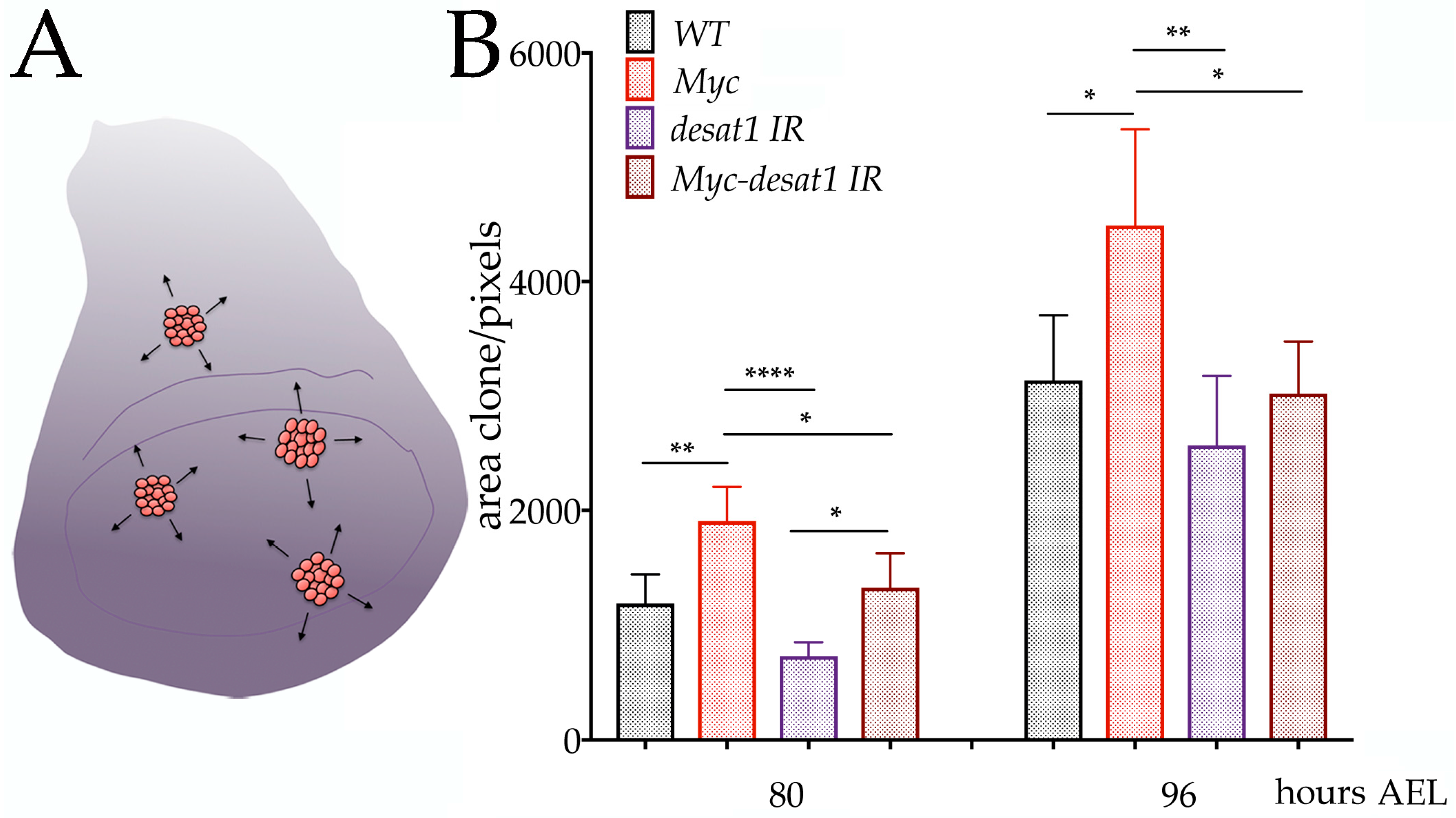
3.4. Reduction of Desat1 Affects the Ability of Myc to Induce Growth in Epithelial Cells
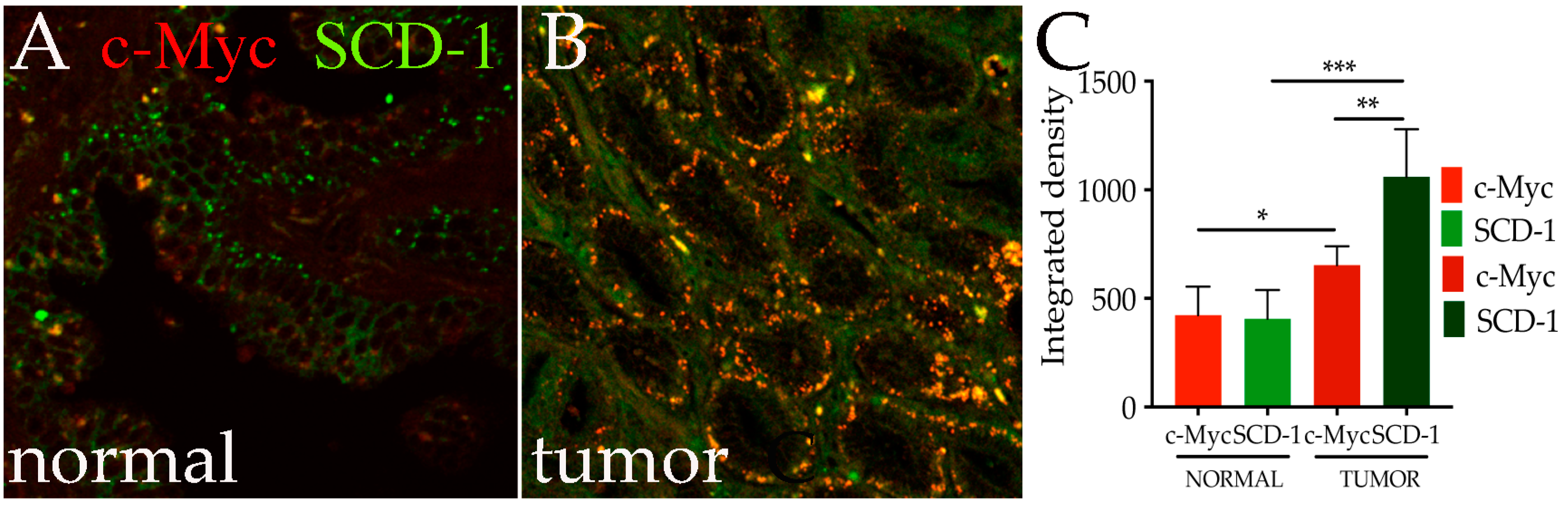
3.5. c-Myc and Human SCD-1 Are Co-Expressed in Epithelial Cells of Prostate Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hsieh, A.L.; Dang, C.V. Myc, metabolic synthetic lethality, and cancer. Recent Results Cancer Res. 2016, 207, 73–91. [Google Scholar] [PubMed]
- Grifoni, D.; Bellosta, P. Drosophila myc: A master regulator of cellular performance. Biochim. Biophys. Acta 2015, 1849, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Riccardo, S.; Daniel, M.; Saqcena, M.; Kundu, N.; Pession, A.; Grifoni, D.; Stocker, H.; Tabak, E.; Bellosta, P. Drosophila insulin and target of rapamycin (TOR) pathways regulate GSK3 beta activity to control myc stability and determine myc expression in vivo. BMC Biol. 2011, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Lipovsky, A.I.; Dibble, C.C.; Sahin, M.; Manning, B.D. S6K1 regulates GSK3 under conditions of mTOR-dependent feedback inhibition of akt. Mol. Cell 2006, 24, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Galletti, M.; Riccardo, S.; Parisi, F.; Lora, C.; Saqcena, M.K.; Rivas, L.; Wong, B.; Serra, A.; Serras, F.; Grifoni, D.; et al. Identification of domains responsible for ubiquitin-dependent degradation of d-Myc by glycogen synthase kinase 3β and casein kinase 1 kinases. Mol. Cell. Biol. 2009, 29, 3424–3434. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yang, X.; Xi, Y. Fat body remodeling and homeostasis control in drosophila. Life Sci. 2016, 167, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Riccardo, S.; Zola, S.; Lora, C.; Grifoni, D.; Brown, L.M.; Bellosta, P. Dm-yc expression in the fat body affects dilp2 release and increases the expression of the fat desaturase desat1 resulting in organismal growth. Dev. Biol. 2013, 379, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Teleman, A.A.; Chen, Y.W.; Cohen, S.M. Drosophila melted modulates foxo and tor activity. Dev. Cell 2005, 9, 271–281. [Google Scholar] [CrossRef] [PubMed]
- De la Cova, C.; Senoo-Matsuda, N.; Ziosi, M.; Wu, D.C.; Bellosta, P.; Quinzii, C.M.; Johnston, L.A. Supercompetitor status of drosophila myc cells requires p53 as a fitness sensor to reprogram metabolism and promote viability. Cell Metab. 2014, 19, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Claveria, C.; Torres, M. Cell competition: Mechanisms and physiological roles. Annu. Rev. Cell Dev. Biol. 2016, 32, 411–439. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, A.; Bowling, S.; Rodriguez, T.A. Cell competition and its role in the regulation of cell fitness from development to cancer. Dev. Cell 2016, 38, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Baenke, F.; Peck, B.; Miess, H.; Schulze, A. Hooked on fat: The role of lipid synthesis in cancer metabolism and tumour development. Dis. Model. Mech. 2013, 6, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Noto, A.; Raffa, S.; de Vitis, C.; Roscilli, G.; Malpicci, D.; Coluccia, P.; Di Napoli, A.; Ricci, A.; Giovagnoli, M.R.; Aurisicchio, L.; et al. Stearoyl-CoA desaturase-1 is a key factor for lung cancer-initiating cells. Cell Death Dis. 2013, 4, e947. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, N.; Swinnen, J.V.; Smans, K. ATP-citrate lyase: A key player in cancer metabolism. Cancer Res. 2012, 72, 3709–3714. [Google Scholar] [CrossRef] [PubMed]
- Von Roemeling, C.A.; Marlow, L.A.; Wei, J.J.; Cooper, S.J.; Caulfield, T.R.; Wu, K.; Tan, W.W.; Tun, H.W.; Copland, J.A. Stearoyl-coA desaturase 1 is a novel molecular therapeutic target for clear cell renal cell carcinoma. Clin. Cancer Res. 2013, 19, 2368–2380. [Google Scholar] [CrossRef] [PubMed]
- Zu, X.; Zhong, J.; Luo, D.; Tan, J.; Zhang, Q.; Wu, Y.; Liu, J.; Cao, R.; Wen, G.; Cao, D. Chemical genetics of acetyl-coa carboxylases. Molecules 2013, 18, 1704–1719. [Google Scholar] [CrossRef] [PubMed]
- Igal, R.A. Stearoyl coA desaturase-1: New insights into a central regulator of cancer metabolism. Biochim. Biophys. Acta 2016, 1861, 1865–1880. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D. Loss of stearoyl-coA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [PubMed]
- Jaishy, B.; Abel, E.D. Lipids, lysosomes, and autophagy. J. Lipid Res. 2016, 57, 1619–1635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Baehrecke, E.H. Eaten alive: Novel insights into autophagy from multicellular model systems. Trends Cell Biol. 2015, 25, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, T.P.; Baehrecke, E.H. Eating on the fly: Function and regulation of autophagy during cell growth, survival and death in drosophila. Autophagy 2008, 4, 557–562. [Google Scholar] [CrossRef] [PubMed]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Gnerlich, J.L.; Yao, K.A.; Fitchev, P.S.; Goldschmidt, R.A.; Bond, M.C.; Cornwell, M.; Crawford, S.E. Peritumoral expression of adipokines and fatty acids in breast cancer. Ann. Surg. Oncol. 2013, 20 (Suppl. 3), S731–S738. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.M.; Biancur, D.E.; Wang, X.; Halbrook, C.J.; Sherman, M.H.; Zhang, L.; Kremer, D.; Hwang, R.F.; Witkiewicz, A.K.; Ying, H.; et al. Pancreatic stellate cells support tumour metabolism through autophagic alanine secretion. Nature 2016, 536, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Katheder, N.S.; Khezri, R.; O’Farrell, F.; Schultz, S.W.; Jain, A.; Rahman, M.M.; Schink, K.O.; Theodossiou, T.A.; Johansen, T.; Juhasz, G.; et al. Microenvironmental autophagy promotes tumour growth. Nature 2017, 541, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Hart, L.S.; Cunningham, J.T.; Datta, T.; Dey, S.; Tameire, F.; Lehman, S.L.; Qiu, B.; Zhang, H.; Cerniglia, G.; Bi, M.; et al. ER stress-mediated autophagy promotes myc-dependent transformation and tumor growth. J. Clin. Investig. 2012, 122, 4621–4634. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Varga, A.; Pircs, K.; Hegedus, K.; Juhasz, G. Myc-driven overgrowth requires unfolded protein response-mediated induction of autophagy and antioxidant responses in drosophila melanogaster. PLoS Genet. 2013, 9, e1003664. [Google Scholar] [CrossRef] [PubMed]
- Kohler, K.; Brunner, E.; Guan, X.L.; Boucke, K.; Greber, U.F.; Mohanty, S.; Barth, J.M.; Wenk, M.R.; Hafen, E. A combined proteomic and genetic analysis identifies a role for the lipid desaturase Desat1 in starvation-induced autophagy in drosophila. Autophagy 2009, 5, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Holder, A.M.; Gonzalez-Angulo, A.M.; Chen, H.; Akcakanat, A.; Do, K.A.; Fraser Symmans, W.; Pusztai, L.; Hortobagyi, G.N.; Mills, G.B.; Meric-Bernstam, F. High stearoyl-coA desaturase 1 expression is associated with shorter survival in breast cancer patients. Breast Cancer Res. Treat. 2013, 137, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Ide, Y.; Waki, M.; Hayasaka, T.; Nishio, T.; Morita, Y.; Tanaka, H.; Sasaki, T.; Koizumi, K.; Matsunuma, R.; Hosokawa, Y.; et al. Human breast cancer tissues contain abundant phosphatidylcholine(36ratio1) with high stearoyl-coA desaturase-1 expression. PLoS ONE 2013, 8, e61204. [Google Scholar] [CrossRef]
- Huang, J.; Fan, X.X.; He, J.; Pan, H.; Li, R.Z.; Huang, L.; Jiang, Z.; Yao, X.J.; Liu, L.; Leung, E.L.; et al. SCD1 is associated with tumor promotion, late stage and poor survival in lung adenocarcinoma. Oncotarget 2016, 7, 39970–39979. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.Y.; Neufeld, T.P. An ATG1/ATG13 complex with multiple roles in tor-mediated autophagy regulation. Mol. Biol. Cell 2009, 20, 2004–2014. [Google Scholar] [CrossRef] [PubMed]
- Bellosta, P.; Hulf, T.; Balla Diop, S.; Usseglio, F.; Pradel, J.; Aragnol, D.; Gallant, P. Myc interacts genetically with tip48/reptin and tip49/pontin to control growth and proliferation during drosophila development. Proc. Natl. Acad. Sci. USA 2005, 102, 11799–11804. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, L.J.; Gao, X.; Chiarelli, D.A.; Li, L.; Pan, D.; Edgar, B.A. Rheb promotes cell growth as a component of the insulin/TOR signalling network. Nat. Cell Biol. 2003, 5, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Da Huang, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using david bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, P3. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from gene ontology and reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, H.; Miyashita, T.; Nakano, Y.; Yamamoto, D. Hspin1, a transmembrane protein interacting with Bcl-2/Bcl-xL, induces a caspase-independent autophagic cell death. Cell Death Differ. 2003, 10, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, Y.; Gibson, S.B. Starvation-induced autophagy is regulated by mitochondrial reactive oxygen species leading to ampk activation. Cell. Signal. 2013, 25, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Kirkin, V.; McEwan, D.G.; Novak, I.; Dikic, I. A role for ubiquitin in selective autophagy. Mol. Cell 2009, 34, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, G.; Hill, J.H.; Yan, Y.; Sass, M.; Baehrecke, E.H.; Backer, J.M.; Neufeld, T.P. The class III PI(3)K Vps34 promotes autophagy and endocytosis but not tor signaling in drosophila. J. Cell Biol. 2008, 181, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Bellenghi, M.; Puglisi, R.; Pedini, F.; De Feo, A.; Felicetti, F.; Bottero, L.; Sangaletti, S.; Errico, M.C.; Petrini, M.; Gesumundo, C.; et al. SCD5-induced oleic acid production reduces melanoma malignancy by intracellular retention of SPARC and cathepsin B. J. Pathol. 2015, 236, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Tameire, F.; Koumenis, C. Perk-ing up autophagy during myc-induced tumorigenesis. Autophagy 2013, 9, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Johnston, L.A. Socializing with MYC: Cell competition in development and as a model for premalignant cancer. Cold Spring Harb Perspect Med. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Kanamoto, R.; Yokota, T.; Hayashi, S.I. Expressions of c-myc and insulin-like growth factor-1 mRNA in the liver of growing rats vary reciprocally in response to changes in dietary protein. J. Nutr. 1994, 124, 2329–2334. [Google Scholar] [PubMed]
- Teleman, A.A.; Hietakangas, V.; Sayadian, A.C.; Cohen, S.M. Nutritional control of protein biosynthetic capacity by insulin via Myc in drosophila. Cell Metab. 2008, 7, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Toh, P.P.; Luo, S.; Menzies, F.M.; Rasko, T.; Wanker, E.E.; Rubinsztein, D.C. Myc inhibition impairs autophagosome formation. Hum. Mol. Genet. 2013, 22, 5237–5248. [Google Scholar] [CrossRef] [PubMed]
- Shajahan-Haq, A.N.; Cook, K.L.; Schwartz-Roberts, J.L.; Eltayeb, A.E.; Demas, D.M.; Warri, A.M.; Facey, C.O.; Hilakivi-Clarke, L.A.; Clarke, R. Myc regulates the unfolded protein response and glucose and glutamine uptake in endocrine resistant breast cancer. Mol. Cancer 2014, 13, 239. [Google Scholar] [CrossRef] [PubMed]
- Janikiewicz, J.; Hanzelka, K.; Dziewulska, A.; Kozinski, K.; Dobrzyn, P.; Bernas, T.; Dobrzyn, A. Inhibition of SCD1 impairs palmitate-derived autophagy at the step of autophagosome-lysosome fusion in pancreatic beta-cells. J. Lipid Res. 2015, 56, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Theodoropoulos, P.C.; Gonzales, S.S.; Winterton, S.E.; Rodriguez-Navas, C.; McKnight, J.S.; Morlock, L.K.; Hanson, J.M.; Cross, B.; Owen, A.E.; Duan, Y.; et al. Discovery of tumor-specific irreversible inhibitors of stearoyl CoA desaturase. Nat. Chem. Biol. 2016, 12, 218–225. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProtKB ID | Protein Name | Fold Change | p-Value * | Unique Peptides ** |
|---|---|---|---|---|
| A1Z7H2 | Acyl-CoA synthetase long-chain(Acsl) | +2.74 | 6.41 × 10−35 | 7 |
| ACADM | Probable medium-chain specific acyl-CoA dehydrogenase, mitochondrial | +1.71 | 1.48 × 10−6 | 16 |
| MECR | Probable trans-2-enoyl-CoA reductase, mitochondrial | +1.3 | 1.61 × 10−22 | 3 |
| Q9VVU1 | Acyl-CoA dehydrogenase activity CG3902 | +1.4 | 1.21 × 10−19 | 14 |
| Q9VDT1 | Arc42 | +1.44 | 1.24 × 10−36 | 7 |
| Q9VCC6 | fatty acyl-CoA synthase activity CG6178 | +1.71 | 3.71 × 10−28 | 7 |
| Q9W5W8 | isomerase activity CG9577 | +1.34 | 7.82 × 10−14 | 10 |
| Q9VXI1 | 3-hydroxyacyl-CoA dehydrogenase activity, CG9914 | +1.5 | 9.76 × 10−25 | 15 |
| CYB5R | Cytochrome b5-related(Cyt-b5-r) | +1.31 | 2.47 × 10−14 | 4 |
| Q7K4Y0 | Desaturase 1(Desat1) | +2.64 | 8.86 × 10−43 | 6 |
| Q9BLE1 | Desaturase 2(Desat2) | +1.3 | 1.40 × 10−3 | 2 |
| Q7JWF1 | Electron transfer flavoprotein-ubiquinone oxidoreductase(Etf-QO) | +1.64 | 0 | 9 |
| GPDA | Glycerol 3 phosphate dehydrogenase(Gpdh) | +1.52 | 1.81 × 10−39 | 23 |
| B7Z0E0 | Isocitrate dehydrogenase(Idh) | +1.49 | 0 | 9 |
| LSD1 | Lipid storage droplet-1(Lsd-1) | +1.51 | 9.85 × 10−13 | 11 |
| Q0KHS7 | Lipid storage droplet-2(Lsd-2) | +2.77 | 1.87 × 10−25 | 2 |
| Q961S9 | Microsomal triacylglycerol transfer protein(Mtp) | +1.48 | 2.45 × 10−5 | 8 |
| Q8IPE8 | Mitochondrial trifunctional protein alpha subunit(Mtpalpha) | +1.37 | 1.60 × 10−14 | 22 |
| Q9VL70 | Yippee interacting protein 2(yip2) | +1.3 | 6.13 × 10−64 | 19 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paiardi, C.; Mirzoyan, Z.; Zola, S.; Parisi, F.; Vingiani, A.; Pasini, M.E.; Bellosta, P. The Stearoyl-CoA Desaturase-1 (Desat1) in Drosophila cooperated with Myc to Induce Autophagy and Growth, a Potential New Link to Tumor Survival. Genes 2017, 8, 131. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8050131
Paiardi C, Mirzoyan Z, Zola S, Parisi F, Vingiani A, Pasini ME, Bellosta P. The Stearoyl-CoA Desaturase-1 (Desat1) in Drosophila cooperated with Myc to Induce Autophagy and Growth, a Potential New Link to Tumor Survival. Genes. 2017; 8(5):131. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8050131
Chicago/Turabian StylePaiardi, Chiara, Zhasmine Mirzoyan, Sheri Zola, Federica Parisi, Andrea Vingiani, Maria Enrica Pasini, and Paola Bellosta. 2017. "The Stearoyl-CoA Desaturase-1 (Desat1) in Drosophila cooperated with Myc to Induce Autophagy and Growth, a Potential New Link to Tumor Survival" Genes 8, no. 5: 131. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8050131