Nuclear Integrity but Not Topology of Mouse Sperm Chromosome is Affected by Oxidative DNA Damage

 and
and
Abstract
:1. Introduction
2. Results
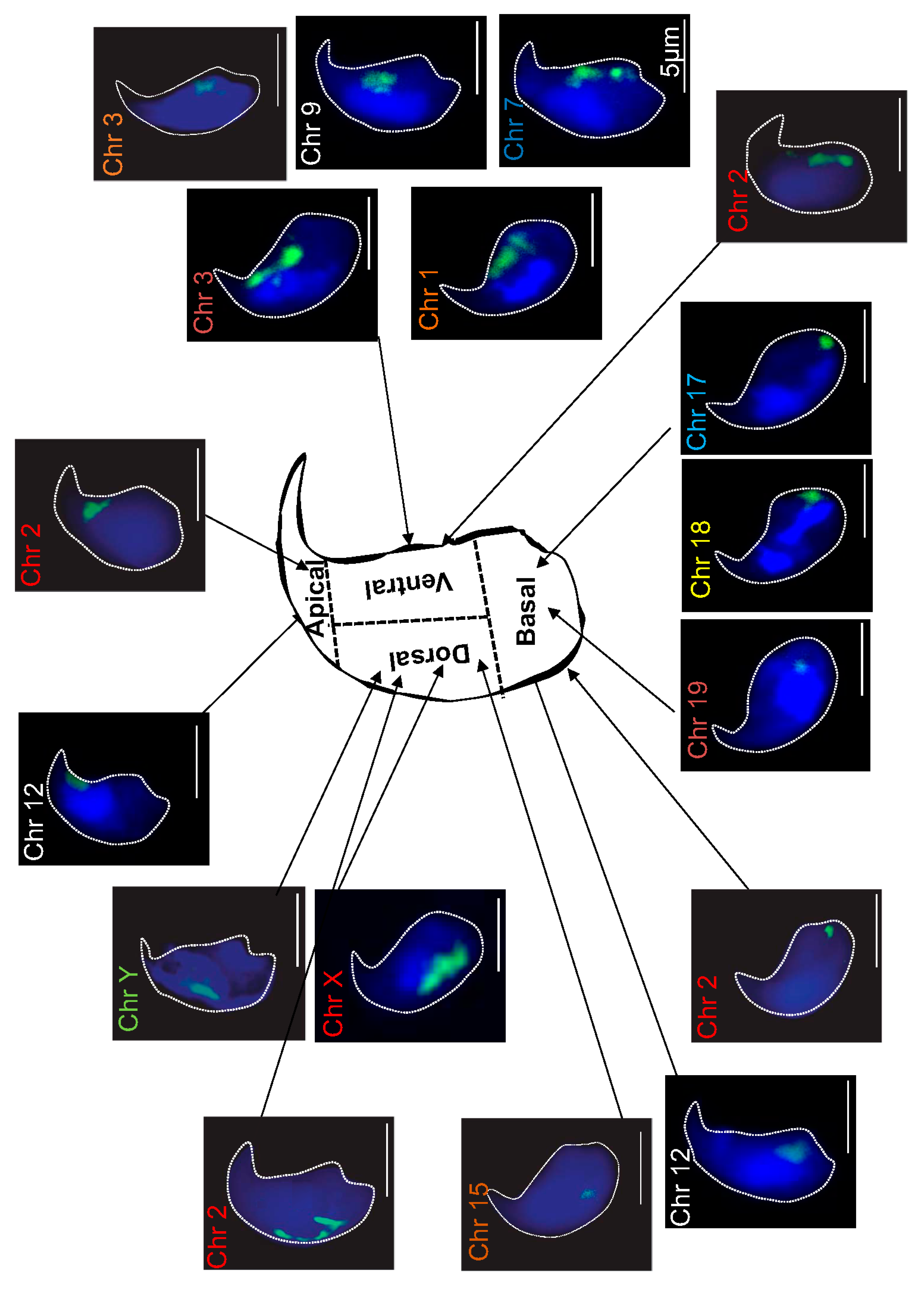
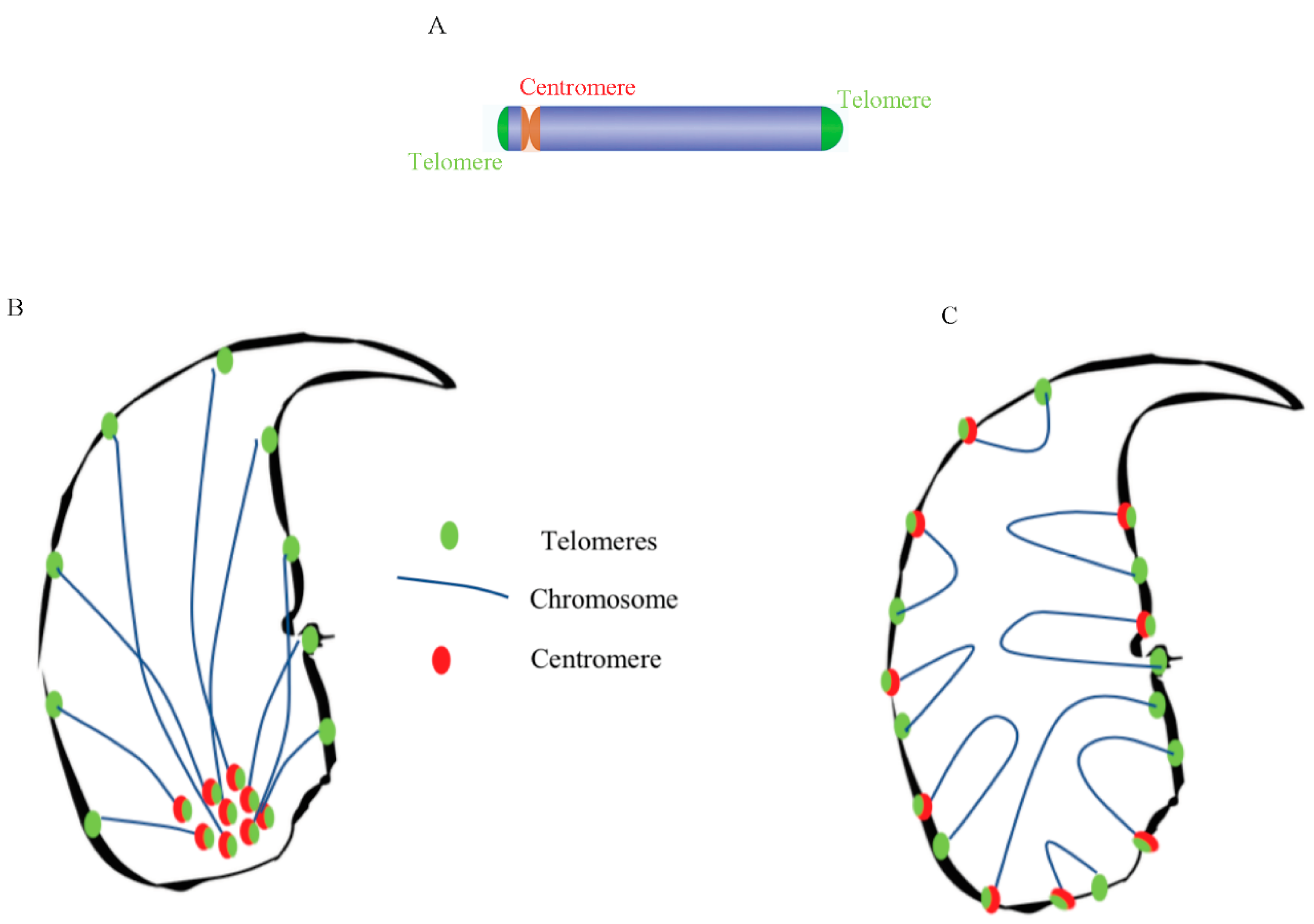
2.1. Localization of Chromosome Territories in Murine Spermatozoa
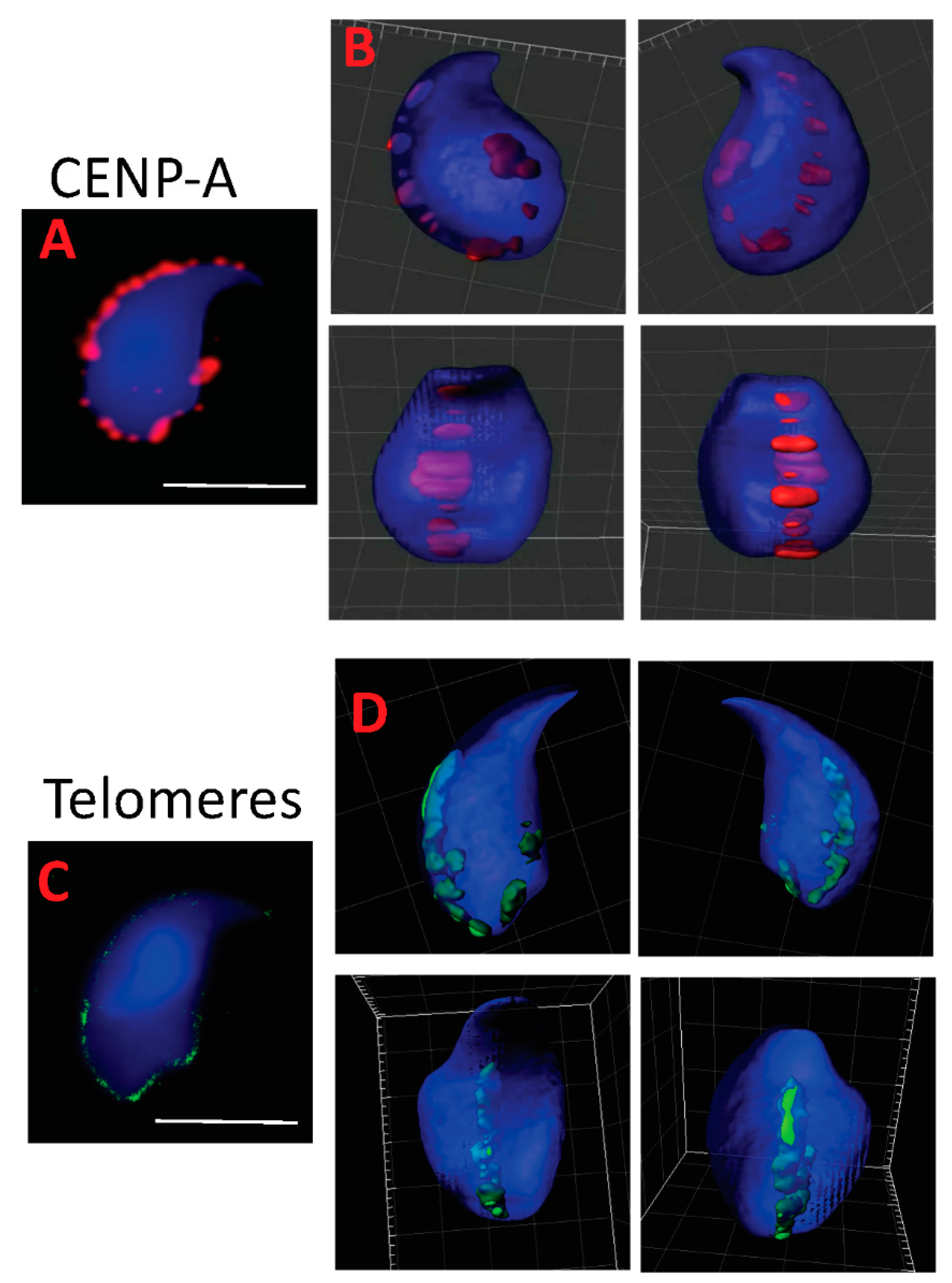
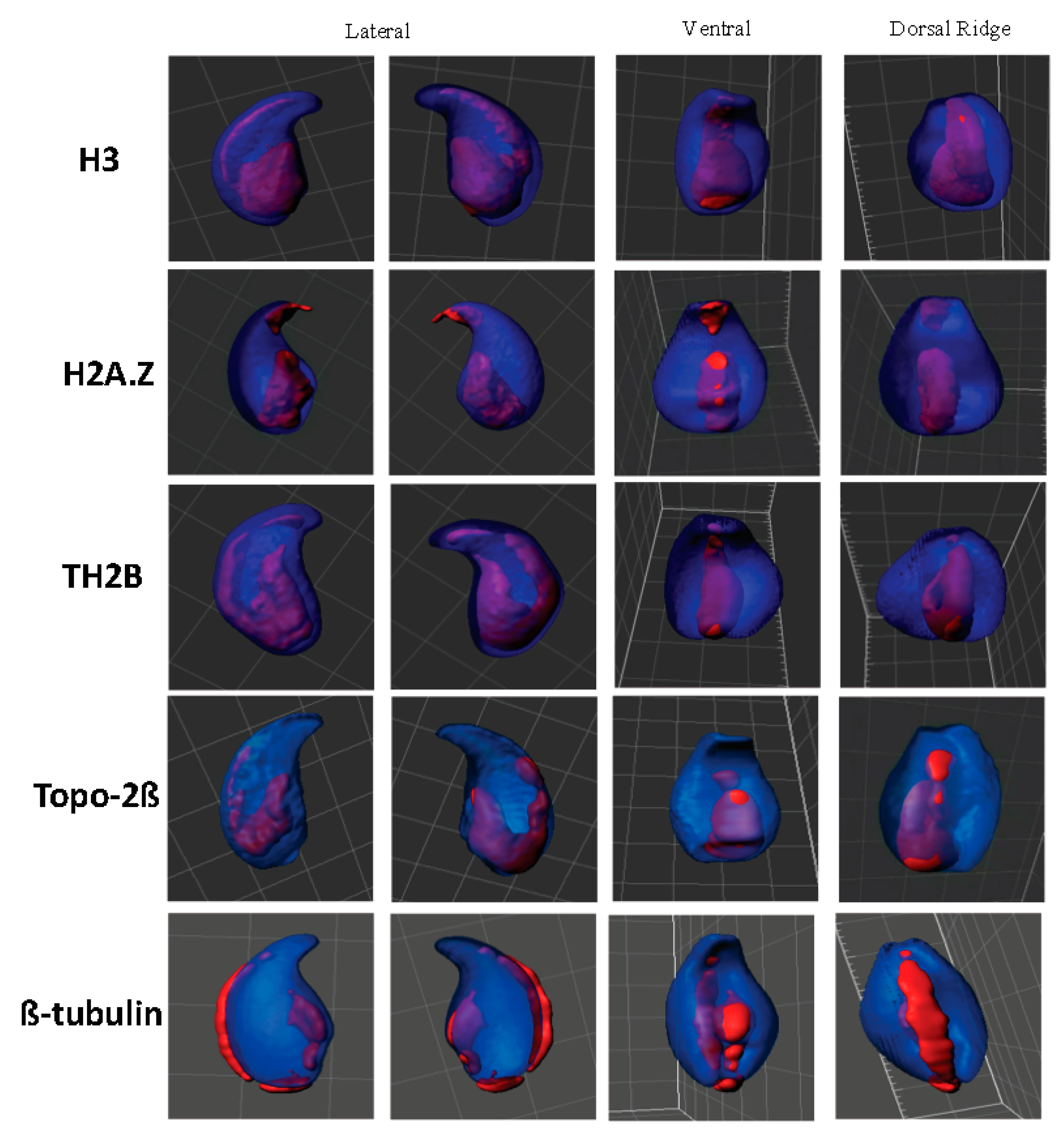
2.2. Centromeres, Telomeres, and Histone-Rich Domains Clustered in the Mouse Sperm Nucleus
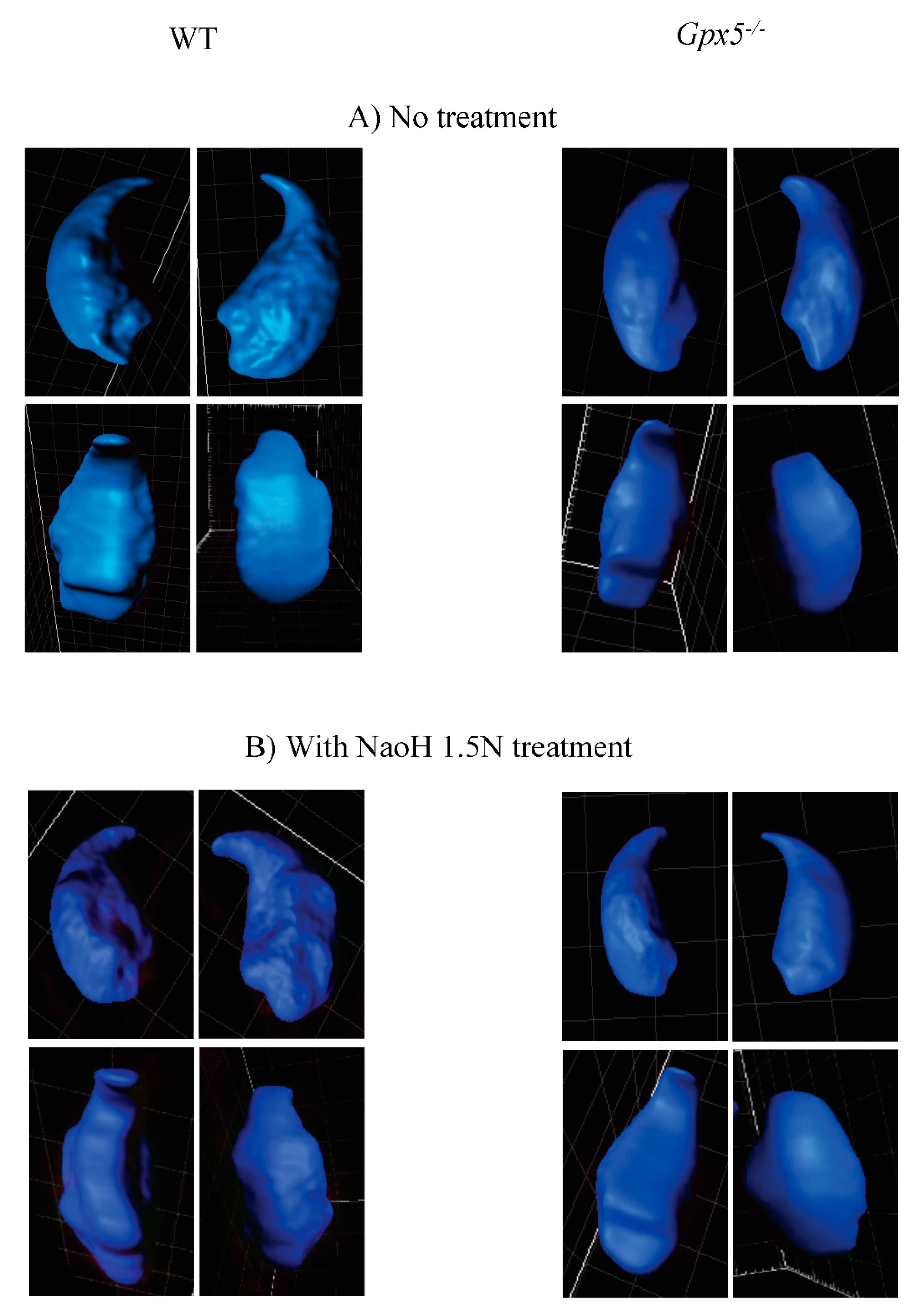
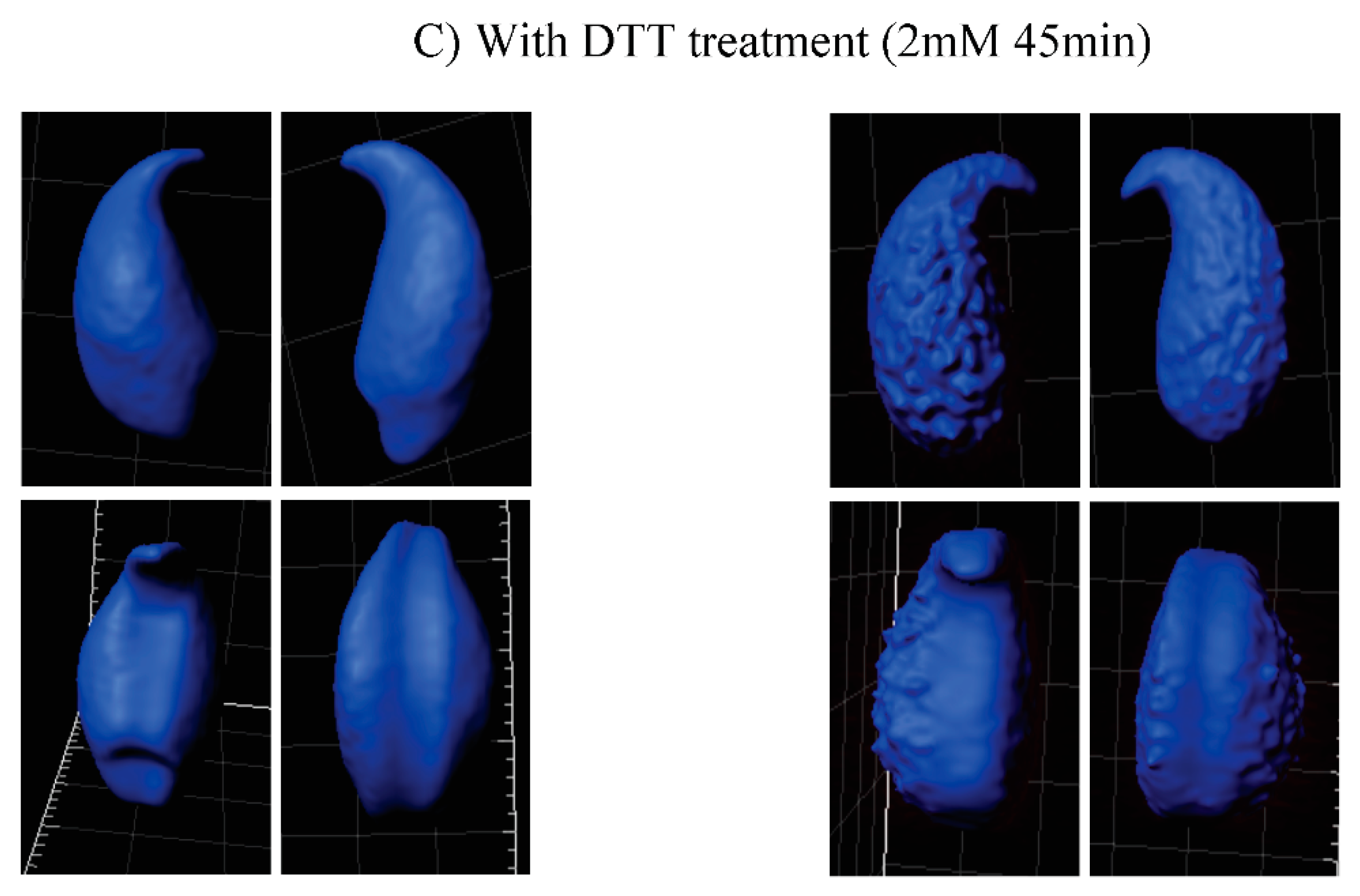
2.3. Oxidative DNA Damage Does Affect 3D-Parameters of the Mouse Sperm Nucleus
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals
4.3. Immunocytochemistry and Fluorescence in situ Hybridization (FISH) Assays
4.4. Microscopy
4.5. Image Analysis Measurements of 3D Parameters
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Champroux, A.; Torres-Carreira, J.; Gharagozloo, P.; Drevet, J.R.; Kocer, A. Mammalian sperm nuclear organization: Resiliencies and vulnerabilities. Basic Clin. Androl. 2016, 26, 17. [Google Scholar] [CrossRef] [PubMed]
- Montellier, E.; Boussouar, F.; Rousseaux, S.; Zhang, K.; Buchou, T.; Fenaille, F.; Shiota, H.; Debernardi, A.; Héry, P.; Curtet, S.; et al. Chromatin-to-nucleoprotamine transition is controlled by the histone H2B variant TH2B. Genes Dev. 2013, 27, 1680–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Romero, R.; Méndez, J.; Ausió, J.; Eirín-López, J.M. Quickly evolving histones, nucleosome stability and chromatin folding: All about histone H2A.Bbd. Gene 2008, 413, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Govin, J.; Escoffier, E.; Rousseaux, S.; Kuhn, L.; Ferro, M.; Thévenon, J.; Catena, R.; Davidson, I.; Garin, J.; Khochbin, S.; et al. Pericentric heterochromatin reprogramming by new histone variants during mouse spermiogenesis. J. Cell Biol. 2007, 176, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoghoughi, N.; Barral, S.; Vargas, A.; Rousseaux, S.; Khochbin, S. Histone variants: Essential actors in the male genome programing. J. Biochem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Balhorn, R. The protamine family of sperm nuclear proteins. Genome Biol. 2007, 8, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathke, C.; Baarends, W.M.; Awe, S.; Renkawitz-Pohl, R. Chromatin dynamics during spermiogenesis. Biochim. Biophys. Acta 2014, 1839, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Bradbury, E.M.; Balhorn, R. AFM analysis of DNA-protamine complexes bound to mica. Nucleic Acids Res. 1997, 25, 2221–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, L.; Corzett, M.; Lau, E.Y.; Balhorn, R. Dynamics of protamine 1 binding to single DNA molecules. J. Biol. Chem. 2003, 278, 42403–42408. [Google Scholar] [CrossRef] [PubMed]
- Hud, N.V.; Allen, M.J.; Downing, K.H.; Lee, J.; Balhorn, R. Identification of the elemental packing unit of DNA in mammalian sperm cells by atomic force microscopy. Biochem. Biophys. Res. Commun. 1993, 193, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Hud, N.V.; Downing, K.H.; Balhorn, R. A constant radius of curvature model for the organization of DNA in toroidal condensates. Proc. Natl. Acad. Sci. USA 1995, 92, 3581–3585. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.S.; Coffey, D.S. DNA packaging and organization in mammalian spermatozoa: Comparison with somatic cells. Biol. Reprod. 1991, 44, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Champroux, A.; Cocquet, J.; Henry-Berger, J.; Drevet, J.R.; Kocer, A. A Decade of Exploring the Mammalian Sperm Epigenome: Paternal Epigenetic and Transgenerational Inheritance. Front. Cell Dev. Biol. 2018, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Foster, H.A.; Abeydeera, L.R.; Griffin, D.K.; Bridger, J.M. Non-random chromosome positioning in mammalian sperm nuclei, with migration of the sex chromosomes during late spermatogenesis. J. Cell Sci. 2005, 118, 1811–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, N.M.; Lau, P.; Hann, M.; Ioannou, D.; Hoffman, D.; Barrionuevo, M.; Maxson, W.; Ory, S.; Tempest, H.G. Hierarchical radial and polar organisation of chromosomes in human sperm. Chromosom. Res. Int. J. Mol. Supramol. Evol. Asp. Chromosom. Biol. 2012, 20, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Zalensky, A.; Zalenskaya, I. Organization of chromosomes in spermatozoa: An additional layer of epigenetic information? Biochem. Soc. Trans. 2007, 35, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Hazzouri, M.; Rousseaux, S.; Mongelard, F.; Usson, Y.; Pelletier, R.; Faure, A.K.; Vourc’h, C.; Sèle, B. Genome organization in the human sperm nucleus studied by FISH and confocal microscopy. Mol. Reprod. Dev. 2000, 55, 307–315. [Google Scholar] [CrossRef]
- Zalensky, A.O.; Breneman, J.W.; Zalenskaya, I.A.; Brinkley, B.R.; Bradbury, E.M. Organization of centromeres in the decondensed nuclei of mature human sperm. Chromosoma 1993, 102, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Shaman, J.A.; Yamauchi, Y.; Ward, W.S. Function of the sperm nuclear matrix. Arch. Androl. 2007, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.S. Function of sperm chromatin structural elements in fertilization and development. Mol. Hum. Reprod. 2010, 16, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Gawecka, J.E.; Ribas-Maynou, J.; Benet, J.; Ward, W.S. A model for the control of DNA integrity by the sperm nuclear matrix. Asian J. Androl. 2015, 17, 610–615. [Google Scholar] [PubMed]
- Yamauchi, Y.; Shaman, J.A.; Ward, W.S. Non-genetic contributions of the sperm nucleus to embryonic development. Asian J. Androl. 2011, 13, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.; Gilchrist, S.; Bridger, J.M.; Mahy, N.L.; Ellis, J.A.; Bickmore, W.A. The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells. Hum. Mol. Genet. 2001, 10, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, J.A.; Bridger, J.M.; Boyle, S.; Perry, P.; Teague, P.; Bickmore, W.A. Differences in the localization and morphology of chromosomes in the human nucleus. J. Cell Biol. 1999, 145, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Bolzer, A.; Kreth, G.; Solovei, I.; Koehler, D.; Saracoglu, K.; Fauth, C.; Müller, S.; Eils, R.; Cremer, C.; Speicher, M.R.; et al. Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 2005, 3, e157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.B.; Shen, J.; Yokota, H. Size-dependent positioning of human chromosomes in interphase nuclei. Biophys. J. 2000, 79, 184–190. [Google Scholar] [CrossRef]
- Bench, G.S.; Friz, A.M.; Corzett, M.H.; Morse, D.H.; Balhorn, R. DNA and total protamine masses in individual sperm from fertile mammalian subjects. Cytometry 1996, 23, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Balhorn, R.; Gledhill, B.L.; Wyrobek, A.J. Mouse sperm chromatin proteins: Quantitative isolation and partial characterization. Biochemistry 1977, 16, 4074–4080. [Google Scholar] [CrossRef] [PubMed]
- Tovich, P.R.; Oko, R.J. Somatic histones are components of the perinuclear theca in bovine spermatozoa. J. Biol. Chem. 2003, 278, 32431–32438. [Google Scholar] [CrossRef] [PubMed]
- Arpanahi, A.; Brinkworth, M.; Iles, D.; Krawetz, S.A.; Paradowska, A.; Platts, A.E.; Saida, M.; Steger, K.; Tedder, P.; Miller, D. Endonuclease-sensitive regions of human spermatozoal chromatin are highly enriched in promoter and CTCF binding sequences. Genome Res. 2009, 19, 1338–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkek, S.; Hisano, M.; Liang, C.; Gill, M.; Murr, R.; Dieker, J.; Schübeler, D.; van der Vlag, J.; Stadler, M.B.; Peters, A.H.F.M. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nat. Struct. Mol. Biol. 2013, 20, 868–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammoud, S.S.; Nix, D.A.; Zhang, H.; Purwar, J.; Carrell, D.T.; Cairns, B.R. Distinctive chromatin in human sperm packages genes for embryo development. Nature 2009, 460, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Gatewood, J.M.; Cook, G.R.; Balhorn, R.; Schmid, C.W.; Bradbury, E.M. Isolation of four core histones from human sperm chromatin representing a minor subset of somatic histones. J. Biol. Chem. 1990, 265, 20662–20666. [Google Scholar] [PubMed]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Noblanc, A.; Damon-Soubeyrand, C.; Karrich, B.; Henry-Berger, J.; Cadet, R.; Saez, F.; Guiton, R.; Janny, L.; Pons-Rejraji, H.; Alvarez, J.G.; et al. DNA oxidative damage in mammalian spermatozoa: Where and why is the male nucleus affected? Free Radic. Biol. Med. 2013, 65, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Zalensky, A.O.; Siino, J.S.; Gineitis, A.A.; Zalenskaya, I.A.; Tomilin, N.V.; Yau, P.; Bradbury, E.M. Human testis/sperm-specific histone H2B (hTSH2B). Molecular cloning and characterization. J. Biol. Chem. 2002, 277, 43474–43480. [Google Scholar] [CrossRef] [PubMed]
- Shaman, J.A.; Yamauchi, Y.; Ward, W.S. The sperm nuclear matrix is required for paternal DNA replication. J. Cell. Biochem. 2007, 102, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Anachkova, B.; Djeliova, V.; Russev, G. Nuclear matrix support of DNA replication. J. Cell. Biochem. 2005, 96, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solov’eva, L.; Svetlova, M.; Bodinski, D.; Zalensky, A.O. Nature of telomere dimers and chromosome looping in human spermatozoa. Chromosom. Res. Int. J. Mol. Supramol. Evol. Asp. Chromosom. Biol. 2004, 12, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, W.S.; Coffey, D.S. Identification of a sperm nuclear annulus: A sperm DNA anchor. Biol. Reprod. 1989, 41, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Kocer, A.; Henry-Berger, J.; Noblanc, A.; Champroux, A.; Pogorelcnik, R.; Guiton, R.; Janny, L.; Pons-Rejraji, H.; Saez, F.; Johnson, G.D.; et al. Oxidative DNA damage in mouse sperm chromosomes: Size matters. Free Radic. Biol. Med. 2015, 89, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Chabory, E.; Damon, C.; Lenoir, A.; Kauselmann, G.; Kern, H.; Zevnik, B.; Garrel, C.; Saez, F.; Cadet, R.; Henry-Berger, J.; et al. Epididymis seleno-independent glutathione peroxidase 5 maintains sperm DNA integrity in mice. J. Clin. Investig. 2009, 119, 2074–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannou, D.; Miller, D.; Griffin, D.K.; Tempest, H.G. Impact of sperm DNA chromatin in the clinic. J. Assist. Reprod. Genet. 2016, 33, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Mudrak, O.S.; Nazarov, I.B.; Jones, E.L.; Zalensky, A.O. Positioning of chromosomes in human spermatozoa is determined by ordered centromere arrangement. PLoS ONE 2012, 7, e52944. [Google Scholar] [CrossRef] [PubMed]
- Zalenskaya, I.A.; Zalensky, A.O. Non-random positioning of chromosomes in human sperm nuclei. Chromosom. Res. Int. J. Mol. Supramol. Evol. Asp. Chromosom. Biol. 2004, 12, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Zalenskaya, I.A.; Bradbury, E.M.; Zalensky, A.O. Chromatin structure of telomere domain in human sperm. Biochem. Biophys. Res. Commun. 2000, 279, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Y.; Baker, H.W.G. Human sperm bound to the zona pellucida have normal nuclear chromatin as assessed by acridine orange fluorescence. Hum. Reprod. 2007, 22, 1597–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannou, D.; Griffin, D.K. Male fertility, chromosome abnormalities, and nuclear organization. Cytogenet. Genome Res. 2011, 133, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Haaf, T.; Ward, D.C. Higher order nuclear structure in mammalian sperm revealed by in situ hybridization and extended chromatin fibers. Exp. Cell Res. 1995, 219, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Jennings, C.; Powell, D. Genome organisation in the murine sperm nucleus. Zygote Camb. Engl. 1995, 3, 123–131. [Google Scholar] [CrossRef]
- Luetjens, C.M.; Payne, C.; Schatten, G. Non-random chromosome positioning in human sperm and sex chromosome anomalies following intracytoplasmic sperm injection. Lancet Lond. Engl. 1999, 353, 1240. [Google Scholar] [CrossRef]
- Meyer-Ficca, M.; Müller-Navia, J.; Scherthan, H. Clustering of pericentromeres initiates in step 9 of spermiogenesis of the rat (Rattus norvegicus) and contributes to a well defined genome architecture in the sperm nucleus. J. Cell Sci. 1998, 111 Pt 10, 1363–1370. [Google Scholar]
- Powell, D.; Cran, D.G.; Jennings, C.; Jones, R. Spatial organization of repetitive DNA sequences in the bovine sperm nucleus. J. Cell Sci. 1990, 97 Pt 1, 185–191. [Google Scholar]
- Sbracia, M.; Baldi, M.; Cao, D.; Sandrelli, A.; Chiandetti, A.; Poverini, R.; Aragona, C. Preferential location of sex chromosomes, their aneuploidy in human sperm, and their role in determining sex chromosome aneuploidy in embryos after ICSI. Hum. Reprod. Oxf. Engl. 2002, 17, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Tilgen, N.; Guttenbach, M.; Schmid, M. Heterochromatin is not an adequate explanation for close proximity of interphase chromosomes 1--Y, 9--Y, and 16--Y in human spermatozoa. Exp. Cell Res. 2001, 265, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Zalensky, A.O.; Tomilin, N.V.; Zalenskaya, I.A.; Teplitz, R.L.; Bradbury, E.M. Telomere-telomere interactions and candidate telomere binding protein(s) in mammalian sperm cells. Exp. Cell Res. 1997, 232, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Greaves, I.K.; Rens, W.; Ferguson-Smith, M.A.; Griffin, D.; Marshall Graves, J.A. Conservation of chromosome arrangement and position of the X in mammalian sperm suggests functional significance. Chromosom. Res. Int. J. Mol. Supramol. Evol. Asp. Chromosom. Biol. 2003, 11, 503–512. [Google Scholar] [CrossRef]
- Watson, J.M.; Meyne, J.; Graves, J.A. Ordered tandem arrangement of chromosomes in the sperm heads of monotreme mammals. Proc. Natl. Acad. Sci. USA 1996, 93, 10200–10205. [Google Scholar] [CrossRef] [PubMed]
- Joffe, B.I.; Solovei, I.V.; Macgregor, H.C. Ordered arrangement and rearrangement of chromosomes during spermatogenesis in two species of planarians (Plathelminthes). Chromosoma 1998, 107, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Solovei, I.V.; Joffe, B.I.; Hori, T.; Thomson, P.; Mizuno, S.; Macgregor, H.C. Unordered arrangement of chromosomes in the nuclei of chicken spermatozoa. Chromosoma 1998, 107, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lalancette, C.; Miller, D.; Krawetz, S.A. Characterization of nucleohistone and nucleoprotamine components in the mature human sperm nucleus. Asian J. Androl. 2008, 10, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.E.; Krawetz, S.A. Computer assisted promoter analysis of a human sperm specific nucleoprotein gene cluster. DNA Seq. J. DNA Seq. Mapp. 1995, 5, 329–337. [Google Scholar] [CrossRef]
- Thilagavathi, J.; Venkatesh, S.; Dada, R. Telomere length in reproduction. Andrologia 2013, 45, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, D.; Millan, N.M.; Jordan, E.; Tempest, H.G. A new model of sperm nuclear architecture following assessment of the organization of centromeres and telomeres in three-dimensions. Sci. Rep. 2017, 7, 41585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotolongo, B.; Huang, T.T.F.; Isenberger, E.; Ward, W.S. An endogenous nuclease in hamster, mouse, and human spermatozoa cleaves DNA into loop-sized fragments. J. Androl. 2005, 26, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Wykes, S.M.; Krawetz, S.A. The structural organization of sperm chromatin. J. Biol. Chem. 2003, 278, 29471–29477. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, D.; Tempest, H.G. Does genome organization matter in spermatozoa? A refined hypothesis to awaken the silent vessel. Syst. Biol. Reprod. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wiland, E.; Fraczek, M.; Olszewska, M.; Kurpisz, M. Topology of chromosome centromeres in human sperm nuclei with high levels of DNA damage. Sci. Rep. 2016, 6, 31614. [Google Scholar] [CrossRef] [PubMed]
- Noblanc, A.; Peltier, M.; Damon-Soubeyrand, C.; Kerchkove, N.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Janny, L.; Pons-Rejraji, H.; et al. Epididymis response partly compensates for spermatozoa oxidative defects in snGPx4 and GPx5 double mutant mice. PLoS ONE 2012, 7, e38565. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | Gpx5−/− | |||||||
|---|---|---|---|---|---|---|---|---|
| Basal | Apical | Ventral | Dorsal | Basal | Apical | Ventral | Dorsal | |
| Chr 1 | 27.9 | 3.4 | 49.7 | 19 | 29.4 | 8.2 | 46.3 | 16.1 |
| Chr 2 | 25.1 | 20.2 | 28.4 | 26.3 | 25.5 | 21 | 27.5 | 26 |
| Chr 3 | 35.9 | 13.8 | 31.8 | 18.5 | N.D. | |||
| Chr 7 | 32.5 | 9.8 | 40.3 | 17.4 | 29 | 7.5 | 47.5 | 16 |
| Chr 9 | 29.5 | 9.8 | 49 | 11.7 | 30 | 3.8 | 44.6 | 21.6 |
| Chr 12 | 36.8 | 32.9 | 13.1 | 17.2 | 34.2 | 27.1 | 18.9 | 19.8 |
| Chr 15 | 21.8 | 2.8 | 22.8 | 52.6 | 19 | 7 | 24 | 50 |
| Chr 17 | 57.2 | 14.1 | 13.2 | 15.5 | 53.8 | 15.8 | 16.2 | 14.2 |
| Chr 18 | 58.2 | 22.4 | 11.2 | 8.2 | 57.2 | 24.3 | 11.1 | 7.4 |
| Chr 19 | 67.2 | 17 | 13.6 | 2.2 | 61.5 | 18.4 | 13.4 | 6.7 |
| Chr X | 7.7 | 20.7 | 7.5 | 64.1 | 5.3 | 30.3 | 4.1 | 60.3 |
| Chr Y | 3.8 | 29.9 | 7.2 | 59.1 | 4.5 | 25.4 | 5.3 | 64.8 |
| WT | Gpx5−/− | |
| Average volume (μm3) | 66 | 54.8 a |
| Average Area (μm2) | 93.9 | 80.2 a |
| Nucleus with NaOH 1.5N Treatment | WT | Gpx5−/− |
| Average volume (μm3) | 109 d | 93.5 b,d |
| Average Area (μm2) | 138 d | 113.5 b,d |
| Nucleus with DTT Treatment (2 mM, 45 min) | WT | Gpx5−/− |
| Average volume (μm3) | 85.3 d | 130.4 c,d |
| Average Area (μm2) | 108.4 d | 143.3 c,d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Champroux, A.; Damon-Soubeyrand, C.; Goubely, C.; Bravard, S.; Henry-Berger, J.; Guiton, R.; Saez, F.; Drevet, J.; Kocer, A. Nuclear Integrity but Not Topology of Mouse Sperm Chromosome is Affected by Oxidative DNA Damage. Genes 2018, 9, 501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9100501
Champroux A, Damon-Soubeyrand C, Goubely C, Bravard S, Henry-Berger J, Guiton R, Saez F, Drevet J, Kocer A. Nuclear Integrity but Not Topology of Mouse Sperm Chromosome is Affected by Oxidative DNA Damage. Genes. 2018; 9(10):501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9100501
Chicago/Turabian StyleChamproux, Alexandre, Christelle Damon-Soubeyrand, Chantal Goubely, Stephanie Bravard, Joelle Henry-Berger, Rachel Guiton, Fabrice Saez, Joel Drevet, and Ayhan Kocer. 2018. "Nuclear Integrity but Not Topology of Mouse Sperm Chromosome is Affected by Oxidative DNA Damage" Genes 9, no. 10: 501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9100501