Telomere Maintenance Mechanisms in Cancer
by
 , , , , and
, , , , and
Tiago Bordeira Gaspar
1,2,3,4,† ,
,
Ana Sá
1,2,4,† ,
,
José Manuel Lopes
1,2,3,5 ,
,
Manuel Sobrinho-Simões
1,2,3,5,
Paula Soares
1,2,4,* and
João Vinagre
1,2,3 1
Cancer Signaling and Metabolism Group, Institute for Research and Innovation in Health Sciences (i3S), University of Porto, 4200-135 Porto, Portugal
2
Cancer Signaling and Metabolism Group, Institute of Molecular Pathology and Immunology of the University of Porto (Ipatimup), 4200-135 Porto, Portugal
3
Medical Faculty of University of Porto (FMUP), 4200-139 Porto, Portugal
4
Abel Salazar Biomedical Sciences Institute (ICBAS), University of Porto, 4050-313 Porto, Portugal
5
Department of Pathology and Oncology, Centro Hospitalar São João, 4200-139 Porto, Portugal
*
Author to whom correspondence should be addressed.
†
These authors contributed equally.
Genes 2018, 9(5), 241; https://0-doi-org.brum.beds.ac.uk/10.3390/genes9050241
Submission received: 14 March 2018
/
Revised: 20 April 2018
/
Accepted: 23 April 2018
/
Published: 3 May 2018
(This article belongs to the Section Human Genomics and Genetic Diseases)
Abstract
:Tumour cells can adopt telomere maintenance mechanisms (TMMs) to avoid telomere shortening, an inevitable process due to successive cell divisions. In most tumour cells, telomere length (TL) is maintained by reactivation of telomerase, while a small part acquires immortality through the telomerase-independent alternative lengthening of telomeres (ALT) mechanism. In the last years, a great amount of data was generated, and different TMMs were reported and explained in detail, benefiting from genome-scale studies of major importance. In this review, we address seven different TMMs in tumour cells: mutations of the TERT promoter (TERTp), amplification of the genes TERT and TERC, polymorphic variants of the TERT gene and of its promoter, rearrangements of the TERT gene, epigenetic changes, ALT, and non-defined TMM (NDTMM). We gathered information from over fifty thousand patients reported in 288 papers in the last years. This wide data collection enabled us to portray, by organ/system and histotypes, the prevalence of TERTp mutations, TERT and TERC amplifications, and ALT in human tumours. Based on this information, we discuss the putative future clinical impact of the aforementioned mechanisms on the malignant transformation process in different setups, and provide insights for screening, prognosis, and patient management stratification.
1. Introduction
Gradual accumulation of genetic errors in cells is a major contributor to the tumourigenic process. In the transition to a malignant tumour (i.e., cancer), an acquired immortality state is mandatory, and tumour cells must cope with selective pressure. It is therefore required that cancer cells gain advantages against tumour suppressive mechanisms. Limiting telomere shortening is one of those mechanisms, being the topic of this review.
Telomeres are DNA–protein complexes at the ends of eukaryotic chromosomes that play a crucial role in cellular survival, by limiting progressive loss of genomic information caused by semi-conservative replication of DNA [1,2]. Most cancer cells maintain the integrity of their telomeres by telomerase reactivation (TR) [3], and the mechanisms accounting for telomere length (TL) maintenance are currently known to comprise transcriptional, post-transcriptional, and epigenetic regulation [4,5]. A small part of tumour cells acquires immortality through the alternative lengthening of telomeres (ALT) mechanism. An understanding of these mechanisms and respective age- and tumour-related changes will hopefully unveil novel biomarkers and targets with diagnostic and prognostic impact, and ultimately influence the development of novel therapeutics [3,6].
In this review, we address seven telomere maintenance mechanisms (TMMs) in tumour cells, including genetic (promoter mutations, amplifications, germline genetic variations, rearrangements) and epigenetic (DNA methylation and non-coding RNAs) events.
2. Telomere Maintenance Mechanisms
2.1. Telomere Maintenance Mechanisms in Non-Malignant Cells
Telomeres are specialized ribonucleoprotein structures composed of DNA and bound proteins localized at the ends of all linear chromosomes [7,8]. Telomeric DNA contains a multiple short non-coding tandem repeat of double-stranded DNA sequence, 5′-(TTAGGG)n-3′ that is 10–15 kilobases (kb) long in humans at birth, and a 3′ G-rich single-stranded tail of 150–200 nucleotides [9,10].
The proteins associated with telomeres comprise the shelterin complex that promotes telomere protection, by ensuring stability and assisting specialized replication machinery for accurate extension of chromosome ends [7,10,11] and recruitment of telomerase [8,12,13]. The shelterin complex consists of six subunits, three DNA-binding (TRF1, TRF2, POT1) interconnected by three additional proteins (TIN2, TPP1, RAP1) that act as adaptors and mediate interactions among the constituents [14,15].
Telomeres play vital roles in dealing with two unavoidable biological challenges, the end protection—by safeguarding chromosomes from being recognized as double stranded free DNA breaks by the DNA damage response (DDR) machinery, that may result in end-fusions and genome instability [12]—and the end replication crises—the inherent limitation of DNA replication, i.e., the gradual shortening of DNA at chromosomal ends at each replicative cycle [8,10].
Telomerase is a complex ribonucleic reverse transcriptase responsible for synthetizing telomeric DNA repeats at the 3′ ends of linear chromosomes [9,15,16]. It comprises the catalytic protein subunit telomerase reverse transcriptase (TERT), encoded by the TERT gene (located at 5p15.33), an essential RNA component (TERC) that functions as the RNA template for the addition of telomeric repeats, encoded by the TERC gene (at chromosome 3q26) [3,4,17], and a series of auxiliary components with important biologic functions that include dyskerin, reptin, pontin, and ribonucleoproteins NOP10, GAR1, and NHP2 [15,18,19]. TERC, additionally to its role in the template for the synthesis of telomere DNA, is also involved in the catalysis, localization, and assembly of telomerase [20].
Defects in these telomerase players are known to cause telomere deficiency syndromes or telomeropathies, as reviewed by some authors [9,21,22].
Telomere length in stem cells is established with a relative size that grants tissue homeostasis during embryogenesis but is short-limited enough to suppress unlimited cell proliferation and tumour initiation [23]. As proliferating cells of self-renewing tissues depend on telomerase activity as a pivotal TMM, most human somatic tissues do not express sufficient telomerase to infinitely sustain TL, leading to gradual telomere shortening [24,25]. Therefore, cells undergo gradual age-related telomere shortening, at a variable rate per mitosis [9,26]. Gradual telomere attrition reflects one of the hallmarks of aging [27].
As reviewed by Jafri et al. [4], telomerase is responsible for a multistep process required for telomere maintenance, that includes TERT protein transport and trafficking into the nucleus, TERC and TERT assembly with accessory components in the nucleus, and recruitment to the telomeres at the correct timing during DNA replication. Repressors and enhancers within TERT promoter engage in a transcriptional suppression of the catalytic subunit in most somatic cells, thus limiting telomerase activity [15,28].
Telomere length is also regulated by epigenetic marking in telomeric chromatin [29,30]. The compacted chromatin state of mammals, which contains histone modifications suggestive of a constitutive heterochromatin, negatively regulates TL [31]. When these heterochromatic marks are lost, telomere elongation occurs, as reported in mouse cells, suggesting that a compacted chromatin state at telomeres is fundamental for controlling TL; i.e., the compaction of chromatin and subsequent difficult access of transcription factors may induce negative regulation of TMMs [30,31,32].
As telomeres shorten, they can also modify at a transcriptionally level the expression of nearby genes, telomere position effect (TPE), or over long distant genes (TPE-OLD) [33]. Telomere position effect involves the spreading of telomeric heterochromatin to silence genes in the vicinity of telomeres according to TL, while TPE-OLD telomeres fold back and physically interact with other chromosome domains, producing widespread changes in gene regulation much sooner than TL decreases above a critical level to induce DDR [33].
Independent of the reactivation of telomerase, ALT represents a TMM based on homologous recombination (HR) and homology-directed telomere synthesis [34,35] that was thought to be exclusive of tumour cells; still, it has been identified in stem cells and healthy tissues of mouse [31,36]. It has also been detected in human cells of the placenta in early gestation [37] and endothelial, stromal, and some epithelial cells of non-neoplastic cells [38]. This mechanism might thus occur naturally in another physiological setting that is not fully understood at this point and can be a recombination-based mechanism. Finally, telomere sequences contain long non-coding RNAs—telomeric repeat-containing RNA (TERRA)—with important functions on telomere homeostasis and telomerase function [39], that will be further addressed.
2.2. Telomere Maintenance Mechanisms in Tumour Cells
The ability to keep telomeres above a critical length represents a vital feature of malignant cells [40]. Activation of a TMM, dependent or independent of the enzyme telomerase, allows tumour cells to survive cellular crisis and achieve immortality, one of the major hallmarks of cancer [41,42,43].
Both TERT and TERC codify limiting protein components of telomerase activity [44]. Transcription, alternative messenger RNA (mRNA) splicing, phosphorylation, maturation, and modification of TERT and of TERC have been reported to play vital roles in the regulation of telomerase activity [3].
Concurring with tumour heterogeneity, TL is also expected to fluctuate [45]. It was reported that genes closer to telomeres display higher expression in tumours than in normal tissues, due to the reduced TL of the first; and this effect seems gradually attenuated as distance to telomeres increases [45].
The central role of TMMs in cancer led to the development of several therapeutic strategies aiming at inhibiting telomerase and/or telomere function, such as the use of small-molecule telomerase inhibitors, oligonucleotide inhibitors, immunotherapy, and G-quadruplex stabilizers [46,47,48].
Telomere maintenance in tumour cells is ensured by TR in over 85% [45,49,50,51,52,53,54] of human tumours, while ALT mechanism occurs in 10–15% [35,55,56].
The most characterized mechanisms and alterations (Figure 1) responsible for maintaining the lengthening of telomeres in tumour cells are: (1) somatic mutations of the TERT promoter (TERTp); (2) amplification of the genes TERT and TERC (3) rearrangements of the TERT gene; (4) germline genetic variants of the TERT gene and its promoter; (5) epigenetic changes; (6) ALT; and (7) non-defined TMM (NDTMM).
The recent study by Barthel et al. [45] highlighted the telomere length and frequencies of telomere maintenance by mechanism and tumour type in The Cancer Genome Atlas (TCGA) cohort. By analysing the data from 288 papers, we collected the percentages of occurrence of five different TMMs (TERTp mutations, TERT and TERC amplifications, TERT rearrangements, and ALT) from over fifty thousand cases.
The different TMMs are extremely diverse amongst several tumours in different locations and histotypes. When considering large cohorts (more than 100 patients) the tumours with the highest prevalence of TERTp mutations are glioblastoma (GB) IDH-wildtype (72%), oligodendroglioma (OD) IDH-mutant and 1p19q-codeleted (95%), anaplastic oligodendroglioma (AOD) (63%), adult sonic hedgehog medulloblastoma (SHH-MB, 89%), hepatocellular carcinoma (HCC, 41%), oral squamous cell carcinoma (SCC) (50%), basal cell carcinoma (BCC) and SCC of the skin (49% and 56% respectively), metastatic cutaneous melanoma (76%), urothelial bladder carcinoma, both non-muscle invasive (NMIBC, 69%), and muscle invasive (MIBC, 68%), that are in contrast to tumours with high cell turnover that present less prevalence of TERTp mutations, e.g., tumours of the digestive system (0–2%) and haematopoietic and lymphoid tissues (0%).
Adenocarcinoma and SCC of the lung (18% and 40%, respectively) contrast with oral SCC (2%) concerning the presence of TERT amplifications in cohorts with more than 200 patients. Cervical intraepithelial neoplasia (CIN) presents a prevalence of TERC amplifications that increases with progression: 24%, 69% and 88% (CIN1, 2, and 3, respectively). Lung SCC was also reported with a high frequency of TERC amplifications (41%).
High-risk and high-stage neuroblastoma (NBL) (15%) is, so far, the best characterized tumour model for rearrangements of the TERT gene (cohort of 292 patients).
ALT mechanism also exhibits lower rates in tumours of the digestive and haematopoietic systems, while neuroblastomas (50%) and osteosarcomas (63%) frequently display this phenotype (cohorts with more than 100 patients). A large cohort of patients with pancreatic neuroendocrine tumours (pNETs) were reported to display 30% of ALT positivity. Sporadic pNETs often present ALT, whereas TERTp mutations are detected in a fraction of hereditary pNETs [57].
These data portray the diverse panoply by which TMMs can be found in human tumours. For the sake of simplicity, we will address each tumour histotype, whenever available, according to the current World Health Organization (WHO) classification, for the several organ/tissue locations. When discrete histotypes were not available or the reported cohorts included few cases, we included them in a not otherwise specified (NOS) group, indicating tumour histotypes as reported by the authors (Table 1, Table 2, Table 3 and Table 4). Molecular associations, prognostic, and clinical implications of TMMs in human tumours are summarized in Table 5. Information regarding the distribution of absent/low frequency TMMs in prevalent tumours can be consulted in Table 6.
2.2.1. TERT Promoter Mutations
Somatic mutations in the coding region of TERT are not frequent in tumours. Somatic mutations in the TERTp, however, have recently emerged as the most prevalent non-coding mutations in human cancer.
TERTp is a 260 bp region that lacks a TATA box or other similar sequence, containing binding motifs for various transcription factors that regulate gene transcription and are responsible for TERT’s transcriptional activity and telomerase activation [3,4].
Independent somatic mutations in the core promoter of TERT were recently reported in melanoma [189,190]. The most frequent mutations resulted in a C>T transition at −124 bp and −146 bp upstream the ATG transcriptional start site; these hotspot mutations are also known as C228T and C250T, respectively, based on their genomic coordinates. Both create an 11 bp nucleotide stretch that contains a consensus binding site for E-twenty-six (ETS) transcription factors within the promoter region. This provides a basis for the biological relevance of these mutations, since ETS transcription factors may be activated through dysregulation of mitogen-activated protein kinase (MAP kinase) signalling, commonly observed with increased gene expression in some tumours [189,190,343]. Tumours with TERTp mutations were consistently reported to express higher levels of TERT mRNA and telomerase activity when compared to those with a wildtype promoter [51,75,189,190,204,344,345].
The level and frequency of these mutations varies greatly between tumour histotypes [3]. After their first report in melanoma, TERTp mutations have been reported in many other types of tumours, such as tumours of the central nervous system (CNS), thyroid, skin, and liver [49,51,70,96,116,172,202,344,346]. This neoplastic spectrum led to the postulation that TERTp mutations preferentially trigger tumorigenesis in tissues with relatively low rates of self-renewal, that are not able to overcome the short-telomere dependent proliferative barrier, and arise as a late tumorigenic event [4,49]. Still, TERTp mutations can also appear as an early tumorigenic event, resulting from environmental factors, namely, ultraviolet radiation or chemical carcinogens. This possibility is supported by their high prevalence in BCC, melanoma, and urothelial bladder carcinoma [22].
The frequencies of TERTp mutations were reviewed by tumour histotype as depicted in Table 1. Regarding diffuse astrocytic and oligodendroglial tumours, gliosarcomas present the highest mutation rate (81%), followed by glioblastomas IDH-wildtype (72%). The prevalence of TERTp mutations varies considerably between primary/secondary (68% vs. 25%) and adult NOS/paediatric GBs (63% vs. 6%). IDH-mutant and 1p19q-codeleted anaplastic oligodendrogliomas present a lower prevalence TERTp mutations (21%) when comparing to NOS histotypes (63%). Anaplastic oligoastrocytomas (AOA, WHO grade III) also have a high mutational rate (41%). Pilocytic astrocytomas (WHO grade I) exhibit low or absent TERTp mutations. Within embryonal tumours, TERTp mutations occur in SHH adult medulloblastomas (89%) and in paediatric SHH-MB (34%). These incidences suggest that TERTp mutations correlate with patient age, as consistently described in studies of CNS tumours, mainly GB [49,50,66,74,98,347]. TERTp mutations seem to be consistently associated with WHO high-grade in CNS tumours. Additionally, it seems that TERTp mutations influence the clinical outcome, since they have been reported to be linked with tumour progression and to predict a worse survival in patients with CNS tumours [65,80,98,346,348].
The association of TERTp mutations with other genetic alterations in CNS tumours needs to be clarified. Isocitrate dehydrogenase (IDH) mutations are common in diffuse astrocytic and oligodendroglial tumours. The combined analysis of different histotypes regarding IDH and TERTp mutational status led to the conclusion that TERTp mutations are more prevalent in IDH-wildtype tumours, such as in GBs (72% in IDH-wildtype vs. 24% in IDH-mutated tumours) and anaplastic astrocytomas (AA) (47% vs. 11%); diffuse astrocytomas present a smaller variation (26% vs. 16%). Remarkably, in ODs IDH-mutated and 1p19q-codeleted, TERTp mutations are associated in a much higher extent to WHO grade II, OD (95%) than in WHO grade III, AOD (21%).
Throughout recent years, several authors have proposed the combination of TERTp mutations with other CNS tumour-associated molecular features, namely IDH mutations, 1p19q loss, and O6-methylguanine-DNA-methyltransferase (O6-MGMT) methylation status, willing to establish a molecular signature that could assist more efficiently in defining an accurate prognosis and the best therapeutic option in patients with diffuse astrocytic and oligodendroglial tumours. The combination of TERTp and IDH mutations, in particular, allows the assignment of diffuse astrocytic and oligodendroglial tumours in discrete groups with different survival rates: TERTp and IDH-mutated tumours present the longest overall survival, while patients with only TERTp-mutated tumours present the lowest survival, as shown in several reports [61,71,73,74,78,79]. Triple positive ODs (TERTp-, IDH-mutated, and 1p19q-codeleted) are associated with better overall survival [71].
In tumours of the digestive system, the highest mutation rates occur in hepatocellular carcinoma (41%) and gallbladder carcinoma (9%). TERTp mutations are absent in hepatocellular adenoma (HCA, 0%); noteworthy, borderline HCA/HCC and HCC derived from HCA present high mutational rates, 48% and 17%, respectively, which suggests the potential involvement of TERTp mutations in the malignant transformation of HCAs [95,96,349].
In some studies, TERTp mutations represent the most common genetic event in HCC [49,96], but other reports present a lower prevalence, which can be due to the studied populations since the mutation frequency varies according to the geographic region [350] and is linked with the prevalence of hepatitis B or C viral infection. TERTp mutations were reported in HCC associated with both viruses, with a higher prevalence for the hepatitis C virus [99,106,351]. Other evaluated tumours of the digestive system presented extremely low or absent prevalence of TERTp mutations, e.g., gastric carcinoma, tumours of the colon and rectum and exocrine pancreas, pointing to a minor role of TERTp mutations in these tumours.
TERTp mutations are highly prevalent in thyroid tumours. The highest mutational rate occurs in aggressive thyroid carcinoma histotypes: anaplastic thyroid carcinoma (ATC, 46%) and poorly differentiated thyroid carcinoma (PDTC, 41%), followed by metastases of well-differentiated papillary and follicular carcinomas (21%), follicular thyroid carcinoma (FTC, 18%), and papillary thyroid carcinoma, NOS (PTC, 11%). Specific subtypes of PTC, such as tall cell variant (TCVPTC), show a higher frequency (19%), consistent with the more aggressive behaviour of this variant of PTC. PTC with TERTp mutations consistently associate with larger tumours, older patient age, higher tumour stage, tumour recurrence, and distant metastases [51,116,119,125,138,333]. TERTp mutations associate with BRAFV600E mutations [117,126,141,146,352]. TERTp mutations are not detected in normal thyroid tissue and the studies in benign tissues and benign thyroid tumours (3%) point to a low frequency; however, the prevalence rises in settings of atypical follicular adenomas (17%) [115] and X-ray irradiation (21%) [139].
Papillary carcinoma-derived anaplastic carcinomas were recently characterized by the co-occurrence of BRAFV600E and TERTp mutations prior to anaplastic transformation. This led the authors to suggest that PTC harbouring TERTp mutations have higher risk of anaplastic transformation [142]. Paediatric thyroid tumours do not seem to harbour these mutations [150,151], which parallels the findings regarding specific subtypes of CNS tumours. At variance with follicular cell derived tumours of the thyroid, medullary thyroid carcinomas do not harbour TERTp mutations.
Within the female reproductive organs, TERTp mutations occur in clear cell carcinoma (CCC) of the uterus and the ovary (21% and 16%, respectively). In ovarian CCC, the mutations were correlated with patient age [153], but not with disease-specific survival [152]. The mutational frequency for uterine endometrial carcinoma, low-grade serous carcinoma of the ovary, and cervical SCC is 11%, 5% and 15%, respectively.
Concerning haematopoietic and lymphoid tissues tumours, TERTp mutations were, so far, detected only in a study of mantle cell lymphomas (22%) [161].
With regard to skin tumours, BCC harbours 49% of TERTp mutations with a UV-signature, indicating UV exposure as a potential cause in these frequent tumours [170,172], being the same signature shared by SCC that harbours 56% TERTp mutations [154]. In cutaneous melanomas, TERTp mutations were reported in several histotypes: superficial spreading (34%), nodular (55%), lentigo maligna (24%), acral lentiginous (9%), and desmoplastic (45%). Metastatic cutaneous melanoma presents a remarkable rate of mutations (76%), as well as metastatic melanoma of unknown primary sites (49%) and of other primary locations (54%). In cutaneous melanomas, the mutations associate with male gender [184,185], older age, tumours with ulceration, and higher Breslow’s thickness [92,175,179,184,221]. In some studies [182,184], the TERTp mutations were not associated with clinical outcome, at variance with the results of Griewank et al. [92] and Pópulo et al. [171], who reported an association between TERTp mutations and poorer overall survival in patients with cutaneous melanomas. The occurrence of TERTp mutations in cutaneous melanoma associates with BRAF mutations and with poorer prognostic features, such as higher Breslow’s thickness, higher mitotic rate, presence of ulceration, absence of regression, and lymph node metastases [92,171,174,190,221]. The BRAF and TERTp mutations may cooperate in cutaneous melanoma and recent evidence indicates that their combination can be used to identify tumours with aggressive behaviour [175,179,186]. Atypical fibroxanthoma and pleomorphic dermal sarcoma are genetically poorly understood and were reported to exhibit a high frequency of TERTp mutations (93% and 77%, respectively), which stands as the most frequent genetic event in such tumours [191]. No mutations were reported either in cutaneous or conjunctival naevi, consistent with a putative late pathogenic role of TERTp mutations in the progression of these melanocytic tumours [90].
TERTp mutations are highly frequent in urinary system tumours. They are the most frequent alterations in both invasive (68%) and non-muscle invasive (69%) urothelial cell carcinoma of the bladder. Their presence is of limited prognostic value due to the equal mutational rate in different stages or grades [208] but may be a useful biomarker for urine-based tumour monitoring (non-invasive diagnostic tool in two settings: early detection in high-risk patients and recurrence in patients with urothelial bladder carcinoma [203,206,209]. High telomerase activity has been correlated with urothelial carcinomas as a better marker of disease aggressiveness than TERTp mutations alone [344], but when combined with FGFR3 mutations, these mutations assist on establishing tumours of poor prognosis [222]. In kidney tumours, TERTp mutations are less prevalent, with a frequency of 9% in clear cell renal cell carcinoma (ccRCC, 9%). The mutations in ccRCC are less prevalent than in many other tumour types, but their presence was reported to characterize a subset of tumours that demand more aggressive treatment [198]. TERTp mutations were not reported, so far, in prostate cancer [49,168,201,219,353].
Chiba et al. [354] recently proposed the contribution of TERTp mutations to tumorigenesis in a two-step mechanism. The authors reported that in an initial phase, TERTp mutations heal the shortest telomeres and extend life-span; in a second phase, genome instability arises as a consequence of critically short telomeres, inducing the upregulation of telomerase to sustain cell proliferation [354]. The selection of TERTp mutations at the transition from pre-neoplastic to malignancy suggests that telomere shortening acts as a critical barrier early in the tumorigenesis of some cancers.
TERTp mutations currently stand as highly prevalent events in a spectrum of tumours, with varying rates according to histotype (Table 1). They present a relevant role in hepatocarcinogenesis and also stand as a powerful tool with impact on clinical management, namely screening of patients with urothelial carcinoma of the bladder, or prognosis stratification of patients with diffuse astrocytic and oligodendroglial, thyroid, and skin tumours.
2.2.2. TERT and TERC Amplifications
Copy number variations (CNVs), or alterations (CNAs), affect a larger fraction of the genome in cancers than do any other type of somatic genetic alteration [355,356,357,358]. CNVs are more successfully detected by next generation sequencing [45], rather than by fluorescence in situ hybridization (FISH) studies used in the past [359].
Gain or loss of genetic material is commonly observed in human malignancies [17,52,360]. DNA amplification is a frequent event in the tumorigenic process and typically culminates in gene overexpression [17,52,235,238,361].
Some authors reported CNVs of whole chromosomes or chromosome arms in a large number of human tumours. In the year 1999, Rooney et al. [360] identified the chromosome arms 3q (16.4%) and 5p (13.2%), respectively, as the sixth and eighth regions where more frequently chromosomal gains occurs, by comparative genomic hybridization (CGH) in 2210 solid tumours of 27 tumour types. As reviewed by Soder et al. [238], Knuutila et al. [362], and Sugita et al. [235], amplifications in 3q have been associated to many tumour types, including ovarian carcinoma [363,364,365], cervical carcinoma [366,367], lung carcinoma (both small cell [368,369,370,371] and non-small cell [365,372]), SCC of the head and neck (SCC-HN) [372,373,374], and non-Hodgkin lymphoma [375]. Amplifications in 5p have also been detected in cervical carcinoma [367], non-small cell lung cancer (NSCLC) [376], SCC-HN [374], and in many other tumours, as osteosarcoma [377], malignant fibrous histiocytoma of soft tissue [378], and gastrointestinal stromal tumour (GIST) [379].
Considering that TERT has been mapped to chromosome 5 at 5p15.33 and TERC has been mapped to chromosome 3 at 3q26.3 [238,359], authors started to search for specific changes in copy number. Both amplifications were reported to be genetic alterations that induce strong TERT [52,163,229,380] and TERC [238,359] overexpression in some tumours, although the specificity of these amplifications remains to be established [238]. Noteworthy, HCC [226] and malignant pleural mesothelioma [163] are tumours in which TERT gene is overexpressed but not 9ssociated with gene copy number gain.
Increased TERT and TERC gene dosage has been detected frequently in a variety of human tumours, and clonal evolution of cells with increased TERT or TERC copy number has been observed, pointing towards a growth advantage in cells with increased TERT or TERC gene dosage [17]. Below, we present the prevalence of TERT and TERC amplifications in several tumour types. For histotype information, report to Table 2.
In cutaneous melanomas (Table 2), an increase of CNVs comes with tumour progression, as TERT amplifications were detected only in invasive melanomas, whereas they were rarely detected in benign naevi, and occasionally present in intermediate lesions, melanomas in situ, and desmoplastic melanomas. TERT amplifications were reported in 23% of acral-lentiginous melanomas and less than 5% in desmoplastic melanomas.
Despite the already stated role of TERTp mutations in hepatocarcinogenesis as an early event in tumour progression and the cause of higher TERT expression [99,103], TERT amplification does not show a clear correlation with progression. In HCC, TERTp mutations and TERT focal amplifications are almost mutually exclusive [99]. Overall, amplifications of the TERT gene were detected in 15% of HCC.
In addition to HCC, TERT amplifications presented a substantial prevalence in lung adenocarcinoma (18%) and lung SCC (25%), colorectal carcinoma (48%), and cervical intraepithelial neoplasm (CIN) 2 and 3 (60% and 88%, respectively).
Besides desmoplastic cutaneous melanomas, TERT amplifications presented the lowest levels in phyllodes tumour of the breast (4%) and oral SCC (2%).
TERT amplifications have also important utility in diagnosis of a variety of solid tumours, including breast (differentiating phyllodes tumours from fibroadenomas) [223], NSCLC [381] and urothelial bladder carcinomas [234]. Also, TERT amplifications may represent a poor prognostic marker in breast [382] and urothelial bladder carcinomas [234], NSCLC [229], and acral-lentiginous melanoma [231].
Amplifications of the TERC gene (Table 2) were reported at high levels in 21% of oesophageal carcinoma [45], 41% of lung SCC [45], ovarian (37%), and cervical tumours (59%) [45,235]. Andersson et al. [240] wrote the first report of consistent gain of TERC in cervical adenocarcinoma. In what concerns CINs, both TERC and TERT amplifications can be accurately detected with FISH technique in routinely collected liquid based cytology (LBC) by Pap smears [227,236,237].
Gains of TERC gene significantly associate with a gradual increasing amplification pattern in tumour progression of ovarian malignancies [227,236,237,383,384]. Visnovsky et al. [227] reported for TERT gene a very similar amplification pattern that also associates with histopathological and cytopathological findings. Liu et al. [236] reported a significant positive correlation between the level of TERC amplification and histologic grade of intraepithelial cervical lesions: lower in low-grade (CIN 1) than in high-grade (CIN 2/3). Both authors describe TERC amplification in all cases of malignant cervical carcinomas evaluated.
Concerning urothelial bladder carcinomas, amplification of the TERT gene appear to be useful in discriminating patients with non-muscle (0%) and muscle invasive (56%) tumours in the study of Yamamoto et al. [234].
In conclusion, TERC and TERT amplification assessment may be useful in the future as a complement to the HPV test and help to establish the risk of malignancy of cervical precursor lesions, aiming the highest combined sensitivity and specificity [227,236,237], i.e., early identification of patients with low-grade lesions and higher progression risk [384].
Copy number variations represent an additional mechanism for telomere maintenance based on the capacity to modulate telomerase overexpression and for some tumour types it was reported a mutual exclusivity towards other TMMs.
2.2.3. TERT Germline Genetic Variations
The 5p15.33 TERT-CLPTM1L chromosomal region has been consistently associated with susceptibility for multiple tumours [385]. Genome-wide association studies (GWAS) performed in large scale in the last decade have strongly contributed to the identification of common variants in TERT locus. Several single nucleotide polymorphisms (SNPs) in this region have been consistently associated with increased risk for developing various types of tumours. These SNPs may arise either in intronic or exonic sequences of TERT or in its promoter. The more described genetic variants for both regions of TERT and their associations with several types of cancers are depicted in Table 3; most studies are genome wide associations where the results are not available by histotypes.
The polymorphism rs2736100 is localized in intron 2 of TERT and it is one of the most described variants of the gene. It has been thoroughly associated with lung cancer risk (mostly adenocarcinoma [242,249]), although there were also reports of no evident association [386]. Wang et al. [387] and Yang et al. [388] published meta-analyses in which they reported the association of rs2736100 with increased lung cancer risk (mainly adenocarcinoma) in both Caucasian and Asian populations. This SNP was identified to associate with an increased risk of myeloproliferative neoplasms (e.g., polycythaemia vera, essential thrombocythemia, and primary myelofibrosis) in Caucasian and Chinese populations [267,269]. Conflicting results have been reported regarding the effect of this SNP on gastric cancer risk, in which rs2736100 was associated with increased risk in a Turkish population [265] but did not show impact on an Asian population, in which it correlated only with the regulation of TERT expression and telomere length [389]. On the other hand, rs2736100 was significantly associated with reduced risk of upper tract urothelial carcinoma [390] and increased prostate cancer aggressiveness [391]. No association was reported for this SNP and colorectal cancer [274] or breast cancer risk [276]. Given the conflicting and heterogeneous results obtained from various studies, several meta-analyses have been published to address the effects of the polymorphism on cancer risk. Zou et al. [392] reported a significant association between rs2736100 and cancer susceptibility, with strong associations for lung and pancreatic cancers and BCC, and also risk alleles for bladder, prostate and cervical cancers, as well as gliomas (including WHO grades II–IV astrocytic, and WHO grades II–III oligodendroglial tumours). Li et al. [393] reported that rs2736100 polymorphism in heterozygous and homozygous variant genetic settings affected cancer susceptibility from a gathering of 16 case-control studies. The evaluated studies reported discrepancies that could be explained by different allele frequencies in different ethnicities. Peng et al. [335] evaluated the association between rs2736100 and the risk of glioma (including WHO grades II–IV astrocytic and WHO grades II–III oligodendroglial tumours) from 16 independent studies and reported that this genetic variation may greatly enhance susceptibility for developing these types of tumours, with consistent results obtained for Caucasians. The authors emphasized the need of more studies for Asian populations and pointed to the fact that the heterogeneity found could be attributed to genetic backgrounds, living environments and patients’ characteristics.
The rs2736098 is a synonymous coding SNP in the second exon of the TERT gene that was associated with telomere length but not TERT expression [271]. Rafnar et al. [271] reported its association with increased risk of BCC and lung, prostate and bladder cancers. This variant has also been reported not to associate with breast cancer risk [288] and to reduce the risk of SCC-HN and oral cavity in Caucasian patients [266,394]. Wu et al. [395] demonstrated that this polymorphism may contribute to the risk of lung cancer (especially adenocarcinoma), but they found it to be only weakly associated with overall cancer risk.
The rs2853676 maps to intron two of TERT [243]. Other types of cancer than the ones detailed in Table 3 (breast, gastric, lung, prostate and ovary; gliomas; and melanomas) have been investigated regarding this SNP, and no associations were found for endometrial cancer [396], BCC or SCC of the skin [279], colorectal cancer [397,398], breast cancer [399] or paediatric acute lymphoblastic leukaemia [256]. Cao et al. [336] performed a systematic review and meta-analysis to ascertain the impact of rs2853676 on cancer risk, and they reported association with increased risk of glioma (including WHO grades II-IV astrocytic and grades II-III oligodendroglial tumours), lung and ovarian cancers among Caucasian populations.
The rs2853669 functional variant is located in the TERTp within a binding site for ETS2 transcription factor [385]. It was reported to affect telomerase activity and telomere length [275,400]. Rs2853669 appears to be associated with prognosis, affecting survival and tumour recurrence in urothelial bladder carcinoma, although these results are conflicting among different studies [204,222]. Depending on the rs2853669 SNP status, glioblastoma patients carrying TERTp mutations displayed worse prognosis and shorter survival [68,76,80]. This polymorphism has also been consistently associated with cutaneous melanoma patients carrying TERTp mutations [188,293], and its use to identify patients at risk of aggressive disease was proposed [175]. Beyond association with increased cancer risk and cancer prognosis per se, rs2853669 has been reported to have a modifying effect on TERTp mutations [65,76,80,175,198,204,292,401], and an eventual prognostic value [402].
TERTp polymorphism rs2735940 acts similarly but is reported in less extent than the rs2853669. The association with cancer risk has been found for lung and gastric cancers [265,294], as well as for paediatric acute lymphoblastic leukaemia [256]. No association between this polymorphism and colorectal cancer or colonic polyps has been found [397], although these results are conflicting among studies [403].
TERT polymorphisms are being addressed as factors with impact on the risk of developing several cancers, with increasing evidence for tumours of the CNS and lung. However, there are still conflicting reports, which renders this a debatable issue. It should be kept in mind that a different genetic background and/or racial and ethnic disparities may play important roles in the pathogenesis and modulate disease incidence. TERTp polymorphisms, their reported modifying effect on TERTp mutations, and their use in patient prognosis are also an interesting target to be further explored.
2.2.4. TERT Rearrangements
Chromosomal rearrangements are another TMM that occur in human tumours. Genomic rearrangements can result in tandem duplications, inverted orientations, interchromosomal changes, amplification, and deletions [404]. The TERT gene can also be a target of this alteration. Unlike other TMMs, rearrangements have only been extensively explored, to our knowledge, in a single tumour subset: neuroblastoma (NBL). NBL is a malignant embryonic tumour that arises from the peripheral sympathetic nerve system and represents the most common extracranial solid tumour in children associated with unfavourable patient outcome [339,340,405].
Amplification of the proto-oncogene MYCN has been used for many years in these patients as a reliable marker for defining high-risk disease [406], but only recently, recurrent genomic rearrangements proximal of the TERT gene have been reported in NBL, defining a subgroup of high-risk tumours with a particular poorer outcome [54,340]. At the present time, both TERT rearrangements [54,339,340] and MYCN amplification [339,340,341] are two well-established indicators of poor prognosis. The most aggressive form of the disease appears to be related with high telomerase activity [339,340,407], which is caused by both alterations [54,408,409,410]. Some studies reported that structural rearrangements affecting the chromosomal region at 5p15.33 lead to juxtaposition of strong enhancer elements in close proximity to the TERT locus [45,54,339,341], resulting in a massive transcriptional upregulation of TERT and adjacent genes distal of the breakpoints, and a strong epigenetic remodelling of the affected region (histone modifications and DNA methylation) [54,339,341]. Epigenetic marks were reported as absent in NBLs without these rearrangements [54]. These rearrangements occurred only in high-risk NBLs [54,339] in a mutually exclusive fashion with MYCN amplifications and ATRX mutations [54] that are common genetic alteration in NBLs [340]. TERT rearrangements are structurally diverse [54], as translocations are both inter and intrachromosomal [405]. Low-risk NBLs lack evidence of active TMMs and high-risk NBLs without TERT or MYCN alterations lack telomerase activity and are characterized by activation of the ALT pathway [339].
Moreover, intratumoural diversity in TL is another feature in NBL [341]. The diversity of TL in individual NBLs was strongly associated with disease progression and death [317,411,412]. On the basis of these studies, Jeison et al. [412] defined two subtypes of NBLs with poor clinical outcome: the first comprising cases with MYCN amplification, typically demonstrating decreased or unaltered TL, and the second comprising cases presenting normal MYCN status and increased TL. When combining high-risk and high-stage NBLs, TERT rearrangements account for 18% of the cases, in contrast to the 13% observed in low to high-stages NBLs [54,340,405].
TERT rearrangements in NBLs represent the second most frequent genetic defect following MYCN alterations. In NBLs, the TERT rearrangements were almost mutually exclusive with MYCN and ATRX (associated to another TMM, ALT), stratifying them in two groups of NBLs with very high risk, reinforcing the concept that tumours do not present multiple TMM simultaneously.
2.2.5. Epigenetic Mechanisms
Epigenetic alterations consist of alterations that affect cell behaviour through events other than direct DNA sequence changes, as the ones previously described [413,414]. Rather, these modifications regulate patterns of gene expression by modifying DNA accessibility by transcription factors and chromatin structure [414,415]. These biochemical pathways are crucial to normal development and differentiation of distinct cell lineages in the adult organism, rendering epigenetic mechanisms (EMs) an important tumorigenic effect [413,414,415].
Noteworthy, lifestyle changes influence epigenetic regulation of TR [416]. For instance, chronic psychological stress is believed to contribute to telomere shortening in humans at different life stages [417,418] in an apparent dose-dependent way [419], and diverse dietary compounds can have an impact on TERT by modulating the activity of DNA methyltransferases (DNMTs) and histone-modifying enzymes [420,421].
Nowadays, the most studied epigenetic alterations associated with neoplastic phenotype include DNA methylation and micro RNA (miRNA) mediated targeting of various genes [413,414]. Reactivation of telomerase is controlled by these mechanisms [420].
DNA methylation was the first identified EM [415,420]. This epigenetic process is crucial in gene expression, and errors in its pattern are tightly related to tumour initiation and progression [31,415,420]. Generally, via DNMTs, DNA methylation occurs genome-wide in non-coding regions, at CpG sites, which occur densely in regions known as CpG islands [31,415,422]. These CpG islands are located within gene promoter regions of approximately 60–70% of human genes and facilitate their expression by enabling the interaction with transcription factors [415,420].
Tumour cells can take advantage of two patterns of DNA methylation [415]: hypermethylation—increased methylation of CpG islands, generally associated with gene silencing of tumour suppressors such as p16 [423], MLH1 [424,425], and MGMT [72]—and hypomethylation—an overall decrease in global DNA methylation pattern, typically associated with overexpression of proto-oncogenes and growth factors [426].
Paradoxically, other genes, such as TERT, represent an exception to this regulation, as some authors reported that TERTp hypermethylation correlates with TERT overexpression in telomerase-positive cells [72,415,427,428], while the absence of TERT methylation was reported in some TERT-negative tumours and TERT-negative normal cells [28,72,429]. These findings are very dependent on the tissue of origin, since some authors did not find any correlation between mean methylation levels or hypermethylation and TERT expression levels in sporadic gastric adenocarcinoma, NOS [430], various histotypes of cervical (adenocarcinoma, SCC, adenosquamous, and carcinofibroma) [431] and ovarian (serous and cystoadenomas, endometrioid, and clear cell) [431,432] carcinomas; in gliomas (including WHO grades II–IV astrocytic and WHO grades II–III oligodendroglial tumours) some results are contradictory [72]. It seems that TERTp methylation is essential for TERT expression, and thus telomerase activity [420]. Indeed, in most TERT-positive tumour cells, most of the TERTp region contains hypermethylated CpG islands [420].
The degree at which the TERT promoter is methylated plays a role in carcinogenesis [420]. Hypermethylated states can prevent transcriptional repressors [433], such as CTCF [434], SIN3A, or MAZ [435] from binding to the target DNA-binding sites in the region. Lower methylation level allows the linkage of transcriptional repressors, resulting in shorter telomeres and lower telomerase activity [436]. Downregulation of DNMTs, which reduces the hypermethylated state of the TERTp by allowing repressor binding, is the proposed mechanism for telomerase reactivation [45,420].
There is a strong association between TERT methylation and telomerase activity in some tumour types, including B-cell lymphocytic leukaemia [437], colorectal carcinoma, NOS [438] and pancreatic ductal adenocarcinoma [439]. Colorectal carcinoma displays a higher level of cell methylation within the TERT promoter and high degree tumours with TERTp methylation reveal high telomerase activity [440]. Both DNA methylation and histone modification seem to operate TERT regulation in HCC [441]. Fan et al. [433] reported that TERTp CpG methylation may represent an alternative pathway to TERTp mutations in cutaneous melanoma, NOS.
Overall, the most characterized TERTp hypermethylation at specific CpG islands, also named as THOR (TERT hypermethylated oncological region), has been reported to have diagnostic and prognostic value in prostate, NOS [442] and pancreatic (exocrine and endocrine, NOS) cancers [443]. Methylation of the TERTp has also been suggested as a biomarker for malignancy and patient outcome in paediatric gliomas, such as pilocytic astrocytoma, medulloblastoma, NOS, ependymoma, choroid plexus carcinoma, among others [338].
Specific non-coding RNA interaction with TERT has been reported in multiple types of tumours [420]. TERT regulation by miRNA was summarized by Lewis et al. [420]. It was proposed that miRNAs can post-transcriptionally alter TERT transcripts directly or indirectly, affecting regulatory transcription factors [5,420]. TERRA transcripts are other types of non-coding RNAs found in eukaryotes that will be further addressed due to their association in ALT.
2.2.6. Alternative Lengthening of Telomeres
About 10 to 15% of tumours achieve immortalization through a telomerase-independent mechanism of telomere lengthening—alternative lengthening of telomeres [34], which was first detected in a telomerase-null mutant yeast [444] and subsequently reported in human tumours and tumour-derived cell lines [35,445]. ALT-positive cells are dependent on the activation of a homologous recombination DNA-repair mechanism to maintain telomere length. The knowledge regarding this mechanism has grown gradually throughout the years [34,444,445,446,447]. These cells are characterized by specific phenotypic features, such as heterogeneous telomere lengths [448,449], ALT-associated promyelocytic bodies (APBs) with decreased telomeric repeat binding factor 2 (TRF2) density [450,451], and telomere recombination with the presence of extrachromosomal (linear and circular) telomeric repeats [452]. Telomere-specific FISH (tel-FISH), APBs immunofluorescence and ALT-associated molecules (mainly C-circles) detection assays are some of the most commonly used techniques to detect ALT; tel-FISH and APBs immunofluorescence can be used in combination [453,454]. When reporting ALT phenotype (Table 4 and Figure 2), it should be kept in mind that, due to the variety of methodologies used, the positivity threshold may vary among studies.
Tumours of mesenchymal origin are reported to activate ALT more frequently, which may be explained by the fact that mesenchymal stem cells express minimal or undetectable telomerase, i.e., their lineage seems dependent on the activation of an alternative mechanism to maintain TL [55,453]. Recent studies reported that mutations in the ATRX or DAXX genes that encode chromatin remodelling proteins essential for the deposition of the histone variant H3.3 at telomeric and pericentromeric regions of the genome influence the activation and maintenance of ALT in pNETs [313,455], paediatric glioblastoma [337], and a spectrum of other tumours [55,321,322]. ATRX/DAXX mutations are thus suggested to be strong contributors to the activation of the ALT pathway. New evidence of epigenetic mechanisms affecting ALT were recently reported, such as microRNA regulation, as reviewed by Naderlinger et al. [5]. This mechanism has also been reported to correlate with high levels of TERRA [456].
The prevalence of ALT phenotype in tumour subtypes is shown in Table 4. ALT presents a high frequency in CNS tumours, such as diffuse and anaplastic astrocytomas, NOS (52% and 44%, respectively), adult and paediatric forms of glioblastoma (15% and 30%), among other types. The presence of ALT mechanism in GBs identifies less aggressive tumours with a longer patient survival, being associated at the same type to younger patients [298,301,303]. At variance, NBLs showing telomere elongation by this mechanism (50%) are characterized by unfavourable prognosis and resistance to chemotherapy [342]. Regarding ATRX status, high-grade astrocytomas have been reported with concomitant ALT phenotype and ATRX loss, in both adult and paediatric tumours [295,296,298,337]. Such findings point towards a central role of ATRX in ALT in CNS tumours [337]. A recent study by Fogli et al. [302] reported the presence of ALT in high-grade gliomas associated with IDH1/2 mutation, O-6-Methylguanine-DNA Methyltransferase (MGMT) methylation, absence of functional ATRX protein and elevated TERRA levels, supporting the need for more studies in these tumours, since their molecular background seems to have major importance in the stratification of patient prognosis.
Various subtypes of tumours that are reported with high frequencies of TERTp mutations, as previously addressed, do not present ALT.
ALT phenotype has also been reported to be highly prevalent in a wide variety of soft tissue and bone tumours: dedifferentiated liposarcoma (33%), pleomorphic liposarcoma (80%), myxofibrosarcoma (76%), leiomyosarcoma (57%), osteosarcoma (63%), malignant peripheral nerve sheath tumours (63%), among others. It has been associated with worse patient prognosis in some of these tumours [319,320,329], and it has been also correlated with loss of ATRX expression [322]. Bone and soft tissue sarcomas show a relatively lower or even absent frequency of TERTp mutations, with the exception of myxoid liposarcoma, that presents a high prevalence of TERTp mutations (67%) and a lower prevalence of ALT (15%).
Pancreatic neuroendocrine tumours are also reported to activate ALT frequently (30%). In this type of tumours, a consistent correlation between ALT phenotype and inactivation of either ATRX or DAXX has been reported [309,311,312,314]; however, ALT-positive cases that preserve expression of ATRX and DAXX indicates the presence of other activators [5]. ALT is also suggested to predict metastatic disease and poor survival in these tumours [306,309,311,312,334].
Overall, ALT phenotype is extremely rare in carcinomas; it has been described in certain subtypes such as ductal breast carcinoma (2%), HCC (7%), clear cell carcinoma, and endometrioid carcinoma of the ovary (4% and 1%, respectively) and chromophobe and sarcomatoid carcinomas of the kidney (9% and 7%, respectively). It is also present in malignant melanoma, NOS (7%), small cell neuroendocrine carcinoma of the bladder (23%), and in medullary thyroid carcinoma (26%). Omori et al. [305] reported a prevalence of 38% in gastric adenocarcinoma, NOS, but a subsequent study with a higher number of cases reported a negative ALT phenotype in such tumours [55].
Noteworthy, ALT was observed not to be present in several benign tumours of different origins, namely colon, hepatocellular, thyroid, and parathyroid adenomas [55]. There are still a lot of unanswered questions about these ALT mechanisms, mainly the ones regarding the molecular basis of its activation in tumour cells with wildtype ATRX or DAXX. The tumours with higher prevalence of ALT are reported as the least TERTp-mutated types, with the exception of gliomas (including WHO grades II–IV diffuse astrocytic and oligodendroglial tumours), in which a high frequency is observed for both mechanisms of cell immortalization. As illustrated in Figure 2, regarding tumours from different origins, the landscape of the distribution of TMMs by organ/anatomical site is quite diverse and with different cumulative prevalence. Evidences regarding mutual exclusivity of TERTp mutations and ALT phenotype in several types of tumours point towards the fact that when cells do not rely on telomerase activation to achieve immortalization, they activate the ALT mechanism. Some studies reported concomitant TERT expression and ALT activation (in adrenocortical carcinoma [308], NBL [317], osteosarcoma [327], nephroblastoma [330]), without clarifying the mechanism underlying telomerase reactivation. A recent study by Hayward et al. [187] has reported unexpected findings in a subset of cutaneous melanomas, in which nine in 10 ATRX-mutated cases also presented TERTp mutations, but these are novel findings that require further clarification.
2.2.7. Non-Defined Telomere Maintenance Mechanism
Data from TMM analyses gathered in the last years unveiled a phenotype in which both telomerase (or TERT) expression and ALT were reported as absent, pointing to a novel TMM: the non-defined telomere maintenance mechanism (NDTMM). Glioblastoma [303,457,458], osteosarcoma [328,459], metastases of cutaneous melanoma [460,461], and other tumour types [35,45] presented such a phenotype. Interestingly, Royds et al. [462] reported a NDTMM in GBs as a distinctive phenotype characterized by reduced patient survival, association with a polymorphism in CDKN2A and rarely IDH1-mutated. Analysing all the information previously described in this review, we recognized that some of the most incident cancers worldwide do not present any reported TMM (Table 6), what could be due to a failure in detection or, alternatively, represent a NDTMM. Noteworthy, the NDTMM frequencies reported in Figure 2 were obtained assuming the reported TMMs as mutually exclusive, what, as aforementioned, may not be transversal to all tumour cases. For this reason, and since most studies aim to assess a single TMM, the frequencies here reported are most likely underestimated. Nonetheless, these results represent a large proportion in several tumour types that must not be neglected. Tumours harbouring a NDTMM do not always present the same telomeric features [45], raising the question of which mechanism(s) is behind the maintenance or even if one exists, warranting the need for more studies on the matter. TERRA molecules may play a role in NDTMM. These are nuclear long noncoding RNAs (lncRNAs) found in all eukaryotes [5,37,463,464] that contain subtelomeric and telomeric UUAGGG-repeats transcribed by RNA polymerase II from the subtelomere towards the telomere [463,465,466]. They can regulate genome function by recruiting chromatin modifiers, regulating protein activity as trans-acting factors, and performing architectural functions [39,467]. TERRAs are also proposed to bind the telomerase core components, TERT and TERC [468], with stronger affinity for the later [469]. TERRA appears to integrate all lncRNAs functions into a single transcript responsible for telomere maintenance regulation in response to cellular signals [39].
Overall, TERRA molecules have been implicated in: (1) heterochromatin formation [470,471]; (2) direct inhibition of telomerase, by potential competition with the telomeric substrate for telomerase interactions [37,463,466,472]; (3) telomere protection [470,473]; (4) telomere replication in altered ALT cells lacking ATRX [474]; (5) telomere elongation by HR through the formation of DNA-TERRA hybrids [475]; and (6) participation in DDR activated by dysfunctional telomeres [471].
Naderlinger et al. [5] pointed to the enticing rational distinction between the potential use of these multi-featured molecules as templates for a new mechanism of telomeric synthesis. Luke et al. [464] suggested that instead of an essential or a permanent constituent of the telomeric chromatin, TERRA may have a transient regulatory role depending on telomeres’ specific functional state, by detecting TERRA molecules by RNA-FISH only in a subset of telomeres at human and mouse chromosome ends. Also, Rippe et al. [39] proposed that TERRA functions might be regulated in a telomere state-dependent manner because different telomere states may result in altered access of TERRA regulators to different telomere types: (a) at normal-length telomeres, TERRA appears to inhibit its own expression through EMs, by recruiting factors that promote a repressive chromatin state via the transcription-silencing network played by histone methyltransferase SUV39H, trimethylated H3K9 histone (H3K9me3) (essential for telomeres that use telomerase as a TMM), and heterochromatin proteins HP1 [5,37,39,476]; (b) when telomeres are shortened or damaged, TERRA levels increase, possibly due in part to their inability to play TPE-OLD, and also to a deactivation of autorepressive mechanisms (decrease of H3K9me3 levels [476] or depletion of TRF2 [476,477], a DNA-binding sheltering subunit) [39,476,478]; (c) when ALT pathway is responsible for telomere maintenance, TERRA expression levels appears more highly expressed than telomerase-positive cells [37,39,479]. It was proposed that the association of TERRA with telomeres in ALT cells is controlled by an interlinked network of TERRA, ATRX, H3K9me3, and TRF2 [39]. As already discussed, ALT cells highly express TERRA [456] and have loss of functional ATRX and incorporation of the histone H3.3 [480]. Consequently, H3K9me3 heterochromatin modification may decrease and ALT-associated decrease density of TRF2 [451] may contribute to raising TERRA levels, by relieving the TRF2-depedent TERRA silencing network [39]. ATRX depletion may stabilize TERRA’s association with telomeres, thus leading to eventual replication stress and increased replication-fork stalling [39]. Still, no clear correlation between ATRX and global TERRA expression levels is apparent [475,479,480]. Low TERRA levels, in combination with low to absent TR, were tentatively associated with favourable patient prognosis in a cohort of patients with grade II–IV astrocytomas [456].
3. Final Remarks
This extensive data collection allowed us to characterize the current panorama of TMMs in human cancers, in what regards to their prevalence, association to histopathological and molecular tumour features, prognostic assessment, and impact on clinical management.
TERTp mutations are the most frequent somatic non-coding alterations harboured by a wide spectrum of human tumours, namely of CNS, thyroid, skin, bladder, and liver. The collected data disclose remarkable differences of prevalence of TERTp mutations in histotypes from the same organs, as well as different TMMs within the same histotypes. The reason(s) for such differences remains unclear. One of the most important results concerning TERTp mutations are their frequent association to worse prognostic features and poorer patient survival. This finding indicates that TERTp mutations may be used as a biomarker for patient stratification in some cancers. Such mutations can arise in the context of malignant transformation in certain histotypes (e.g., liver), but overall, they represent a late event in most cancers. In tumours arising from tissues that are highly exposed to environmental factors (e.g., skin and bladder), TERTp mutations represent an early event. At variance with this aforementioned influence of TERTp mutations in many human cancers, there are some histotypes that do not present such alterations. The absence of selection for TERTp mutations can be partially explained by the fact that such tumours occur in tissues with fast cellular renewal, such as gastrointestinal or haematological malignancies. In the later context, the telomere length needs to be regularly maintained and may present intrinsic telomerase activity, making the existence of an activating telomere maintenance mechanism less important as a means for providing an additional selective advantage to cancer cells. The collected data show that for some tumours, TERTp mutations can be additionally modulated by TERTp germline genetic variations. Actually, such SNPs have been reported to impact the prognosis of TERTp-mutated tumours (e.g., urothelial bladder carcinomas and glioblastomas). The SNPs that modulate TERT transcriptional capacity are not restricted to the TERTp: as GWA studies have demonstrated, there are TERT germline genetic variations that lead to an increased risk of developing several cancer types. Since the results are sometimes conflicting with regard to the same histotypes, the interpretation of the impact of such polymorphisms must be carefully balanced, taking into consideration population (or ethnic) disparities and, ultimately, tumour genetic backgrounds.
TERT and TERC amplifications and TERT rearrangements were found in a small percentage of the reviewed cases. The scarce knowledge about these mechanisms determines the need for studying larger series in order to evaluate the real impact and frequency of such findings. The data obtained up to now provide promising evidence to be used as a diagnostic and prognostic tool in uterine malignancies and neuroblastomas, respectively, a feature that may be incorporated in the future clinical practice.
As previously indicated, ALT is highly prevalent in tumours of mesenchymal origin (e.g., soft tissues and bone tumours). Striking exceptions were grades II to IV diffuse astrocytic and oligodendroglial tumours, which are prone to exhibit TERTp mutations and ALT, usually displaying mutual exclusivity. Both alterations aid in the prognostic stratification of the observed patients. The aforementioned tumour types are the best examples of the role played by telomere status on prognosis using multiple maintenance mechanisms. ATRX inactivating mutations are intrinsically linked to ALT in CNS tumours and other tumour histotypes (e.g., pNETs). However, in ATRX-wildtype tumours, the mechanisms underlying ALT activation remain to be elucidated. Until recently, there was a mutual exclusivity of TERTp mutations and ALT activation, but recent studies reported their concomitant presence. Ongoing studies evaluating tumour inter- and intra-heterogeneity will be important to clarify the aforesaid category to find if subclones within a tumour or even if the same cell may at some point harbour both mechanisms simultaneously and possibly select one of them afterwards.
It was noteworthy that tumours of the breast, stomach, small intestine, colon and rectum, exocrine pancreas, lung, and prostate, that represent some of the most frequent tumours worldwide, were also those that presented a low frequency or absence of known TMMs. Breast and colorectal tumours were found to have a high prevalence of TERT amplifications, although the respective cohorts were too small to solely assign this mechanism to telomere maintenance. In the study of Barthel et al. [45], 22% of the cases had no detectable TERT expression nor alterations in the genes directly linked to ALT activation—ATRX and DAXX. Barthel et al. [45] hypothesized that not all tumours harbour immortalized cells with a TMM or that additional mechanisms may yet exist, ‘something’ we may designate as a non-defined TMM. Such (yet) undefined TMM may involve RB1 or TP53 alterations due to telomere-driven genomic instability, that may surpass the DNA repair mechanisms [45].
Finally, we have emphasized throughout the text the intriguing questions that remain to be answered, such as the reasons behind the (a) gradual increase in TMM activation with grade progression; (b) high-grade dependence of some histotypes for specific TMMs; (c) the homogenous distribution of TMMs frequencies among very different tumour grades; (d) better prognosis conferred by TMMs in exceptional cases, and, at last; for the (e) apparent absence of TMMs in some tumours. Pursuing these questions will open new avenues in the understanding of mechanisms that may surpass the classical TMMs function but with the same end-result—to assure cancer cell immortality.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4425/9/5/241/s1, Table S1: Prevalence of different TMMs by organ/system (complete data).
Acknowledgments
This work was supported by PhD grant from FCT—Fundação para a Ciência e a Tecnologia to T.B.G. (SFRH/BD/129431/2017). This work was also financed by FEDER—Fundo Europeu de Desenvolvimento Regional funds through the COMPETE 2020—Operacional Programme for Competitiveness and Internationalization (POCI), Portugal 2020, and by Portuguese funds through FCT in the framework of the project “Institute for Research and Innovation in Health Sciences” (POCI-01-0145-FEDER-007274). Further funding was obtained from the project “Advancing cancer research: from basic knowledgment to application”; NORTE-01-0145-FEDER-000029; “Projetos Estruturados de I&D&I”, funded by Norte 2020—Programa Operacional Regional do Norte. Further funded by the European Regional Development Fund (ERDF) through the Operational Programme for Competitiveness and Internationalization—COMPETE2020, and Portuguese national funds via FCT, under project POCI-01-0145-FEDER-016390: CANCEL STEM”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Karlseder, J. Complex interactions between the DNA-damage response and mammalian telomeres. Nat. Struct. Mol. Biol. 2015, 22, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Kumar, R. TERT promoter mutations in telomere biology. Mutat. Res. 2017, 771, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Naderlinger, E.; Holzmann, K. Epigenetic Regulation of Telomere Maintenance for Therapeutic Interventions in Gliomas. Genes 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Unal, B.; Tergaonkar, V. Reactivation of telomerase in cancer. Cell. Mol. Life Sci. 2016, 73, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, J.; Cech, T.R. Finding the end: Recruitment of telomerase to telomeres. Nat. Rev. Mol. Cell Biol. 2013, 14, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Blasco, M.A. Telomeres and human disease: Ageing, cancer and beyond. Nat. Rev. Genet. 2005, 6, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, S.; Stohr, B.A. The role of telomere biology in cancer. Annu. Rev. Pathol. 2013, 8, 49–78. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; de Lange, T. Removal of shelterin reveals the telomere end-protection problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. How telomeres solve the end-protection problem. Science 2009, 326, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Erdel, F.; Kratz, K.; Willcox, S.; Griffith, J.D.; Greene, E.C.; de Lange, T. Telomere Recognition and Assembly Mechanism of Mammalian Shelterin. Cell Rep. 2017, 18, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Blasco, M.A. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat. Rev. Cancer 2011, 11, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Low, K.C.; Tergaonkar, V. Telomerase: Central regulator of all of the hallmarks of cancer. Trends Biochem. Sci. 2013, 38, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Bryan, T.M.; Reddel, R.R. Increased copy number of the TERT and TERC telomerase subunit genes in cancer cells. Cancer Sci. 2008, 99, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.C.; Cech, T.R. Human telomerase: Biogenesis, trafficking, recruitment, and activation. Genes Dev. 2015, 29, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.B.; Graham, M.E.; Lovrecz, G.O.; Bache, N.; Robinson, P.J.; Reddel, R.R. Protein composition of catalytically active human telomerase from immortal cells. Science 2007, 315, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kim, N.K.; Feigon, J. Architecture of human telomerase RNA. Proc. Natl. Acad. Sci. USA 2011, 108, 20325–20332. [Google Scholar] [CrossRef] [PubMed]
- Holohan, B.; Wright, W.E.; Shay, J.W. Telomeropathies: An emerging spectrum disorder. J. Cell Biol. 2014, 205, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Pinto, V.; Celestino, R.; Reis, M.; Populo, H.; Boaventura, P.; Melo, M.; Catarino, T.; Lima, J.; Lopes, J.M.; et al. Telomerase promoter mutations in cancer: An emerging molecular biomarker? Virchows Arch. 2014, 465, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G. Telomere dynamics and aging. Prog. Mol. Biol. Transl. Sci. 2014, 125, 89–111. [Google Scholar] [PubMed]
- Podlevsky, J.D.; Chen, J.J. It all comes together at the ends: Telomerase structure, function, and biogenesis. Mutat. Res. 2012, 730, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, M.I.; Shcherbakova, D.M.; Dontsova, O.A. Telomerase: Structure, functions, and activity regulation. Biochemistry 2011, 75, 1563–1583. [Google Scholar] [CrossRef]
- Bourgeron, T.; Xu, Z.; Doumic, M.; Teixeira, M.T. The asymmetry of telomere replication contributes to replicative senescence heterogeneity. Sci. Rep. 2015, 5, 15326. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Fujiwara, T.; Inoue, M. Understanding and exploiting hTERT promoter regulation for diagnosis and treatment of human cancers. Cancer Sci. 2008, 99, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Blasco, M.A. Role of Rb family in the epigenetic definition of chromatin. Cell Cycle 2005, 4, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Blasco, M.A. Telomere epigenetics: A higher-order control of telomere length in mammalian cells. Carcinogenesis 2004, 25, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Jaco, I.; Fraga, M.F.; Chen, T.; Li, E.; Esteller, M.; Blasco, M.A. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006, 8, 416–424. [Google Scholar] [CrossRef] [PubMed]
- García-Cao, M.; O’Sullivan, R.; Peters, A.H.; Jenuwein, T.; Blasco, M.A. Epigenetic regulation of telomere length in mammalian cells by the SUV39H1 and SUV39H2 histone methyltransferases. Nat. Genet. 2004, 36, 94. [Google Scholar] [CrossRef] [PubMed]
- Robin, J.D.; Ludlow, A.T.; Batten, K.; Magdinier, F.; Stadler, G.; Wagner, K.R.; Shay, J.W.; Wright, W.E. Telomere position effect: Regulation of gene expression with progressive telomere shortening over long distances. Genes Dev. 2014, 28, 2464–2476. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative lengthening of telomeres: Models, mechanisms and implications. Nat. Rev. Genet. 2010, 11, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Bryan, T.M.; Englezou, A.; Dalla-Pozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997, 3, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.A.; Watson, C.M.; Noble, J.R.; Pickett, H.A.; Tam, P.P.; Reddel, R.R. Alternative lengthening of telomeres in normal mammalian somatic cells. Genes Dev. 2013, 27, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Napier, C.E.; Vryer, R.; Dimitriadis, E.; Manuelpillai, U.; Sharkey, A.; Craig, J.M.; Reddel, R.R.; Saffery, R. DNA methylation mediated up-regulation of TERRA non-coding RNA is coincident with elongated telomeres in the human placenta. Mol. Hum. Reprod. 2016, 22, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Slatter, T.L.; Tan, X.; Yuen, Y.C.; Gunningham, S.; Ma, S.S.; Daly, E.; Packer, S.; Devenish, C.; Royds, J.A.; Hung, N.A. The alternative lengthening of telomeres pathway may operate in non-neoplastic human cells. J. Pathol. 2012, 226, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Rippe, K.; Luke, B. TERRA and the state of the telomere. Nat. Struct. Mol. Biol. 2015, 22, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.J.; Perdigones, N. Telomere biology and translational research. Transl. Res. 2013, 162, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Pestana, A.; Vinagre, J.; Sobrinho-Simoes, M.; Soares, P. TERT biology and function in cancer: Beyond immortalisation. J. Mol. Endocrinol. 2017, 58, R129–R146. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.M.; Reddel, R.R. Telomere maintenance mechanisms and cellular immortalization. Curr. Opin. Genet. Dev. 1999, 9, 97–103. [Google Scholar] [CrossRef]
- Cairney, C.J.; Keith, W.N. Telomerase redefined: Integrated regulation of HTR and hTERT for telomere maintenance and telomerase activity. Biochimie 2008, 90, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Neidle, S. Quadruplex Nucleic Acids as Novel Therapeutic Targets. J. Med. Chem. 2016, 59, 5987–6011. [Google Scholar] [CrossRef] [PubMed]
- Jager, K.; Walter, M. Therapeutic targeting of telomerase. Genes 2016, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating cancer by targeting telomeres and telomerase. Antioxidants 2017, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Sahm, F.; Capper, D.; Reuss, D.; Sturm, D.; Jones, D.T.; Kool, M.; Northcott, P.A.; Wiestler, B.; Bohmer, K.; et al. Distribution of TERT promoter mutations in pediatric and adult tumors of the nervous system. Acta Neuropathol. 2013, 126, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinagre, J.; Almeida, A.; Populo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013, 4, 2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Zheng, C.; Lindvall, C.; Hou, M.; Ekedahl, J.; Lewensohn, R.; Yan, Z.; Yang, X.; Henriksson, M.; Blennow, E.; et al. Frequent amplification of the telomerase reverse transcriptase gene in human tumors. Cancer Res. 2000, 60, 6230–6235. [Google Scholar] [PubMed]
- Saito, T.; Akaike, K.; Kurisaki-Arakawa, A.; Toda-Ishii, M.; Mukaihara, K.; Suehara, Y.; Takagi, T.; Kaneko, K.; Yao, T. TERT promoter mutations are rare in bone and soft tissue sarcomas of Japanese patients. Mol. Clin. Oncol. 2016, 4, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Hertwig, F.; Roels, F.; Dreidax, D.; Gartlgruber, M.; Menon, R.; Kramer, A.; Roncaioli, J.L.; Sand, F.; Heuckmann, J.M.; et al. Telomerase activation by genomic rearrangements in high-risk neuroblastoma. Nature 2015, 526, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the alternative lengthening of telomeres telomere maintenance mechanism in human cancer subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Greenberg, R.A. ALTernative telomere maintenance and cancer. Trends Cancer 2015, 1, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Nabais, J.; Pinheiro, J.; Batista, R.; Oliveira, R.C.; Goncalves, A.P.; Pestana, A.; Reis, M.; Mesquita, B.; Pinto, V.; et al. TERT promoter mutations in pancreatic endocrine tumours are rare and mainly found in tumours from patients with hereditary syndromes. Sci. Rep. 2016, 6, 29714. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Ogawa, R.; Yoshida, H.; Maeshima, A.; Kanai, Y.; Kinoshita, T.; Hiraoka, N.; Sekine, S. TERT promoter mutations are frequent and show association with MED12 mutations in phyllodes tumors of the breast. Br. J. Cancer 2015, 113, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Nozad, S.; Sheehan, C.E.; Gay, L.M.; Elvin, J.A.; Vergilio, J.A.; Suh, J.; Ramkissoon, S.; Schrock, A.B.; Hirshfield, K.M.; Ali, N.; et al. Comprehensive genomic profiling of malignant phyllodes tumors of the breast. Breast Cancer Res. Treat. 2017, 162, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Gay-Bellile, M.; Veronese, L.; Combes, P.; Eymard-Pierre, E.; Kwiatkowski, F.; Dauplat, M.M.; Cayre, A.; Privat, M.; Abrial, C.; Bignon, Y.J.; et al. TERT promoter status and gene copy number gains: Effect on TERT expression and association with prognosis in breast cancer. Oncotarget 2017, 8, 77540–77551. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Pirozzi, C.J.; Healy, P.; Reitman, Z.J.; Lipp, E.; Rasheed, B.A.; Yang, R.; Diplas, B.H.; Wang, Z.; Greer, P.K.; et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget 2014, 5, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hosen, I.; Volz, F.; Hemminki, K.; Weyerbrock, A.; Kumar, R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget 2015, 6, 10617–10633. [Google Scholar] [CrossRef] [PubMed]
- Pekmezci, M.; Rice, T.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Hansen, H.; Sicotte, H.; Kollmeyer, T.M.; McCoy, L.S.; Sarkar, G.; Perry, A.; Giannini, C.; et al. Adult infiltrating gliomas with WHO 2016 integrated diagnosis: Additional prognostic roles of ATRX and TERT. Acta Neuropathol. 2017, 133, 1001–1016. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Narita, Y.; Fukushima, S.; Tateishi, K.; Matsushita, Y.; Yoshida, A.; Miyakita, Y.; Ohno, M.; Collins, V.P.; Kawahara, N.; et al. Upregulating mutations in the TERT promoter commonly occur in adult malignant gliomas and are strongly associated with total 1p19q loss. Acta Neuropathol. 2013, 126, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Hosen, I.; Gousias, K.; Rachakonda, S.; Heidenreich, B.; Gessi, M.; Schramm, J.; Hemminki, K.; Waha, A.; Kumar, R. TERT promoter mutations: A novel independent prognostic factor in primary glioblastomas. Neuro-Oncology 2015, 17, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, Y.; Liu, Y.; Liu, X.; Zhang, C.; Wang, L.; Li, S.; Ma, J.; Jiang, T. Brain regions associated with telomerase reverse transcriptase promoter mutations in primary glioblastomas. J. Neuro-Oncol. 2016, 128, 455–462. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Wu, Y.; Chang, K.; Shi, X.; Chen, X.D.; Yan, W.; Li, R. Paradoxical prognostic impact of TERT promoter mutations in gliomas depends on different histological and genetic backgrounds. CNS Neurosci. Ther. 2017, 23, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Mosrati, M.A.; Malmstrom, A.; Lysiak, M.; Krysztofiak, A.; Hallbeck, M.; Milos, P.; Hallbeck, A.L.; Bratthall, C.; Strandeus, M.; Stenmark-Askmalm, M.; et al. TERT promoter mutations and polymorphisms as prognostic factors in primary glioblastoma. Oncotarget 2015, 6, 16663–16673. [Google Scholar] [CrossRef] [PubMed]
- Nencha, U.; Rahimian, A.; Giry, M.; Sechi, A.; Mokhtari, K.; Polivka, M.; Schmitt, Y.; Di Stefano, A.L.; Alentorn, A.; Labussiere, M.; et al. TERT promoter mutations and rs2853669 polymorphism: Prognostic impact and interactions with common alterations in glioblastomas. J. Neuro-Oncol. 2016, 126, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, G.; Shan, Y.; Hartmann, C.; von Deimling, A.; Xing, M. Highly prevalent TERT promoter mutations in bladder cancer and glioblastoma. Cell Cycle 2013, 12, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Passow, J.E.; Lachance, D.H.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Sicotte, H.; Pekmezci, M.; Rice, T.; Kosel, M.L.; Smirnov, I.V.; et al. Glioma groups based on 1p/19q, IDH, and TERT promoter mutations in tumors. N. Engl. J. Med. 2015, 372, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Narita, Y.; Takami, H.; Fukushima, S.; Matsushita, Y.; Yoshida, A.; Miyakita, Y.; Ohno, M.; Shibui, S.; Ichimura, K. TERT promoter mutations rather than methylation are the main mechanism for TERT upregulation in adult gliomas. Acta Neuropathol. 2013, 126, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Cai, J.; Yan, W.; Zhang, W.; Wang, Y.; Chen, B.; Li, G.; Li, S.; Wu, C.; Yao, K.; et al. Classification based on mutations of TERT promoter and IDH characterizes subtypes in grade II/III gliomas. Neuro-Oncology 2016, 18, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Labussiere, M.; Di Stefano, A.L.; Gleize, V.; Boisselier, B.; Giry, M.; Mangesius, S.; Bruno, A.; Paterra, R.; Marie, Y.; Rahimian, A.; et al. TERT promoter mutations in gliomas, genetic associations and clinico-pathological correlations. Br. J. Cancer 2014, 111, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.S.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Meng, Q.; Ye, Z.Y.; Wang, W.; Jiang, X.T.; et al. Recurrent TERT promoter mutations identified in a large-scale study of multiple tumour types are associated with increased TERT expression and telomerase activation. Eur. J. Cancer 2015, 51, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Batista, R.; Cruvinel-Carloni, A.; Vinagre, J.; Peixoto, J.; Catarino, T.A.; Campanella, N.C.; Menezes, W.; Becker, A.P.; de Almeida, G.C.; Matsushita, M.M.; et al. The prognostic impact of TERT promoter mutations in glioblastomas is modified by the rs2853669 single nucleotide polymorphism. Int. J. Cancer 2016, 139, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Li, G.; Qu, Y.; Wang, M.; Cui, B.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations and long telomere length predict poor survival and radiotherapy resistance in gliomas. Oncotarget 2016, 7, 8712–8725. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Koh, J.; Kim, S.I.; Won, J.K.; Park, C.K.; Choi, S.H.; Park, S.H. The frequency and prognostic effect of TERT promoter mutation in diffuse gliomas. Acta Neuropathol. Commun. 2017, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Yamasaki, K.; Matsushita, Y.; Nakamura, T.; Shimokawa, A.; Takami, H.; Tanaka, S.; Mukasa, A.; Shirahata, M.; Shimizu, S.; et al. A combination of TERT promoter mutation and MGMT methylation status predicts clinically relevant subgroups of newly diagnosed glioblastomas. Acta Neuropathol. Commun. 2016, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Spiegl-Kreinecker, S.; Lotsch, D.; Ghanim, B.; Pirker, C.; Mohr, T.; Laaber, M.; Weis, S.; Olschowski, A.; Webersinke, G.; Pichler, J.; et al. Prognostic quality of activating TERT promoter mutations in glioblastoma: Interaction with the rs2853669 polymorphism and patient age at diagnosis. Neuro-Oncology 2015, 17, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Nonoguchi, N.; Ohta, T.; Oh, J.E.; Kim, Y.H.; Kleihues, P.; Ohgaki, H. TERT promoter mutations in primary and secondary glioblastomas. Acta Neuropathol. 2013, 126, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.E.; Woo, S.R.; Nam, H.; Nam, D.H.; Lee, J.H.; Joo, K.M. Preclinical and clinical implications of TERT promoter mutation in glioblastoma multiforme. Oncol. Lett. 2017, 14, 8213–8219. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Remke, M.; Ramaswamy, V.; Peacock, J.; Shih, D.J.; Koelsche, C.; Northcott, P.A.; Hill, N.; Cavalli, F.M.; Kool, M.; Wang, X.; et al. TERT promoter mutations are highly recurrent in SHH subgroup medulloblastoma. Acta Neuropathol. 2013, 126, 917–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kool, M.; Jones, D.T.; Jager, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. Genome sequencing of SHH medulloblastoma predicts genotype-related response to smoothened inhibition. Cancer Cell 2014, 25, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, J.C.; Schwalbe, E.C.; Potluri, S.; Bailey, S.; Williamson, D.; Clifford, S.C. TERT promoter mutation and aberrant hypermethylation are associated with elevated expression in medulloblastoma and characterise the majority of non-infant SHH subgroup tumours. Acta Neuropathol. 2014, 127, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Viana-Pereira, M.; Almeida, G.C.; Stavale, J.N.; Malheiro, S.; Clara, C.; Lobo, P.; Pimentel, J.; Reis, R.M. Study of hTERT and Histone 3 Mutations in Medulloblastoma. Pathobiology 2017, 84, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Goutagny, S.; Nault, J.C.; Mallet, M.; Henin, D.; Rossi, J.Z.; Kalamarides, M. High incidence of activating TERT promoter mutations in meningiomas undergoing malignant progression. Brain Pathol. 2014, 24, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Schrimpf, D.; Olar, A.; Koelsche, C.; Reuss, D.; Bissel, J.; Kratz, A.; Capper, D.; Schefzyk, S.; Hielscher, T.; et al. TERT Promoter Mutations and Risk of Recurrence in Meningioma. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, A.E.; Ober, K.; Dubbink, H.J.; Paridaens, D.; Naus, N.C.; Belunek, S.; Krist, B.; Post, E.; Zwarthoff, E.C.; de Klein, A. Prevalence and Implications of TERT Promoter Mutation in Uveal and Conjunctival Melanoma and in Benign and Premalignant Conjunctival Melanocytic Lesions. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6024–6030. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Scholz, S.; Sucker, A.; Song, M.; Susskind, D.; Grabellus, F.; Zimmer, L.; Hillen, U.; et al. TERT promoter mutations in ocular melanoma distinguish between conjunctival and uveal tumours. Br. J. Cancer 2013, 109, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Puig-Butille, J.A.; Schilling, B.; Livingstone, E.; Potrony, M.; Carrera, C.; Schimming, T.; Moller, I.; Schwamborn, M.; et al. TERT Promoter Mutation Status as an Independent Prognostic Factor in Cutaneous Melanoma. JNCI J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.L.; Thomasen, H.; Reis, H.; Moller, I.; Darawsha, R.; Muller, B.; Dekowski, D.; Sucker, A.; Schilling, B.; Schadendorf, D.; et al. Frequent TERT promoter mutations in ocular surface squamous neoplasia. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5854–5861. [Google Scholar] [CrossRef] [PubMed]
- Dono, M.; Angelini, G.; Cecconi, M.; Amaro, A.; Esposito, A.I.; Mirisola, V.; Maric, I.; Lanza, F.; Nasciuti, F.; Viaggi, S.; et al. Mutation frequencies of GNAQ, GNA11, BAP1, SF3B1, EIF1AX and TERT in uveal melanoma: Detection of an activating mutation in the TERT gene promoter in a single case of uveal melanoma. Br. J. Cancer 2014, 110, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Pilati, C.; Letouze, E.; Nault, J.C.; Imbeaud, S.; Boulai, A.; Calderaro, J.; Poussin, K.; Franconi, A.; Couchy, G.; Morcrette, G.; et al. Genomic profiling of hepatocellular adenomas reveals recurrent FRK-activating mutations and the mechanisms of malignant transformation. Cancer Cell 2014, 25, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Mallet, M.; Pilati, C.; Calderaro, J.; Bioulac-Sage, P.; Laurent, C.; Laurent, A.; Cherqui, D.; Balabaud, C.; Zucman-Rossi, J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat. Commun. 2013, 4, 2218. [Google Scholar] [CrossRef] [PubMed]
- Quaas, A.; Oldopp, T.; Harun, L.; Klingenfeld, C.; Krech, T.; Sauter, G.; Grob, T.J. Frequency of TERT promoter mutations in primary tumors of the liver. Virchows Arch. 2014, 465, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Han, S.; Meng, L.; Li, Z.; Zhang, X.; Wu, A. TERT promoter mutations lead to high transcriptional activity under hypoxia and temozolomide treatment and predict poor prognosis in gliomas. PLoS ONE 2014, 9, e100297. [Google Scholar] [CrossRef] [PubMed]
- Totoki, Y.; Tatsuno, K.; Covington, K.R.; Ueda, H.; Creighton, C.J.; Kato, M.; Tsuji, S.; Donehower, L.A.; Slagle, B.L.; Nakamura, H.; et al. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat. Genet. 2014, 46, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Ally, A.; Balasundaram, M.; Carlsen, R.; Chuah, E.; Clarke, A.; Dhalla, N.; Holt, R.A.; Jones, S.J.; Lee, D.; Ma, Y.; et al. Comprehensive and integrative genomic characterization of hepatocellular carcinoma. Cell 2017, 169, 1327–1341. [Google Scholar] [CrossRef] [PubMed]
- Cevik, D.; Yildiz, G.; Ozturk, M. Common telomerase reverse transcriptase promoter mutations in hepatocellular carcinomas from different geographical locations. World J. Gastroenterol. 2015, 21, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, A.; Furuta, M.; Shiraishi, Y.; Gotoh, K.; Kawakami, Y.; Arihiro, K.; Nakamura, T.; Ueno, M.; Ariizumi, S.; Nguyen, H.H.; et al. Whole-genome mutational landscape of liver cancers displaying biliary phenotype reveals hepatitis impact and molecular diversity. Nat. Commun. 2015, 6, 6120. [Google Scholar] [CrossRef] [PubMed]
- Schulze, K.; Imbeaud, S.; Letouze, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Chang, S.H.; Kim, W.Y.; Lim, S.D.; Kim, W.S.; Hwang, T.S.; Han, H.S. Frequent somatic TERT promoter mutations and CTNNB1 mutations in hepatocellular carcinoma. Oncotarget 2016, 7, 69267–69275. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, F.; Izzo, F.; Buonaguro, L.; Annunziata, C.; Tatangelo, F.; Botti, G.; Buonaguro, F.M.; Tornesello, M.L. Tumor specific mutations in TERT promoter and CTNNB1 gene in hepatitis B and hepatitis C related hepatocellular carcinoma. Oncotarget 2016, 7, 54253–54262. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guo, X.; Chen, Y.; Chen, G.; Ma, Y.; Huang, K.; Zhang, Y.; Zhao, Q.; Winkler, C.A.; An, P.; et al. Telomerase reverse transcriptase promoter mutations in hepatitis B virus-associated hepatocellular carcinoma. Oncotarget 2016, 7, 27838–27847. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhou, W.; Li, C.; Yang, Y.; Shang, Y.K.; Chen, C.; Zhang, J.; Yao, R.; Wang, P.; Wen, W.; et al. Promoter mutations and cellular distribution of telomerase in non-clear cell and clear cell hepatocellular carcinoma. Oncotarget 2017, 8, 26288–26297. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Park, T.I.; Jang, S.Y.; Park, S.Y.; Park, W.J.; Jung, S.J.; Lee, J.H. Clinicopathological characteristics of TERT promoter mutation and telomere length in hepatocellular carcinoma. Medicine 2017, 96, e5766. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Cheng, G.H.; Yu, J.Y.; Zheng, S.Z.; Sun, C.; Sun, Q.; Li, K.L.; Lin, Z.M.; Liu, T.T.; Li, P.; et al. The TERT promoter mutation incidence is modified by germline TERT rs2736098 and rs2736100 polymorphisms in hepatocellular carcinoma. Oncotarget 2017, 8, 23120–23129. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Shi, L.; Wang, D.; Zhang, B.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. Low frequency of TERT promoter mutations in a large cohort of gallbladder and gastric cancers. Int. J. Cancer 2014, 134, 2993–2994. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, Y.; Chen, Z.; Hu, X.; Zhou, F.; He, J. Low frequency of TERT promoter somatic mutation in 313 sporadic esophageal squamous cell carcinomas. Int. J. Cancer 2014, 134, 493–494. [Google Scholar] [CrossRef] [PubMed]
- Van Nistelrooij, A.M.; Zwarthoff, E.C.; Post, E.; Lurkin, I.; van Marion, R.; Korpershoek, E.; Biermann, K.; Wijnhoven, B.P.; Dinjens, W.N. Absence of TERT promoter mutations in esophageal adenocarcinoma. Int. J. Cancer 2014, 134, 2014–2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liang, X.; Bjorkholm, M.; Jia, J.; Xu, D. The absence of TERT promoter mutations in primary gastric cancer. Gene 2014, 540, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Eichenmuller, M.; Trippel, F.; Kreuder, M.; Beck, A.; Schwarzmayr, T.; Haberle, B.; Cairo, S.; Leuschner, I.; von Schweinitz, D.; Strom, T.M.; et al. The genomic landscape of hepatoblastoma and their progenies with HCC-like features. J. Hepatol. 2014, 61, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, T.T.; Sofiadis, A.; Juhlin, C.C.; Zedenius, J.; Hoog, A.; Larsson, C.; Xu, D.W. TERT Promoter Mutation as an Early Genetic Event Activating Telomerase in Follicular Thyroid Adenoma (FTA) and Atypical FTA. Cancer 2014, 120, 2965–2979. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bishop, J.; Shan, Y.; Pai, S.; Liu, D.; Murugan, A.K.; Sun, H.; El-Naggar, A.K.; Xing, M. Highly prevalent TERT promoter mutations in aggressive thyroid cancers. Endocr. Relat. Cancer 2013, 20, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.Z.; Liu, R.Y.; Liu, X.L.; Murugan, A.K.; Zhu, G.W.; Zeiger, M.A.; Pai, S.; Bishop, J. BRAFV600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J. Clin. Oncol. 2014, 32, 2718. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, G.; Ragazzi, M.; Frasoldati, A.; Piana, S.; Ciarrocchi, A.; Sancisi, V. TERT promoter mutations are associated with distant metastases in papillary thyroid carcinoma. Eur. J. Endocrinol. 2015, 172, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Muzza, M.; Colombo, C.; Rossi, S.; Tosi, D.; Cirello, V.; Perrino, M.; De Leo, S.; Magnani, E.; Pignatti, E.; Vigo, B.; et al. Telomerase in differentiated thyroid cancer: Promoter mutations, expression and localization. Mol. Cell. Endocrinol. 2015, 399, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Qasem, E.; Murugan, A.K.; Al-Hindi, H.; Xing, M.; Almohanna, M.; Alswailem, M.; Alzahrani, A.S. TERT promoter mutations in thyroid cancer: A report from a Middle Eastern population. Endocr. Relat. Cancer 2015, 22, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.J.; Kim, W.G.; Sim, S.; Lim, S.; Kwon, H.; Kim, T.Y.; Shong, Y.K.; Kim, W.B. Low prevalence of somatic tert promoter mutations in classic papillary thyroid carcinoma. Endocrinol. Metab. 2016, 31, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Hwang, T.S.; Choi, Y.-L.; Han, H.S.; Kim, W.S.; Jang, M.H.; Kim, S.K.; Yang, J.H. Prognostic significance of TERT promoter mutations in papillary thyroid carcinomas in a BRAFV600E mutation–prevalent population. Thyroid 2016, 26, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, J.; Lu, J.; Gao, J.; Ren, X.; Teng, L.; Duan, H.; Lin, Y.; Li, X.; Zhang, B.; et al. BRAFV600E and TERT promoter mutations in papillary thyroid carcinoma in chinese patients. PLoS ONE 2016, 11, e0153319. [Google Scholar] [CrossRef]
- Bullock, M.; Ren, Y.; O’Neill, C.; Gill, A.; Aniss, A.; Sywak, M.; Sidhu, S.; Delbridge, L.; Learoyd, D.; de Vathaire, F.; et al. TERT promoter mutations are a major indicator of recurrence and death due to papillary thyroid carcinomas. Clin. Endocrinol. 2016, 85, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Batista, R.; Peixoto, J.; Tavares, C.; Celestino, R.; Almeida, A.; Salgado, C.; Eloy, C.; et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J. Clin. Endocrinol. Metab. 2014, 99, E754–E765. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, M.S.; Schmitt, A.; Steinert, H.; Capper, D.; Moch, H.; Komminoth, P.; Perren, A. Tall cell papillary thyroid carcinoma: New diagnostic criteria and mutations in BRAF and TERT. Endocr. Relat. Cancer 2015, 22, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ganly, I.; Chan, T.A.; Mitsutake, N.; Matsuse, M.; Ibrahimpasic, T.; Ghossein, R.A.; Fagin, J.A. Frequent somatic TERT promoter mutations in thyroid cancer: Higher prevalence in advanced forms of the disease. J. Clin. Endocrinol. Metab. 2013, 98, E1562–E1566. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xing, M. Diagnostic and prognostic TERT promoter mutations in thyroid fine-needle aspiration biopsy. Endocr. Relat. Cancer 2014, 21, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, N.; Cao, J.; Sofiadis, A.; Dinets, A.; Zedenius, J.; Larsson, C.; Xu, D. The age- and shorter telomere-dependent TERT promoter mutation in follicular thyroid cell-derived carcinomas. Oncogene 2014, 33, 4978–4984. [Google Scholar] [CrossRef] [PubMed]
- George, J.R.; Henderson, Y.C.; Williams, M.D.; Roberts, D.B.; Hei, H.; Lai, S.Y.; Clayman, G.L. Association of TERT promoter mutation, but not BRAF mutation, with increased mortality in PTC. J. Clin. Endocrinol. Metab. 2015, 100, E1550–E1559. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chen, E.; Dong, S.; Cai, Y.; Zhang, X.; Zhou, Y.; Zeng, R.; Yang, F.; Pan, C.; Liu, Y.; et al. BRAF and TERT promoter mutations in the aggressiveness of papillary thyroid carcinoma: A study of 653 patients. Oncotarget 2016, 7, 18346–18355. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Kim, Y.E.; Ahn, S.; Kim, J.Y.; Ki, C.S.; Oh, Y.L.; Kim, K.; Yun, J.W.; Park, W.Y.; Choe, J.H.; et al. TERT promoter mutations and long-term survival in patients with thyroid cancer. Endocr. Relat. Cancer 2016, 23, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ibrahimpasic, T.; Boucai, L.; Sinha, R.; Knauf, J.A.; Shah, R.H.; Dogan, S.; Ricarte-Filho, J.C.; Krishnamoorthy, G.P.; Xu, B.; et al. Genomic and transcriptomic hallmarks of poorly differentiated and anaplastic thyroid cancers. J. Clin. Investig. 2016, 126, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.K.; Kwak, B.K.; Lim, J.A.; Lee, M.C.; Kim, M.J. TERT Promoter mutations and tumor persistence/recurrence in papillary thyroid cancer. Cancer Res. Treat. 2016, 48, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Kim, T.H.; Kim, S.W.; Ki, C.S.; Jang, H.W.; Kim, J.S.; Kim, J.H.; Choe, J.H.; Shin, J.H.; Hahn, S.Y.; et al. Comprehensive screening for PD-L1 expression in thyroid cancer. Endocr. Relat. Cancer 2017, 24, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Ki, C.S.; Kim, H.S.; Kim, K.; Choe, J.H.; Kim, J.H.; Kim, J.S.; Oh, Y.L.; Hahn, S.Y.; Shin, J.H.; et al. Refining dynamic risk stratification and prognostic groups for differentiated thyroid cancer with TERT promoter mutations. J. Clin. Endocrinol. Metab. 2017, 102, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Ki, C.S.; Hahn, S.Y.; Oh, Y.L.; Jang, H.W.; Kim, S.W.; Chung, J.H.; Shin, J.H. Ultrasonographic prediction of highly aggressive telomerase reverse transcriptase (TERT) promoter-mutated papillary thyroid cancer. Endocrine 2017, 57, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; Gaspar da Rocha, A.; Batista, R.; Vinagre, J.; Martins, M.J.; Costa, G.; Ribeiro, C.; Carrilho, F.; Leite, V.; Lobo, C.; et al. TERT, BRAF, and NRAS in primary thyroid cancer and metastatic disease. J. Clin. Endocrinol. Metab. 2017, 102, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, P.; Batista, R.; Pestana, A.; Reis, M.; Mendes, A.; Eloy, C.; Sobrinho-Simoes, M.; Soares, P. TERT promoter mutations: A genetic signature of benign and malignant thyroid tumours occurring in the context of tinea capitis irradiation. Eur. J. Endocrinol. 2017, 176, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Chindris, A.M.; Casler, J.D.; Bernet, V.J.; Rivera, M.; Thomas, C.; Kachergus, J.M.; Necela, B.M.; Hay, I.D.; Westphal, S.A.; Grant, C.S.; et al. Clinical and molecular features of Hurthle cell carcinoma of the thyroid. J. Clin. Endocrinol. Metab. 2015, 100, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, R.; Qu, S.; Zhu, G.; Bishop, J.; Liu, X.; Sun, H.; Shan, Z.; Wang, E.; Luo, Y.; et al. Association of TERT promoter mutation 1,295,228 C>T with BRAFV600E mutation, older patient age, and distant metastasis in anaplastic thyroid cancer. J. Clin. Endocrinol. Metab. 2015, 100, E632–E637. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Kondo, T.; Ebina, A.; Sato, Y.; Akaishi, J.; Hino, R.; Yamamoto, N.; Mochizuki, K.; Nakazawa, T.; Yokomichi, H.; et al. Molecular alterations of coexisting thyroid papillary carcinoma and anaplastic carcinoma: Identification of TERT mutation as an independent risk factor for transformation. Mod. Pathol. 2017, 30, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, J.; Li, X.; Liang, Z.; Gao, W.; Liang, J.; Cheng, S.; Lin, Y. TERT promoter mutation predicts radioiodine-refractory character in distant metastatic differentiated thyroid cancer. J. Nucl. Med. 2017, 58, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Brown, T.C.; Juhlin, C.C.; Andreasson, A.; Wang, N.; Backdahl, M.; Healy, J.M.; Prasad, M.L.; Korah, R.; Carling, T.; et al. The activating TERT promoter mutation C228T is recurrent in subsets of adrenal tumors. Endocr. Relat. Cancer 2014, 21, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.S.; de Castro, M.; Calado, R.T. Absence of TERT promoter mutations in pituitary adenomas. J. Endocrinol. Investig. 2016, 39, 933–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Qu, S.; Liu, R.Y.; Sheng, C.J.; Shi, X.G.; Zhu, G.W.; Murugan, A.K.; Guan, H.X.; Yu, H.Y.; Wang, Y.G.; et al. TERT Promoter Mutations and Their Association with BRAFV600E Mutation and Aggressive Clinicopathological Characteristics of Thyroid Cancer. J. Clin. Endocrinol. Metab. 2014, 99, E1130–E1136. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, Y.; Jeon, S.; Bae, J.S.; Jung, S.L.; Jung, C.K. Cytologic, clinicopathologic, and molecular features of papillary thyroid carcinoma with prominent hobnail features: 10 case reports and systematic literature review. Int. J. Clin. Exp. Pathol. 2015, 8, 7988–7997. [Google Scholar] [PubMed]
- De Biase, D.; Gandolfi, G.; Ragazzi, M.; Eszlinger, M.; Sancisi, V.; Gugnoni, M.; Visani, M.; Pession, A.; Casadei, G.; Durante, C.; et al. TERT promoter mutations in papillary thyroid microcarcinomas. Thyroid 2015, 25, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, T.; Matsuse, M.; Hirokawa, M.; Yamashita, S.; Mitsutake, N.; Miyauchi, A. TERT promoter mutations were not found in papillary thyroid microcarcinomas that showed disease progression on active surveillance. Thyroid 2017, 27, 1206–1207. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, A.S.; Qasem, E.; Murugan, A.K.; Al-Hindi, H.N.; AlKhafaji, D.; Almohanna, M.; Xing, M.; Alhomaidah, D.; AlSwailem, M. Uncommon TERT promoter mutations in pediatric thyroid cancer. Thyroid 2016, 26, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Onder, S.; Ozturk, S.S.; Yegen, G.; Sormaz, I.C.; Yilmaz, I.; Poyrazoglu, S.; Sanli, Y.; Senyurek, Y.G.; Kapran, Y.; Mete, O. Classic architecture with multicentricity and local recurrence, and absence of TERT Promoter mutations are correlates of BRAFV600E harboring pediatric papillary thyroid carcinomas. Endocr. Pathol. 2016, 27, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.C.; Ayhan, A.; Maeda, D.; Kim, K.R.; Clarke, B.A.; Shaw, P.; Chui, M.H.; Rosen, B.; Shih, I.M.; Wang, T.L. Frequent somatic mutations of the telomerase reverse transcriptase promoter in ovarian clear cell carcinoma but not in other major types of gynaecological malignancy. J. Pathol. 2014, 232, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Chiang, Y.C.; Cheng, W.F.; Chen, C.A.; Lin, M.C.; Kuo, K.T. Molecular alterations in endometrial and ovarian clear cell carcinomas: Clinical impacts of telomerase reverse transcriptase promoter mutation. Mod. Pathol. 2015, 28, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.A.; Kurtis, B.; Babayeva, S.; Zhuge, J.; Tantchou, I.; Cai, D.; Lafaro, R.J.; Fallon, J.T.; Zhong, M. Heterogeneity of TERT promoter mutations status in squamous cell carcinomas of different anatomical sites. Ann. Diagn. Pathol. 2015, 19, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Vinothkumar, V.; Arunkumar, G.; Revathidevi, S.; Arun, K.; Manikandan, M.; Kuha, A.; Rao, M.; Rajkumar, K.S.; Ajay, C.; Rajaraman, R. TERT promoter hot spot mutations are frequent in Indian cervical and oral squamous cell carcinomas. Tumor Biol. 2016, 37, 7907–7913. [Google Scholar] [CrossRef] [PubMed]
- Dias-Santagata, D.; Selim, M.A.; Su, Y.; Peng, Y.; Vollmer, R.; Chlopik, A.; Tell-Marti, G.; Paral, K.M.; Shalin, S.C.; Shea, C.R.; et al. KIT mutations and CD117 overexpression are markers of better progression-free survival in vulvar melanomas. Br. J. Dermatol. 2017, 177, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Dang, S.; Wu, K.; Shao, Y.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations predict worse survival in laryngeal cancer patients. Int. J. Cancer 2014, 135, 1008–1010. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Wang, C.I.; Pickering, C.R.; Huang, Y.; Tsai, C.N.; Tsang, N.M.; Kao, H.K.; Cheng, M.H.; Myers, J.N. Prevalence of promoter mutations in the TERT gene in oral cavity squamous cell carcinoma. Head Neck 2017, 39, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, D.; Sohn, J.H.; Kim, Y.H.; Lee, J.H.; Lee, H. TERT Promoter Mutation and Telomere Length in Salivary Gland Tumors. Pathol. Oncol. Res. 2017. [Google Scholar] [CrossRef]
- Lam, G.; Xian, R.R.; Li, Y.; Burns, K.H.; Beemon, K.L. Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma. Genes 2016, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Panero, J.; Alves-Paiva, R.M.; Roisman, A.; Santana-Lemos, B.A.; Falcao, R.P.; Oliveira, G.; Martins, D.; Stanganelli, C.; Slavutsky, I.; Calado, R.T. Acquired TERT promoter mutations stimulate TERT transcription in mantle cell lymphoma. Am. J. Hematol. 2016, 91, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Mosrati, M.A.; Willander, K.; Falk, I.J.; Hermanson, M.; Hoglund, M.; Stockelberg, D.; Wei, Y.; Lotfi, K.; Soderkvist, P. Association between TERT promoter polymorphisms and acute myeloid leukemia risk and prognosis. Oncotarget 2015, 6, 25109–25120. [Google Scholar] [CrossRef] [PubMed]
- Tallet, A.; Nault, J.C.; Renier, A.; Hysi, I.; Galateau-Salle, F.; Cazes, A.; Copin, M.C.; Hofman, P.; Andujar, P.; Le Pimpec-Barthes, F.; et al. Overexpression and promoter mutation of the TERT gene in malignant pleural mesothelioma. Oncogene 2014, 33, 3748–3752. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gong, R.; Wang, R.; Pan, Y.; Cai, D.; Pan, B.; Li, Y.; Xiang, J.; Li, H.; Zhang, J.; et al. Recurrent TERT promoter mutations in non-small cell lung cancers. Lung Cancer 2014, 86, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Cao, J.L.; Abuduwufuer, A.; Wang, L.M.; Yuan, X.S.; Lv, W.; Hu, J. Clinical Characteristics and Prognostic Significance of TERT Promoter Mutations in Cancer: A Cohort Study and a Meta-Analysis. PLoS ONE 2016, 11, e0146803. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hao, L.; Li, Y.; Wang, S.; Chen, H.; Zhang, L.; Ke, B.; Yin, Y.; Suo, H.; Sun, B.; et al. Prognostic value analysis of mutational and clinicopathological factors in non-small cell lung cancer. PLoS ONE 2014, 9, e107276. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.J.; Kim, D.S.; Park, W.J.; Lee, H.; Choi, I.J.; Park, J.Y.; Lee, J.H. Mutation of the TERT promoter leads to poor prognosis of patients with non-small cell lung cancer. Oncol. Lett. 2017, 14, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhuge, J.; Bezerra, S.M.; Faraj, S.F.; Munari, E.; Fallon, J.T.; Yang, X.J.; Argani, P.; Netto, G.J.; Zhong, M. High frequency of TERT promoter mutation in small cell carcinoma of bladder, but not in small cell carcinoma of other origins. J. Hematol. Oncol. 2014, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Lindner, S.; Bachmann, H.S.; Odersky, A.; Schaefers, S.; Klein-Hitpass, L.; Hero, B.; Fischer, M.; Eggert, A.; Schramm, A.; Schulte, J.H. Absence of telomerase reverse transcriptase promoter mutations in neuroblastoma. Biomed. Rep. 2015, 3, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Schimming, T.; Moller, I.; Moll, I.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Schadendorf, D.; et al. TERT promoter mutations are frequent in cutaneous basal cell carcinoma and squamous cell carcinoma. PLoS ONE 2013, 8, e80354. [Google Scholar] [CrossRef] [PubMed]
- Populo, H.; Boaventura, P.; Vinagre, J.; Batista, R.; Mendes, A.; Caldas, R.; Pardal, J.; Azevedo, F.; Honavar, M.; Guimaraes, I.; et al. TERT promoter mutations in skin cancer: The effects of sun exposure and X-irradiation. J. Investig. Dermatol. 2014, 134, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.A.; Laughlin, T.S.; Rothberg, P.G. Mutations of the TERT promoter are common in basal cell carcinoma and squamous cell carcinoma. Mod. Pathol. 2014, 27, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Liao, S.L.; Hong, J.B.; Chu, C.Y.; Sheen, Y.S.; Jhuang, J.Y.; Tsai, J.H.; Liau, J.Y. TERT promoter mutations in periocular carcinomas: Implications of ultraviolet light in pathogenesis. Br. J. Ophthalmol. 2016, 100, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Macerola, E.; Loggini, B.; Giannini, R.; Garavello, G.; Giordano, M.; Proietti, A.; Niccoli, C.; Basolo, F.; Fontanini, G. Coexistence of TERT promoter and BRAF mutations in cutaneous melanoma is associated with more clinicopathological features of aggressiveness. Virchows Arch. 2015, 467, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Nagore, E.; Heidenreich, B.; Rachakonda, S.; Garcia-Casado, Z.; Requena, C.; Soriano, V.; Frank, C.; Traves, V.; Quecedo, E.; Sanjuan-Gimenez, J.; et al. TERT promoter mutations in melanoma survival. Int. J. Cancer 2016, 139, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Jeng, Y.M.; Chu, C.Y.; Kuo, K.T.; Liang, C.W. TERT promoter mutation is uncommon in acral lentiginous melanoma. J. Cutan. Pathol. 2014, 41, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, V.d.L.; Vicente, A.L.; Carloni, A.; Berardinelli, G.; Soares, P.; Scapulatempo, C.; Martinho, O.; Reis, R.M. Molecular profiling, including TERT promoter mutations, of acral lentiginous melanomas. Melanoma Res. 2016, 26, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Kong, Y.; Chi, Z.; Sheng, X.; Cui, C.; Wang, X.; Mao, L.; Tang, B.; Li, S.; Lian, B.; et al. MAPK pathway and tert promoter gene mutation pattern and its prognostic value in melanoma patients: A retrospective study of 2793 cases. Clin. Cancer Res. 2017, 23, 6120–6127. [Google Scholar] [CrossRef] [PubMed]
- Roh, M.R.; Park, K.H.; Chung, K.Y.; Shin, S.J.; Rha, S.Y.; Tsao, H. Telomerase reverse transcriptase (TERT) promoter mutations in Korean melanoma patients. Am. J. Cancer Res. 2017, 7, 134–138. [Google Scholar] [PubMed]
- Yang, S.; Leone, D.; Frydenlund, N.; Hoang, M.; Deng, A.; Hernandez-Perez, M.; Biswas, A.; Singh, R.; Yaar, R.; Mahalingam, M. Frequency of telomerase reverse transcripter promoter mutations in desmoplastic melanoma subtypes: Analyses of 76 cases. Melanoma Res. 2016, 26, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Shain, A.H.; Garrido, M.; Botton, T.; Talevich, E.; Yeh, I.; Sanborn, J.Z.; Chung, J.; Wang, N.J.; Kakavand, H.; Mann, G.J.; et al. Exome sequencing of desmoplastic melanoma identifies recurrent NFKBIE promoter mutations and diverse activating mutations in the MAPK pathway. Nat. Genet. 2015, 47, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Egberts, F.; Kruger, S.; Behrens, H.M.; Bergner, I.; Papaspyrou, G.; Werner, J.A.; Alkatout, I.; Haag, J.; Hauschild, A.; Rocken, C. Melanomas of unknown primary frequently harbor TERT-promoter mutations. Melanoma Res. 2014, 24, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Barnhill, R.L.; Dummer, R.; Dalton, J.; Wu, J.; Pappo, A.; Bahrami, A. TERT Promoter Mutations Are Predictive of Aggressive Clinical Behavior in Patients with Spitzoid Melanocytic Neoplasms. Sci. Rep. 2015, 5, 11200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egberts, F.; Bohne, A.S.; Kruger, S.; Hedderich, J.; Rompel, R.; Haag, J.; Rocken, C.; Hauschild, A. Varying Mutational Alterations in Multiple Primary Melanomas. J. Mol. Diagn. 2016, 18, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Ekedahl, H.; Lauss, M.; Olsson, H.; Griewank, K.G.; Schadendorf, D.; Ingvar, C.; Jonsson, G. High TERT promoter mutation frequency in non-acral cutaneous metastatic melanoma. Pigment Cell Melanoma Res. 2016, 29, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Nagore, E.; Heidenreich, B.; Requena, C.; Garcia-Casado, Z.; Martorell-Calatayud, A.; Pont-Sanjuan, V.; Jimenez-Sanchez, A.I.; Kumar, R. TERT promoter mutations associate with fast-growing melanoma. Pigment Cell Melanoma Res. 2016, 29, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Ofner, R.; Ritter, C.; Heidenreich, B.; Kumar, R.; Ugurel, S.; Schrama, D.; Becker, J.C. Distribution of TERT promoter mutations in primary and metastatic melanomas in Austrian patients. J. Cancer Res. Clin. Oncol. 2017, 143, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Schilling, B.; Murali, R.; Bielefeld, N.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Hillen, U.; Schaller, J.; Brenn, T.; et al. TERT promoter mutations are frequent in atypical fibroxanthomas and pleomorphic dermal sarcomas. Mod. Pathol. 2014, 27, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Liu, T.; Wang, N.; Bjornhagen, V.; Hoog, A.; Larsson, C.; Lui, W.O.; Xu, D. TERT promoter mutations and gene amplification: Promoting TERT expression in Merkel cell carcinoma. Oncotarget 2014, 5, 10048–10057. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Yang, S.; Succaria, F.; Bhawan, J. Telomerase (TERT) promoter mutations are infrequent in Merkel cell carcinomas. J. Am. Acad. Dermatol. 2015, 73, 864–865. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Renner, M.; Hartmann, W.; Brandt, R.; Lehner, B.; Waldburger, N.; Alldinger, I.; Schmitt, T.; Egerer, G.; Penzel, R.; et al. TERT promoter hotspot mutations are recurrent in myxoid liposarcomas but rare in other soft tissue sarcoma entities. J. Exp. Clin. Cancer Res. 2014, 33, 33. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Lee, S.; Schaefer, I.M.; Boland, J.M.; Patton, K.T.; Pounds, S.; Fletcher, C.D. TERT promoter mutations and prognosis in solitary fibrous tumor. Mod. Pathol. 2016, 29, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Dubbink, H.J.; Bakels, H.; Post, E.; Zwarthoff, E.C.; Verdijk, R.M. TERT promoter mutations and BRAF mutations are rare in sporadic, and TERT promoter mutations are absent in NF1-related malignant peripheral nerve sheath tumors. J. Neuro-Oncol. 2014, 120, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Campanella, N.C.; Celestino, R.; Pestana, A.; Scapulatempo-Neto, C.; de Oliveira, A.T.; Brito, M.J.; Gouveia, A.; Lopes, J.M.; Guimaraes, D.P.; Soares, P.; et al. Low frequency of TERT promoter mutations in gastrointestinal stromal tumors (GISTs). Eur. J. Hum. Genet. 2015, 23, 877–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; Sitaram, R.T.; Ljungberg, B.; Roos, G.; Hemminki, K.; Kumar, R. TERT promoter mutations in clear cell renal cell carcinoma. Int. J. Cancer 2015, 136, 2448–2452. [Google Scholar] [CrossRef] [PubMed]
- Casuscelli, J.; Becerra, M.F.; Manley, B.J.; Zabor, E.C.; Reznik, E.; Redzematovic, A.; Arcila, M.E.; Tennenbaum, D.M.; Ghanaat, M.; Kashan, M.; et al. Characterization and Impact of TERT Promoter Region Mutations on Clinical Outcome in Renal Cell Carcinoma. Eur. Urol. Focus 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, L.; Liu, J.; Liu, C.; Wang, C.; Ge, N.; Ren, H.; Yan, K.; Hu, S.; et al. TERT promoter mutations in renal cell carcinomas and upper tract urothelial carcinomas. Oncotarget 2014, 5, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, P.; Li, C.; Huang, Y.; Li, X.; Wang, Y.; Chen, C.; Lv, Z.; Tang, A.; Sun, X.; et al. Telomerase reverse transcriptase gene promoter mutations help discern the origin of urogenital tumors: A genomic and molecular study. Eur. Urol. 2014, 65, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Allory, Y.; Beukers, W.; Sagrera, A.; Flandez, M.; Marques, M.; Marquez, M.; van der Keur, K.A.; Dyrskjot, L.; Lurkin, I.; Vermeij, M.; et al. Telomerase reverse transcriptase promoter mutations in bladder cancer: High frequency across stages, detection in urine, and lack of association with outcome. Eur. Urol. 2014, 65, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurst, C.D.; Platt, F.M.; Knowles, M.A. Comprehensive mutation analysis of the TERT promoter in bladder cancer and detection of mutations in voided urine. Eur. Urol. 2014, 65, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, P.S.; Hosen, I.; de Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, C.; Meng, Y.; Yuan, X.; Liu, L.; Ge, N.; Liu, J.; Wang, C.; Ren, H. TERT promoter mutations and TERT mRNA but not FGFR3 mutations are urinary biomarkers in Han Chinese patients with urothelial bladder cancer. Oncologist 2015, 20, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Descotes, F.; Kara, N.; Decaussin-Petrucci, M.; Piaton, E.; Geiguer, F.; Rodriguez-Lafrasse, C.; Terrier, J.E.; Lopez, J.; Ruffion, A. Non-invasive prediction of recurrence in bladder cancer by detecting somatic TERT promoter mutations in urine. Br. J. Cancer 2017, 117, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Liu, C.; Wang, K.; Liu, L.; Liu, T.T.; Ge, N.; Kong, F.; Yang, L.; Bjorkholm, M.; Fan, Y.D.; et al. The genetic difference between Western and Chinese urothelial cell carcinomas: Infrequent FGFR3 mutation in Han Chinese patients. Oncotarget 2016, 7, 25826–25835. [Google Scholar] [CrossRef] [PubMed]
- Pietzak, E.J.; Bagrodia, A.; Cha, E.K.; Drill, E.N.; Iyer, G.; Isharwal, S.; Ostrovnaya, I.; Baez, P.; Li, Q.; Berger, M.F.; et al. Next-generation sequencing of nonmuscle invasive bladder cancer reveals potential biomarkers and rational therapeutic targets. Eur. Urol. 2017, 72, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Kinde, I.; Munari, E.; Faraj, S.F.; Hruban, R.H.; Schoenberg, M.; Bivalacqua, T.; Allaf, M.; Springer, S.; Wang, Y.; Diaz, L.A., Jr.; et al. TERT promoter mutations occur early in urothelial neoplasia and are biomarkers of early disease and disease recurrence in urine. Cancer Res. 2013, 73, 7162–7167. [Google Scholar] [CrossRef] [PubMed]
- Vail, E.; Zheng, X.; Zhou, M.; Yang, X.; Fallon, J.T.; Epstein, J.I.; Zhong, M. Telomerase reverse transcriptase promoter mutations in glandular lesions of the urinary bladder. Ann. Diagn. Pathol. 2015, 19, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Lew, M.; Weigelin, H.C.; Weizer, A.Z.; Montgomery, J.S.; Betz, B.L.; Mehra, R. Comparative study of TERT promoter mutation status within spatially, temporally and morphologically distinct components of urothelial carcinoma. Histopathology 2018, 72, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.; Springer, S.; Nguyen, D.; Taheri, D.; Guner, G.; Rodriguez, M.A.; Wang, Y.; Kinde, I.; VandenBussche, C.J.; Olson, M.T.; et al. High prevalence of TERT promoter mutations in primary squamous cell carcinoma of the urinary bladder. Mod. Pathol. 2016, 29, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.L.; Springer, S.; Nguyen, D.; Taheri, D.; Guner, G.; Mendoza Rodriguez, M.A.; Wang, Y.; Kinde, I.; Del Carmen Rodriguez Pena, M.; VandenBussche, C.J.; et al. Detection of TERT promoter mutations in primary adenocarcinoma of the urinary bladder. Hum. Pathol. 2016, 53, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Pena, M.D.C.; Tregnago, A.C.; Eich, M.L.; Springer, S.; Wang, Y.; Taheri, D.; Ertoy, D.; Fujita, K.; Bezerra, S.M.; Cunha, I.W.; et al. Spectrum of genetic mutations in de novo PUNLMP of the urinary bladder. Virchows Arch. 2017, 471, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.T.; Ge, N.; Liu, L.; Yuan, X.T.; Liu, J.K.; Kong, F.; Wang, C.; Ren, H.B.; Yan, K.Q.; et al. TERT promoter mutations are associated with distant metastases in upper tract urothelial carcinomas and serve as urinary biomarkers detected by a sensitive castPCR. Oncotarget 2014, 5, 12428–12439. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lopez-Beltran, A.; Osunkoya, A.O.; Wang, M.; Zhang, S.; Davidson, D.D.; Emerson, R.E.; Williamson, S.R.; Tan, P.H.; Kaimakliotis, H.Z.; et al. TERT promoter mutation status in sarcomatoid urothelial carcinomas of the upper urinary tract. Future Oncol. 2017, 13, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Taheri, D.; Springer, S.; Cowan, M.; Guner, G.; Mendoza Rodriguez, M.A.; Wang, Y.; Kinde, I.; VandenBussche, C.J.; Olson, M.T.; et al. High prevalence of TERT promoter mutations in micropapillary urothelial carcinoma. Virchows Arch. 2016, 469, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kurtis, B.; Zhuge, J.; Ojaimi, C.; Ye, F.; Cai, D.; Zhang, D.; Fallon, J.T.; Zhong, M. Recurrent TERT promoter mutations in urothelial carcinoma and potential clinical applications. Ann. Diagn. Pathol. 2016, 21, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Priemer, D.S.; Wang, M.; Zhang, S.; Lopez-Beltran, A.; Kouba, E.; Montironi, R.; Davidson, D.D.; MacLennan, G.T.; Wang, L.; Osunkoya, A.O.; et al. Small-cell carcinomas of the urinary bladder and prostate: TERT promoter mutation status differentiates sites of malignancy and provides evidence of common clonality between small-cell carcinoma of the urinary bladder and urothelial carcinoma. Eur. Urol. Focus 2017. [Google Scholar] [CrossRef] [PubMed]
- Lack, E.E. Tumours of the adrenal glands and extraadrenal paraganglia; American Registry of Pathology in collaboration with the Armed Forces Institute of Pathology: Washington, DC, USA, 2007. [Google Scholar]
- Heidenreich, B.; Nagore, E.; Rachakonda, P.S.; Garcia-Casado, Z.; Requena, C.; Traves, V.; Becker, J.; Soufir, N.; Hemminki, K.; Kumar, R. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat. Commun. 2014, 5, 3401. [Google Scholar] [CrossRef] [PubMed]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; de Verdier, P.J.; Ryk, C.; Steineck, G.; Hemminki, K.; Kumar, R. Mutations in TERT promoter and FGFR3 and telomere length in bladder cancer. Int. J. Cancer 2015, 137, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Piscuoglio, S.; Ng, C.K.Y.; Murray, M.; Burke, K.A.; Edelweiss, M.; Geyer, F.C.; Macedo, G.S.; Inagaki, A.; Papanastasiou, A.D.; Martelotto, L.G.; et al. Massively parallel sequencing of phyllodes tumours of the breast reveals actionable mutations, and TERT promoter hotspot mutations and TERT gene amplification as likely drivers of progression. J. Pathol. 2016, 238, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, Y.; Kratz, J.; Brat, D.J.; Robitaille, Y.; Moghrabi, A.; Perlman, E.J.; Dang, C.V.; Burger, P.C.; Eberhart, C.G. hTERT gene amplification and increased mRNA expression in central nervous system embryonal tumors. Am. J. Pathol. 2003, 162, 1763–1769. [Google Scholar] [CrossRef]
- Palmqvist, R.; Zhang, A.; Xu, D.; Golovleva, I.; Norrback, K.F.; Gruber, A.; Oberg, A.; Stenling, R.; Roos, G. hTERT gene copy number is not associated with hTERT RNA expression or telomerase activity in colorectal cancer. Int. J. Cancer 2005, 116, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Takuma, Y.; Nouso, K.; Kobayashi, Y.; Nakamura, S.; Tanaka, H.; Matsumoto, E.; Fujikawa, T.; Suzuki, M.; Hanafusa, T.; Shiratori, Y. Telomerase reverse transcriptase gene amplification in hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2004, 19, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Visnovsky, J.; Kudela, E.; Farkasova, A.; Balharek, T.; Krkoska, M.; Danko, J. Amplification of TERT and TERC genes in cervical intraepithelial neoplasia and cervical cancer. Neuroendocrinol. Lett. 2014, 35, 518–522. [Google Scholar] [PubMed]
- Freier, K.; Pungs, S.; Flechtenmacher, C.; Bosch, F.X.; Lichter, P.; Joos, S.; Hofele, C. Frequent high telomerase reverse transcriptase expression in primary oral squamous cell carcinoma. J. Oral. Pathol. Med. 2007, 36, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Cutz, J.C.; Liu, N.; Lau, D.; Shepherd, F.A.; Squire, J.A.; Tsao, M.S. Amplification of telomerase (hTERT) gene is a poor prognostic marker in non-small-cell lung cancer. Br. J. Cancer 2006, 94, 1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Ludovini, V.; Trere, D.; Siggillino, A.; Vannucci, J.; Bellezza, G.; Crino, L.; Montanaro, L. Dyskerin and TERC expression may condition survival in lung cancer patients. Oncotarget 2015, 6, 21755–21760. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Puig-Butille, J.A.; Munoz, C.; Costa, D.; Diez, A.; Garcia-Herrera, A.; Carrera, C.; Badenas, C.; Sole, F.; Malvehy, J.; Puig, S.; Alos, L. TERT gene amplification is associated with poor outcome in acral lentiginous melanoma. J. Am. Acad. Dermatol. 2014, 71, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Puig-Butillé, J.A.; Badenas, C.; Ogbah, Z.; Carrera, C.; Aguilera, P.; Malvehy, J.; Puig, S. Genetic alterations in RAS-regulated pathway in acral lentiginous melanoma. Exp. Dermatol. 2013, 22, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Opresko, P.; Pappo, A.; Kirkwood, J.M.; Bahrami, A. Association of TERT promoter mutations with telomerase expression in melanoma. Pigment Cell Melanoma Res. 2016, 29, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Chochi, Y.; Matsuyama, H.; Eguchi, S.; Kawauchi, S.; Furuya, T.; Oga, A.; Kang, J.J.; Naito, K.; Sasaki, K. Gain of 5p15.33 is associated with progression of bladder cancer. Oncology 2007, 72, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Sugita, M.; Tanaka, N.; Davidson, S.; Sekiya, S.; Varella-Garcia, M.; West, J.; Drabkin, H.A.; Gemmill, R.M. Molecular definition of a small amplification domain within 3q26 in tumors of cervix, ovary, and lung. Cancer Genet. Cytogenet. 2000, 117, 9–18. [Google Scholar] [CrossRef]
- Liu, H.Q.; Liu, S.L.; Wang, H.; Xie, X.Y.; Chen, X.L.; Zhang, X.M.; Zhang, Y.C. Genomic amplification of the human telomerase gene (hTERC) associated with human papillomavirus is related to the progression of uterine cervical dysplasia to invasive cancer. Diagn. Pathol. 2012, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Sowjanya, P.; Wangsa, D.; Hjerpe, A.; Johansson, B.; Auer, G.; Gravitt, P.E.; Larsson, C.; Wallin, K.L.; Ried, T.; et al. Detection of genomic amplification of the human telomerase gene TERC, a potential marker for triage of women with HPV-positive, abnormal Pap smears. Am. J. Pathol. 2009, 175, 1831–1847. [Google Scholar] [CrossRef] [PubMed]
- Soder, A.I.; Hoare, S.F.; Muir, S.; Going, J.J.; Parkinson, E.K.; Keith, W.N. Amplification, increased dosage and in situ expression of the telomerase RNA gene in human cancer. Oncogene 1997, 14, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-C.; Shyong, W.-Y.; Chang, M.-S.; Chen, Y.-J.; Lin, C.-H.; Huang, Z.-D.; Hsu, M.-T.; Chen, M.-L. Frequent gain of copy number on the long arm of chromosome 3 in human cervical adenocarcinoma. Cancer Genet. Cytogenet. 2001, 131, 48–53. [Google Scholar] [CrossRef]
- Andersson, S.; Wallin, K.L.; Hellstrom, A.C.; Morrison, L.E.; Hjerpe, A.; Auer, G.; Ried, T.; Larsson, C.; Heselmeyer-Haddad, K. Frequent gain of the human telomerase gene TERC at 3q26 in cervical adenocarcinomas. Br. J. Cancer 2006, 95, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Xu, L.; Shu, Y.; Tian, T.; Liang, J.; Xu, Y.; Wang, F.; Chen, J.; Dai, J.; Hu, Z.; et al. Common genetic variants on 5p15.33 contribute to risk of lung adenocarcinoma in a Chinese population. Carcinogenesis 2009, 30, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.T.; Chatterjee, N.; Yu, K.; Goldin, L.R.; Goldstein, A.M.; Rotunno, M.; Mirabello, L.; Jacobs, K.; Wheeler, W.; Yeager, M.; et al. A genome-wide association study of lung cancer identifies a region of chromosome 5p15 associated with risk for adenocarcinoma. Am. J. Hum. Genet. 2009, 85, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Shete, S.; Hosking, F.J.; Robertson, L.B.; Dobbins, S.E.; Sanson, M.; Malmer, B.; Simon, M.; Marie, Y.; Boisselier, B.; Delattre, J.Y.; et al. Genome-wide association study identifies five susceptibility loci for glioma. Nat. Genet. 2009, 41, 899–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Broderick, P.; Matakidou, A.; Eisen, T.; Houlston, R.S. Role of 5p15.33 (TERT-CLPTM1L), 6p21.33 and 15q25.1 (CHRNA5-CHRNA3) variation and lung cancer risk in never-smokers. Carcinogenesis 2010, 31, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, C.A.; Lan, Q.; Hong, Y.C.; Chen, C.J.; Hosgood, H.D.; Chang, I.S.; Chatterjee, N.; Brennan, P.; Wu, C.; Zheng, W.; et al. The 5p15.33 locus is associated with risk of lung adenocarcinoma in never-smoking females in Asia. PLoS Genet. 2010, 6, e1001051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, D.; Kubo, M.; Takahashi, A.; Yoon, K.A.; Kim, J.; Lee, G.K.; Zo, J.I.; Lee, J.S.; Hosono, N.; Morizono, T.; et al. Variation in TP63 is associated with lung adenocarcinoma susceptibility in Japanese and Korean populations. Nat. Genet. 2010, 42, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, Y.; Zhao, Y.; Fan, W.; Zhou, K.; Liu, Y.; Zhou, L.; Mao, Y.; Wei, Q.; Xu, J.; et al. Association of sequence variants on chromosomes 20, 11, and 5 (20q13.33, 11q23.3, and 5p15.33) with glioma susceptibility in a Chinese population. Am. J. Epidemiol. 2011, 173, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.Y.; Lee, S.Y.; Kang, B.K.; Lee, E.J.; Choi, Y.Y.; Kang, H.G.; Choi, J.E.; Jeon, H.S.; Lee, W.K.; Kam, S. Replication of results of genome-wide association studies on lung cancer susceptibility loci in a Korean population. Respirology 2012, 17, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Cai, S.; Chen, Q.G.; Ni, Z.H.; Tang, J.H.; Xu, D.W.; Wang, X.B. Multiple variants of TERT and CLPTM1L constitute risk factors for lung adenocarcinoma. Genet. Mol. Res. 2012, 11, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.L.; Enciso-Mora, V.; Marie, Y.; Desestret, V.; Labussière, M.; Boisselier, B.; Mokhtari, K.; Idbaih, A.; Hoang-Xuan, K.; Delattre, J.-Y. Association between glioma susceptibility loci and tumour pathology defines specific molecular etiologies. Neuro-Oncology 2012, 15, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Kinnersley, B.; Migliorini, G.; Broderick, P.; Whiffin, N.; Dobbins, S.E.; Casey, G.; Hopper, J.; Sieber, O.; Lipton, L.; Kerr, D.J.; et al. The TERT variant rs2736100 is associated with colorectal cancer risk. Br. J. Cancer 2012, 107, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, J.; Hu, L.; Ding, C.; Kan, Y.; Shen, Y.; Chen, X.; Shen, H.; Guo, X.; Hu, Z. Common genetic variants in TERT contribute to risk of cervical cancer in a Chinese population. Mol. Carcinog. 2012, 51 (Suppl. 1), E118–E122. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Cawthon, R.; Gao, Y.; Hu, W.; Hosgood, H.D., 3rd; Barone-Adesi, F.; Ji, B.T.; Bassig, B.; Chow, W.H.; Shu, X.; et al. Longer telomere length in peripheral white blood cells is associated with risk of lung cancer and the rs2736100 (CLPTM1L-TERT) polymorphism in a prospective cohort study among women in China. PLoS ONE 2013, 8, e59230. [Google Scholar] [CrossRef] [PubMed]
- Myneni, A.A.; Chang, S.C.; Niu, R.; Liu, L.; Ochs-Balcom, H.M.; Li, Y.; Zhang, C.; Zhao, B.; Shi, J.; Han, X.; et al. Genetic polymorphisms of TERT and CLPTM1L and risk of lung cancer—A case-control study in a Chinese population. Lung Cancer 2013, 80, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Safaeian, M.; Rajaraman, P.; Hartge, P.; Yeager, M.; Linet, M.; Butler, M.A.; Ruder, A.M.; Purdue, M.P.; Hsing, A.; Beane-Freeman, L.; et al. Joint effects between five identified risk variants, allergy, and autoimmune conditions on glioma risk. Cancer Causes Control 2013, 24, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Tong, N.; Tao, G.; Luo, D.; Wang, M.; Fang, Y.; Li, J.; Xu, M.; Zhang, Z.; Wu, D. TERT polymorphisms modify the risk of acute lymphoblastic leukemia in Chinese children. Carcinogenesis 2013, 34, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Jager, R.; Harutyunyan, A.S.; Rumi, E.; Pietra, D.; Berg, T.; Olcaydu, D.; Houlston, R.S.; Cazzola, M.; Kralovics, R. Common germline variation at the TERT locus contributes to familial clustering of myeloproliferative neoplasms. Am. J. Hematol. 2014, 89, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Oddsson, A.; Kristinsson, S.Y.; Helgason, H.; Gudbjartsson, D.F.; Masson, G.; Sigurdsson, A.; Jonasdottir, A.; Jonasdottir, A.; Steingrimsdottir, H.; Vidarsson, B.; et al. The germline sequence variant rs2736100_C in TERT associates with myeloproliferative neoplasms. Leukemia 2014, 28, 1371–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.M.; Zhang, Y.; Shen, L.; Ren, Y.W.; Li, X.L.; Yin, Z.H.; Zhou, B.S. Genetic variations in TERT-CLPTM1L genes and risk of lung cancer in a Chinese population. Asian Pac. J. Cancer Prev. 2014, 15, 2809–2813. [Google Scholar] [CrossRef] [PubMed]
- Campa, D.; Rizzato, C.; Stolzenberg-Solomon, R.; Pacetti, P.; Vodicka, P.; Cleary, S.P.; Capurso, G.; Bueno-de-Mesquita, H.B.; Werner, J.; Gazouli, M.; et al. TERT gene harbors multiple variants associated with pancreatic cancer susceptibility. Int. J. Cancer 2015, 137, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Krahling, T.; Balassa, K.; Kiss, K.P.; Bors, A.; Batai, A.; Halm, G.; Egyed, M.; Fekete, S.; Remenyi, P.; Masszi, T. Co-occurrence of myeloproliferative neoplasms and solid tumors is attributed to a synergism between cytoreductive therapy and the common TERT polymorphism rs2736100. Cancer Epidemiol. Prev. Biomark. 2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.G.; Ma, L.; Cen, Q.H.; Huang, J.S.; Zhang, J.X.; Zhang, J.J. Association of genetic polymorphisms in TERT-CLPTM1L with lung cancer in a Chinese population. Genet. Mol. Res. 2015, 14, 4469–4476. [Google Scholar] [CrossRef] [PubMed]
- Tapper, W.; Jones, A.V.; Kralovics, R.; Harutyunyan, A.S.; Zoi, K.; Leung, W.; Godfrey, A.L.; Guglielmelli, P.; Callaway, A.; Ward, D.; et al. Genetic variation at MECOM, TERT, JAK2 and HBS1L-MYB predisposes to myeloproliferative neoplasms. Nat. Commun. 2015, 6, 6691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Cao, L.; Pu, H.; Wang, H.; Zheng, Y.; Niu, X.; Weng, X.; Zhang, H.; Favus, M.; Zhang, L.; et al. TERT Polymorphism rs2736100-C is associated with EGFR mutation-positive non-small cell lung cancer. Clin. Cancer Res. 2015, 21, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Bayram, S.; Ulger, Y.; Sumbul, A.T.; Kaya, B.Y.; Genc, A.; Rencuzogullari, E.; Dadas, E. Polymorphisms in human telomerase reverse transcriptase (hTERT) gene and susceptibility to gastric cancer in a Turkish population: Hospital-based case-control study. Gene 2016, 585, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Carkic, J.; Nikolic, N.; Radojevic-Skodric, S.; Kuzmanovic-Pficer, J.; Brajovic, G.; Antunovic, M.; Milasin, J.; Popovic, B. The role of TERT-CLPTM1L SNPs, hTERT expression and telomere length in the pathogenesis of oral squamous cell carcinoma. J. Oral. Sci 2016, 58, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Dahlstrom, J.; Liu, T.; Yuan, X.; Saft, L.; Ghaderi, M.; Wei, Y.B.; Lavebratt, C.; Li, P.; Zheng, C.; Bjorkholm, M.; et al. TERT rs2736100 genotypes are associated with differential risk of myeloproliferative neoplasms in Swedish and Chinese male patient populations. Ann. Hematol. 2016, 95, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Shi, M.; An, C.; Yang, W.; Nie, X.; Zhang, J.; Lv, Z.; Li, J.; Zhou, L.; Du, Z.; et al. Functional evaluation of TERT-CLPTM1L genetic variants associated with susceptibility of papillary thyroid carcinoma. Sci. Rep. 2016, 6, 26037. [Google Scholar] [CrossRef] [PubMed]
- Trifa, A.P.; Bănescu, C.; Tevet, M.; Bojan, A.; Dima, D.; Urian, L.; Török-Vistai, T.; Popov, V.M.; Zdrenghea, M.; Petrov, L. TERT rs2736100 A> C SNP and JAK2 46/1 haplotype significantly contribute to the occurrence of JAK2V617F and CALR mutated myeloproliferative neoplasms—A multicentric study on 529 patients. Br. J. Haematol. 2016, 174, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Prescott, J.; Yuan, C.; Zhang, M.; Kraft, P.; Babic, A.; Morales-Oyarvide, V.; Qian, Z.R.; Buring, J.E.; Cochrane, B.B.; et al. Leucocyte telomere length, genetic variants at the TERT gene region and risk of pancreatic cancer. Gut 2017, 66, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Thakur, A.; Liang, Y.; Zhang, S.; Wang, T.; Chen, T.; Meng, J.; Wang, L.; Wu, F.; Jin, T.; et al. Polymorphisms in the TERT gene are associated with lung cancer risk in the Chinese Han population. Eur. J. Cancer Prev. 2014, 23, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Su, L.Y.; Li, X.L.; Shen, L.; Zhang, Y.; Zhao, M.M.; Yin, Z.H.; Su, H.Y.; Zhou, B.S. Polymorphisms of TERT and CLPTM1L and the risk of hepatocellular carcinoma in Chinese males. Asian Pac. J. Cancer Prev. 2014, 15, 8197–8201. [Google Scholar] [CrossRef] [PubMed]
- Jannuzzi, A.T.; Karaman, E.; Oztas, E.; Yanar, H.T.; Ozhan, G. Telomerase Reverse Transcriptase (TERT) Gene Variations and Susceptibility of Colorectal Cancer. Genet. Test. Mol. Biomark. 2015, 19, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.S.; Do, S.K.; Choi, J.E.; Lee, S.Y.; Lee, J.; Cha, S.I.; Kim, C.H.; Park, J.Y. TERT Polymorphism rs2853669 Influences on Lung Cancer Risk in the Korean Population. J. Korean Med. Sci. 2015, 30, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Oztas, E.; Kara, H.; Kara, Z.P.; Aydogan, M.U.; Uras, C.; Ozhan, G. Association Between Human Telomerase Reverse Transcriptase Gene Variations and Risk of Developing Breast Cancer. Genet. Test. Mol. Biomark. 2016, 20, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Zhang, H.; Zhai, Y.; Wang, Z.; Li, P.; Yu, L.; Xia, X.; Zhang, Y.; Zeng, Y.; et al. Common variations in TERT-CLPTM1L locus are reproducibly associated with the risk of nasopharyngeal carcinoma in Chinese populations. Oncotarget 2016, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Egan, K.M.; Thompson, R.C.; Nabors, L.; Olson, J.J.; Brat, D.J.; LaRocca, R.V.; Brem, S.; Moots, P.L.; Madden, M.H.; Browning, J.E. Cancer susceptibility variants and the risk of adult glioma in a US case-control study. J. Neuro-Oncol. 2011, 104, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Nan, H.; Qureshi, A.A.; Prescott, J.; de Vivo, I.; Han, J. Genetic variants in telomere-maintaining genes and skin cancer risk. Hum. Genet. 2011, 129, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jin, T.B.; Wei, X.B.; He, S.M.; Liang, H.J.; Yang, H.X.; Cui, Y.; Chen, C.; Cai, L.B.; Gao, G.D. Selected polymorphisms of GSTP1 and TERT were associated with glioma risk in Han Chinese. Cancer Epidemiol. 2012, 36, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Terry, K.L.; Tworoger, S.S.; Vitonis, A.F.; Wong, J.; Titus-Ernstoff, L.; De Vivo, I.; Cramer, D.W. Telomere length and genetic variation in telomere maintenance genes in relation to ovarian cancer risk. Cancer Epidemiol. Biomark. Prev. 2012, 21, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.B.; Zhang, J.Y.; Li, G.; Du, S.L.; Geng, T.T.; Gao, J.; Liu, Q.P.; Gao, G.D.; Kang, L.L.; Chen, C.; et al. RTEL1 and TERT polymorphisms are associated with astrocytoma risk in the Chinese Han population. Tumor Biol. 2013, 34, 3659–3666. [Google Scholar] [CrossRef] [PubMed]
- Kote-Jarai, Z.; Saunders, E.J.; Leongamornlert, D.A.; Tymrakiewicz, M.; Dadaev, T.; Jugurnauth-Little, S.; Ross-Adams, H.; Al Olama, A.A.; Benlloch, S.; Halim, S.; et al. Fine-mapping identifies multiple prostate cancer risk loci at 5p15, one of which associates with TERT expression. Hum. Mol. Genet. 2013, 22, 2520–2528. [Google Scholar] [CrossRef] [PubMed]
- Pellatt, A.J.; Wolff, R.K.; Torres-Mejia, G.; John, E.M.; Herrick, J.S.; Lundgreen, A.; Baumgartner, K.B.; Giuliano, A.R.; Hines, L.M.; Fejerman, L. Telomere length, telomere-related genes, and breast cancer risk: The breast cancer health disparities study. Genes Chromosomes Cancer 2013, 52, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, C.; Yang, L.; Zhang, X.; Zhao, X.; Song, X.; Li, X.; Wang, J.; Qian, J.; Yang, Y.; et al. Significant association of 5p15.33 (TERT-CLPTM1L genes) with lung cancer in Chinese Han population. Exp. Lung Res. 2013, 39, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Fesinmeyer, M.D.; Timofeeva, M.; Caberto, C.P.; Kocarnik, J.M.; Han, Y.; Love, S.A.; Young, A.; Dumitrescu, L.; Lin, Y.; et al. Pleiotropic associations of risk variants identified for other cancers with lung cancer risk: The PAGE and TRICL consortia. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Cao, W.; Wang, L.; Liu, S.; Liu, Z.; Zhang, B.; Yang, H.; Feng, T.; Zhang, J.; Zhang, X.; et al. Genetic variants in TERT are associated with risk of gastric cancer in a Chinese Han population. Oncotarget 2016, 7, 82727–82732. [Google Scholar] [CrossRef] [PubMed]
- Savage, S.A.; Chanock, S.J.; Lissowska, J.; Brinton, L.A.; Richesson, D.; Peplonska, B.; Bardin-Mikolajczak, A.; Zatonski, W.; Szeszenia-Dabrowska, N.; Garcia-Closas, M. Genetic variation in five genes important in telomere biology and risk for breast cancer. Br. J. Cancer 2007, 97, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Gammon, M.D.; Wu, H.C.; Terry, M.B.; Wang, Q.; Bradshaw, P.T.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M. Multiple genetic variants in telomere pathway genes and breast cancer risk. Cancer Epidemiol. Biomark. Prev. 2010, 19, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Liu, L.; Zou, L.; Zhu, Y.; Chen, W.; Zhu, B.; Shen, N.; Rui, R.; Long, L.; Ke, J. Genetic variations in TERT-CLPTM1L locus are associated with risk of lung cancer in chinese population. Mol. Carcinog. 2013, 52 (Suppl. 1), 118–126. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, A.S.; Boyarskikh, U.A.; Oskina, N.A.; Sinkina, T.V.; Lazarev, A.F.; Petrova, V.D.; Filipenko, M.L. TERT polymorphisms rs2853669 and rs7726159 influence on prostate cancer risk in Russian population. Tumor Biol. 2015, 36, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Seo, H.W.; Jung, E.S.; Kim, B.H.; Jung, G. The TERT promoter SNP rs2853669 decreases E2F1 transcription factor binding and increases mortality and recurrence risks in liver cancer. Oncotarget 2016, 7, 684–699. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Maturo, M.G.; Martorelli, C.; Suppa, M.; Antonini, A.; Kostaki, D.; Verna, L.; Landi, M.T.; Peris, K.; Fargnoli, M.C. Characterization of melanoma susceptibility genes in high-risk patients from Central Italy. Melanoma Res. 2017, 27, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Kang, H.G.; Jang, J.S.; Choi, Y.Y.; Kim, M.J.; Kim, J.S.; Jeon, H.S.; Lee, W.K.; Cha, S.I.; Kim, C.H.; et al. Polymorphisms in telomere maintenance genes and risk of lung cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, F.J.; Vizcaino, M.A.; Blakeley, J.; Heaphy, C.M. Frequent alternative lengthening of telomeres and ATRX loss in adult NF1-associated diffuse and high-grade astrocytomas. Acta Neuropathol. 2016, 132, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Abedalthagafi, M.; Phillips, J.J.; Kim, G.E.; Mueller, S.; Haas-Kogen, D.A.; Marshall, R.E.; Croul, S.E.; Santi, M.R.; Cheng, J.; Zhou, S.M.; et al. The alternative lengthening of telomere phenotype is significantly associated with loss of ATRX expression in high-grade pediatric and adult astrocytomas: A multi-institutional study of 214 astrocytomas. Mod. Pathol. 2013, 26, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Hakin-Smith, V.; Teo, M.; Xinarianos, G.E.; Jellinek, D.A.; Carroll, T.; McDowell, D.; MacFarlane, M.R.; Boet, R.; Baguley, B.C.; et al. Association of mutant TP53 with alternative lengthening of telomeres and favorable prognosis in glioma. Cancer Res. 2006, 66, 6473–6476. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Heaphy, C.M.; Wilde, R.F.; Orr, B.A.; Odia, Y.; Eberhart, C.G.; Meeker, A.K.; Rodriguez, F.J. Molecular and Morphologic Correlates of the Alternative Lengthening of Telomeres Phenotype in High-Grade Astrocytomas. Brain Pathol. 2013, 23, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Hannay, J.A.; McCarthy, S.W.; Royds, J.A.; Yeager, T.R.; Robinson, R.A.; Wharton, S.B.; Jellinek, D.A.; Arbuckle, S.M.; Yoo, J.Y.; et al. A robust assay for alternative lengthening of telomeres in tumors shows the significance of alternative lengthening of telomeres in sarcomas and astrocytomas. Clin. Cancer Res. 2005, 11, 217–225. [Google Scholar] [PubMed]
- Mangerel, J.; Price, A.; Castelo-Branco, P.; Brzezinski, J.; Buczkowicz, P.; Rakopoulos, P.; Merino, D.; Baskin, B.; Wasserman, J.; Mistry, M.; et al. Alternative lengthening of telomeres is enriched in, and impacts survival of TP53 mutant pediatric malignant brain tumors. Acta Neuropathol. 2014, 128, 853–862. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.L.; McDonnell, J.; Muntoni, A.; Henson, J.D.; Hegi, M.E.; von Deimling, A.; Wheeler, H.R.; Cook, R.J.; Biggs, M.T.; Little, N.S.; et al. Presence of alternative lengthening of telomeres mechanism in patients with glioblastoma identifies a less aggressive tumor type with longer survival. J. Neuropathol. Exp. Neurol. 2010, 69, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Fogli, A.; Demattei, M.V.; Corset, L.; Vaurs-Barriere, C.; Chautard, E.; Biau, J.; Kemeny, J.L.; Godfraind, C.; Pereira, B.; Khalil, T.; et al. Detection of the alternative lengthening of telomeres pathway in malignant gliomas for improved molecular diagnosis. J. Neuro-Oncol. 2017, 135, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Hakin-Smith, V.; Jellinek, D.A.; Levy, D.; Carroll, T.; Teo, M.; Timperley, W.R.; McKay, M.J.; Reddel, R.R.; Royds, J.A. Alternative lengthening of telomeres and survival in patients with glioblastoma multiforme. Lancet 2003, 361, 836–838. [Google Scholar] [CrossRef]
- Wood, L.D.; Heaphy, C.M.; Daniel, H.D.; Naini, B.V.; Lassman, C.R.; Arroyo, M.R.; Kamel, I.R.; Cosgrove, D.P.; Boitnott, J.K.; Meeker, A.K.; et al. Chromophobe hepatocellular carcinoma with abrupt anaplasia: A proposal for a new subtype of hepatocellular carcinoma with unique morphological and molecular features. Mod. Pathol. 2013, 26, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Omori, Y.; Nakayama, F.; Li, D.; Kanemitsu, K.; Semba, S.; Ito, A.; Yokozaki, H. Alternative lengthening of telomeres frequently occurs in mismatch repair system-deficient gastric carcinoma. Cancer Sci. 2009, 100, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Dogeas, E.; Karagkounis, G.; Heaphy, C.M.; Hirose, K.; Pawlik, T.M.; Wolfgang, C.L.; Meeker, A.; Hruban, R.H.; Cameron, J.L.; Choti, M.A. Alternative lengthening of telomeres predicts site of origin in neuroendocrine tumor liver metastases. J. Am. Coll. Surg. 2014, 218, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xu, D.; Sofiadis, A.; Hoog, A.; Vukojevic, V.; Backdahl, M.; Zedenius, J.; Larsson, C. Telomerase-dependent and independent telomere maintenance and its clinical implications in medullary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2014, 99, E1571–E1579. [Google Scholar] [CrossRef] [PubMed]
- Else, T.; Giordano, T.J.; Hammer, G.D. Evaluation of telomere length maintenance mechanisms in adrenocortical carcinoma. J. Clin. Endocrinol. Metab. 2008, 93, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, I.; Kurrer, A.S.; Vassella, E.; Dettmer, M.; Rudolph, T.; Banz, V.; Hunger, F.; Pasquinelli, S.; Speel, E.J.; Perren, A. Loss of DAXX and ATRX are associated with chromosome instability and reduced survival of patients with pancreatic neuroendocrine tumors. Gastroenterology 2014, 146, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, H.S.; Nam, K.H.; Choi, J.; Kim, W.H. Telomere length abnormalities and telomerase RNA component expression in gastroenteropancreatic neuroendocrine tumors. Anticancer Res. 2015, 35, 3501–3510. [Google Scholar] [PubMed]
- Singhi, A.D.; Liu, T.C.; Roncaioli, J.L.; Cao, D.; Zeh, H.J.; Zureikat, A.H.; Tsung, A.; Marsh, J.W.; Lee, K.K.; Hogg, M.E.; et al. Alternative Lengthening of Telomeres and Loss of DAXX/ATRX Expression Predicts Metastatic Disease and Poor Survival in Patients with Pancreatic Neuroendocrine Tumors. Clin. Cancer Res. 2017, 23, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Brosnan-Cashman, J.A.; An, S.; Kim, S.J.; Song, K.B.; Kim, M.S.; Kim, M.J.; Hwang, D.W.; Meeker, A.K.; Yu, E.; et al. Alternative Lengthening of Telomeres in Primary Pancreatic Neuroendocrine Tumors Is Associated with Aggressive Clinical Behavior and Poor Survival. Clin. Cancer Res. 2017, 23, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- VandenBussche, C.J.; Allison, D.B.; Graham, M.K.; Charu, V.; Lennon, A.M.; Wolfgang, C.L.; Hruban, R.H.; Heaphy, C.M. Alternative lengthening of telomeres and ATRX/DAXX loss can be reliably detected in FNAs of pancreatic neuroendocrine tumors. Cancer Cytopathol. 2017, 125, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; de Wilde, R.F.; Jiao, Y.; Klein, A.P.; Edil, B.H.; Shi, C.; Bettegowda, C.; Rodriguez, F.J.; Eberhart, C.G.; Hebbar, S.; et al. Altered telomeres in tumors with ATRX and DAXX mutations. Science 2011, 333, 425. [Google Scholar] [CrossRef] [PubMed]
- Villa, R.; Daidone, M.G.; Motta, R.; Venturini, L.; De Marco, C.; Vannelli, A.; Kusamura, S.; Baratti, D.; Deraco, M.; Costa, A.; et al. Multiple mechanisms of telomere maintenance exist and differentially affect clinical outcome in diffuse malignant peritoneal mesothelioma. Clin. Cancer Res. 2008, 14, 4134–4140. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-K.; Park, N.-H.; Lee, H. Prognostic Value of Alternative Lengthening of Telomeres-Associated Biomarkers in Uterine Sarcoma and Uterine Carcinosarcoma. Int. J. Gynecol. Cancer 2012, 22, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Pezzolo, A.; Pistorio, A.; Gambini, C.; Haupt, R.; Ferraro, M.; Erminio, G.; De Bernardi, B.; Garaventa, A.; Pistoia, V. Intratumoral diversity of telomere length in individual neuroblastoma tumors. Oncotarget 2015, 6, 7493–7503. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E.; Varkonyi, R.J.; Schwalm, J.; Cragle, R.; Klein-Szanto, A.; Patchefsky, A.; Cukierman, E.; von Mehren, M.; Broccoli, D. Multiple mechanisms of telomere maintenance exist in liposarcomas. Clin. Cancer Res. 2005, 11, 5347–5355. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Daidone, M.G.; Daprai, L.; Villa, R.; Cantu, S.; Pilotti, S.; Mariani, L.; Gronchi, A.; Henson, J.D.; Reddel, R.R.; et al. Telomere maintenance mechanisms in liposarcomas: Association with histologic subtypes and disease progression. Cancer Res. 2006, 66, 8918–8924. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Motta, R.; Gronchi, A.; Daidone, M.; Zaffaroni, N. Prognostic relevance of ALT-associated markers in liposarcoma: A comparative analysis. BMC Cancer 2010, 10, 254. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Jeng, Y.M.; Liau, J.Y.; Tsai, J.H.; Hsu, H.H.; Yang, C.Y. Alternative lengthening of telomeres and loss of ATRX are frequent events in pleomorphic and dedifferentiated liposarcomas. Mod. Pathol. 2015, 28, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Lee, J.C.; Tsai, J.H.; Yang, C.Y.; Liu, T.L.; Ke, Z.L.; Hsu, H.H.; Jeng, Y.M. Comprehensive screening of alternative lengthening of telomeres phenotype and loss of ATRX expression in sarcomas. Mod. Pathol. 2015, 28, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Jeng, Y.M.; Lee, J.C.; Hsu, H.H.; Yang, C.Y. Leiomyosarcoma with Alternative Lengthening of Telomeres Is Associated with Aggressive Histologic Features, Loss of ATRX Expression, and Poor Clinical Outcome. Am. J. Surg. Pathol. 2015, 39, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ohali, A.; Avigad, S.; Naumov, I.; Goshen, Y.; Ash, S.; Yaniv, I. Different telomere maintenance mechanisms in alveolar and embryonal rhabdomyosarcoma. Genes Chromosomes Cancer 2008, 47, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Yang, C.Y.; Lee, J.C.; Liang, C.W.; Hsu, H.H.; Jeng, Y.M. Alternative lengthening of telomeres phenotype in malignant vascular tumors is highly associated with loss of ATRX expression and is frequently observed in hepatic angiosarcomas. Hum. Pathol. 2015, 46, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Daidone, M.G.; Motta, R.; Cimino-Reale, G.; Hoare, S.F.; Gronchi, A.; Folini, M.; Keith, W.N.; Zaffaroni, N. Telomere maintenance mechanisms in malignant peripheral nerve sheath tumors: Expression and prognostic relevance. Neuro-Oncology 2012, 14, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Ulaner, G.A.; Huang, H.Y.; Otero, J.; Zhao, Z.Q.; Ben-Porat, L.; Satagopan, J.M.; Gorlick, R.; Meyers, P.; Healey, J.H.; Huvos, A.G.; et al. Absence of a telomere maintenance mechanism as a favorable prognostic factor in patients with osteosarcoma. Cancer Res. 2003, 63, 1759–1763. [Google Scholar] [PubMed]
- Sanders, R.P.; Drissi, R.; Billups, C.A.; Daw, N.C.; Valentine, M.B.; Dome, J.S. Telomerase expression predicts unfavorable outcome in osteosarcoma. J. Clin. Oncol. 2004, 22, 3790–3797. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Shay, J.W.; Wright, W.E.; Hiyama, E.; Shimose, S.; Kubo, T.; Sugita, T.; Yasunaga, Y.; Ochi, M. Telomere-maintenance mechanisms in soft-tissue malignant fibrous histiocytomas. J. Bone Joint Surg. Am. 2009, 91, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Venturini, L.; Daidone, M.G.; Motta, R.; Collini, P.; Spreafico, F.; Terenziani, M.; Piva, L.; Radice, P.; Perotti, D.; Zaffaroni, N. Telomere maintenance in Wilms tumors: First evidence for the presence of alternative lengthening of telomeres mechanism. Genes Chromosomes Cancer 2011, 50, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Calderaro, J.; Di Tommaso, L.; Balabaud, C.; Zafrani, E.S.; Bioulac-Sage, P.; Roncalli, M.; Zucman-Rossi, J. Telomerase reverse transcriptase promoter mutation is an early somatic genetic alteration in the transformation of premalignant nodules in hepatocellular carcinoma on cirrhosis. Hepatology 2014, 60, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Pinyol, R.; Tovar, V.; Llovet, J.M. TERT promoter mutations: Gatekeeper and driver of hepatocellular carcinoma. J. Hepatol. 2014, 61, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Kim, Y.; Jeon, S.; Kim, S.H.; Kim, T.J.; Lee, S.; Kim, M.H.; Lim, D.J.; Lee, Y.S.; Jung, C.K. Clinical utility of TERT promoter mutations and ALK rearrangement in thyroid cancer patients with a high prevalence of the BRAFV600E mutation. Diagn. Pathol. 2016, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, A.; Chang, D.K.; Nones, K.; Corbo, V.; Patch, A.M.; Bailey, P.; Lawlor, R.T.; Johns, A.L.; Miller, D.K.; Mafficini, A.; et al. Whole-genome landscape of pancreatic neuroendocrine tumours. Nature 2017, 543, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Tian, D.; Chen, Q.; Zhang, S.; Liu, B.; Ji, B. Telomerase reverse transcriptase (TERT) rs2736100 polymorphism contributes to increased risk of glioma: Evidence from a meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 422–430. [Google Scholar] [PubMed]
- Cao, J.L.; Yuan, P.; Abuduwufuer, A.; Lv, W.; Yang, Y.H.; Hu, J. Association between the TERT Genetic Polymorphism rs2853676 and Cancer Risk: Meta-Analysis of 76,108 Cases and 134,215 Controls. PLoS ONE 2015, 10, e0128829. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.Y.; Jones, D.T.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.A.; Tonjes, M.; et al. Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Choufani, S.; Mack, S.; Gallagher, D.; Zhang, C.; Lipman, T.; Zhukova, N.; Walker, E.J.; Martin, D.; Merino, D.; et al. Methylation of the TERT promoter and risk stratification of childhood brain tumours: An integrative genomic and molecular study. Lancet Oncol. 2013, 14, 534–542. [Google Scholar] [CrossRef]
- Hertwig, F.; Peifer, M.; Fischer, M. Telomere maintenance is pivotal for high-risk neuroblastoma. Cell Cycle 2016, 15, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Kojima, M.; Ueda, Y.; Kurihara, S.; Hiyama, E. Telomere biology including TERT rearrangements in neuroblastoma: A useful indicator for surgical treatments. J. Pediatr. Surg. 2016, 51, 2080–2085. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.F.; Zhao, Q. TERT-mediated and ATRX-mediated Telomere Maintenance and Neuroblastoma. J. Pediatr. Hematol. Oncol. 2018, 40, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Onitake, Y.; Hiyama, E.; Kamei, N.; Yamaoka, H.; Sueda, T.; Hiyama, K. Telomere biology in neuroblastoma: Telomere binding proteins and alternative strengthening of telomeres. J.Pediatr. Surg. 2009, 44, 2258–2266. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Yamada, T. Molecular biology of the ETS family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Borah, S.; Xi, L.; Zaug, A.J.; Powell, N.M.; Dancik, G.M.; Cohen, S.B.; Costello, J.C.; Theodorescu, D.; Cech, T.R. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science 2015, 347, 1006–1010. [Google Scholar]
- Xi, L.H.; Schmidt, J.C.; Zaug, A.J.; Ascarrunz, D.R.; Cech, T.R. A novel two-step genome editing strategy with CRISPR-Cas9 provides new insights into telomerase action and TERT gene expression. Genome Biol. 2015, 16, 231. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hemminki, K.; Kumar, R. TERT promoter mutations in cancer development. Curr. Opin. Genet. Dev. 2014, 24, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Labussiere, M.; Boisselier, B.; Mokhtari, K.; Di Stefano, A.L.; Rahimian, A.; Rossetto, M.; Ciccarino, P.; Saulnier, O.; Paterra, R.; Marie, Y.; et al. Combined analysis of TERT, EGFR, and IDH status defines distinct prognostic glioblastoma classes. Neurology 2014, 83, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qi, C.; Maling, G.; Xiang, W.; Yanhui, L.; Ruofei, L.; Yunhe, M.; Jiewen, L.; Qing, M. TERT mutation in glioma: Frequency, prognosis and risk. J. Clin. Neurosci. 2016, 26, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Zucman-Rossi, J. TERT promoter mutations in primary liver tumors. Clin. Res. Hepatol. Gastroenterol. 2016, 40, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Jeng, Y.M.; Chang, C.N.; Lee, H.J.; Hsu, H.C.; Lai, P.L.; Yuan, R.H. TERT promoter mutation in resectable hepatocellular carcinomas: A strong association with hepatitis C infection and absence of hepatitis B infection. Int. J. Surg. 2014, 12, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, F.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Frequency and geographic distribution of TERT promoter mutations in primary hepatocellular carcinoma. Infect. Agents Cancer 2017, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Sobrinho-Simões, M.; Soares, P. Coexistence of TERT promoter and BRAF mutations in papillary thyroid carcinoma: Added value in patient prognosis? J. Clin. Oncol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Stoehr, R.; Taubert, H.; Zinnall, U.; Giedl, J.; Gaisa, N.T.; Burger, M.; Ruemmele, P.; Hurst, C.D.; Knowles, M.A.; Wullich, B.; et al. Frequency of TERT Promoter Mutations in Prostate Cancer. Pathobiology 2015, 82, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Lorbeer, F.K.; Shain, A.H.; McSwiggen, D.T.; Schruf, E.; Oh, A.; Ryu, J.; Darzacq, X.; Bastian, B.C.; Hockemeyer, D. Mutations in the promoter of the telomerase gene TERT contribute to tumorigenesis by a two-step mechanism. Science 2017, 357, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The cancer genome. Nature 2009, 458, 719. [Google Scholar] [CrossRef] [PubMed]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-M.; Xi, R.; Luquette, L.J.; Park, R.W.; Johnson, M.D.; Park, P.J. Functional genomic analysis of chromosomal aberrations in a compendium of 8000 cancer genomes. Genome Res. 2013, 23, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhang, C.-Z.; Wala, J.; Mermel, C.H. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134. [Google Scholar] [CrossRef] [PubMed]
- Bryce, L.A.; Morrisont, N.; Hoare, S.F.; Muir, S.; Keith, W.N. Mapping of the gene for the human telomerase reverse transcriptase, hTERT, to chromosome 5p15. 33 by fluorescence in situ hybridization. Neoplasia 2000, 2, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Rooney, P.H.; Murray, G.I.; Stevenson, D.A.; Haites, N.E.; Cassidy, J.; McLeod, H.L. Comparative genomic hybridization and chromosomal instability in solid tumours. Br. J. Cancer 1999, 80, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Santarius, T.; Shipley, J.; Brewer, D.; Stratton, M.R.; Cooper, C.S. A census of amplified and overexpressed human cancer genes. Nat. Rev. Cancer 2010, 10, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Knuutila, S.; Bjorkqvist, A.M.; Autio, K.; Tarkkanen, M.; Wolf, M.; Monni, O.; Szymanska, J.; Larramendy, M.L.; Tapper, J.; Pere, H.; et al. DNA copy number amplifications in human neoplasms: Review of comparative genomic hybridization studies. Am. J. Pathol. 1998, 152, 1107–1123. [Google Scholar] [PubMed]
- Iwabuchi, H.; Sakamoto, M.; Sakunaga, H.; Ma, Y.-Y.; Carcangiu, M.L.; Pinkel, D.; Yang-Feng, T.L.; Gray, J.W. Genetic analysis of benign, low-grade, and high-grade ovarian tumors. Cancer Res. 1995, 55, 6172–6180. [Google Scholar] [PubMed]
- Arnold, N.; Hägele, L.; Walz, L.; Schempp, W.; Pfisterer, J.; Bauknecht, T.; Kiechle, M. Overrepresentation of 3q and 8q material and loss of 18q material are recurrent findings in advanced human ovarian cancer. Genes Chromosomes Cancer 1996, 16, 46–54. [Google Scholar] [CrossRef]
- Balsara, B.R.; Sonoda, G.; Du Manoir, S.; Siegfried, J.M.; Gabrielson, E.; Testa, J.R. Comparative genomic hybridization analysis detects frequent, often high-level, overrepresentation of DNA sequences at 3q, 5p, 7p, and 8q in human non-small cell lung carcinomas. Cancer Res. 1997, 57, 2116–2120. [Google Scholar] [PubMed]
- Heselmeyer, K.; Schröck, E.; Du Manoir, S.; Blegen, H.; Shah, K.; Steinbeck, R.; Auer, G.; Ried, T. Gain of chromosome 3q defines the transition from severe dysplasia to invasive carcinoma of the uterine cervix. Proc. Natl. Acad. Sci. USA 1996, 93, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Heselmeyer-Haddad, K.; Macville, M.; Schröck, E.; Blegen, H.; Hellström, A.-C.; Shah, K.; Auer, G.; Ried, T. Advanced-stage cervical carcinomas are defined by a recurrent pattern of chromosomal aberrations revealing high genetic instability and a consistent gain of chromosome arm 3q. Genes Chromosom. Cancer 1997, 19, 233–240. [Google Scholar] [CrossRef]
- Ried, T.; Petersen, I.; Holtgreve-Grez, H.; Speicher, M.R.; Schröck, E.; du Manoir, S.; Cremer, T. Mapping of multiple DNA gains and losses in primary small cell lung carcinomas by comparative genomic hybridization. Cancer Res. 1994, 54, 1801–1806. [Google Scholar] [PubMed]
- Levin, N.A.; Brzoska, P.; Gupta, N.; Minna, J.D.; Gray, J.W.; Christman, M.F. Identification of frequent novel genetic alterations in small cell lung carcinoma. Cancer Res. 1994, 54, 5086–5091. [Google Scholar] [PubMed]
- Levin, N.A.; Bnorka, P.M.; Warnock, M.L.; Gray, J.W.; Christman, M.F. Identification of novel regions of altered DNA copy number in small cell lung tumors. Genes Chromosomes Cancer 1995, 13, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Brass, N.; Ukena, I.; Remberger, K.; Mack, U.; Sybrecht, G.; Meese, E. DNA amplification on chromosome 3q26.1-q26.3 in squamous cell carcinoma of the lung detected by reverse chromosome painting. Eur. J. Cancer 1996, 32, 1205–1208. [Google Scholar] [CrossRef]
- Brzoska, P.M.; Levin, N.A.; Fu, K.K.; Kaplan, M.J.; Singer, M.I.; Gray, J.W.; Christman, M.F. Frequent novel DNA copy number increase in squamous cell head and neck tumors. Cancer Res. 1995, 55, 3055–3059. [Google Scholar] [PubMed]
- Speicher, M.R.; Howe, C.; Crotty, P.; du Manoir, S.; Costa, J.; Ward, D.C. Comparative genomic hybridization detects novel deletions and amplifications in head and neck squamous cell carcinomas. Cancer Res. 1995, 55, 1010–1013. [Google Scholar] [PubMed]
- Bockmühl, U.; Schwendel, A.; Dietel, M.; Petersen, I. Distinct patterns of chromosomal alterations in high-and low-grade head and neck squamous cell carcinomas. Cancer Res. 1996, 56, 5325–5329. [Google Scholar] [PubMed]
- Dierlamm, J.; Rosenberg, C.; Stul, M.; Pittaluga, S.; Wlodarska, I.; Michaux, L.; Dehaen, M.; Verhoef, G.; Thomas, J.; De Kelver, W. Characteristic pattern of chromosomal gains and losses in marginal zone B cell lymphoma detected by comparative genomic hybridization. Leukemia 1997, 11, 747. [Google Scholar] [CrossRef] [PubMed]
- Björkqvist, A.-M.; Husgafvel-Pursiainen, K.; Anttila, S.; Karjalainen, A.; Tammilehto, L.; Mattson, K.; Vainio, H.; Knuutila, S. DNA gains in 3q occur frequently in squamous cell carcinoma of the lung, but not in adenocarcinoma. Genes Chromosomes Cancer 1998, 22, 79–82. [Google Scholar] [CrossRef]
- Tarkkanen, M.; Elomaa, I.; Blomqvist, C.; Kivioja, A.H.; Kellokumpu-Lehtinen, P.; Bohling, T.; Valle, J.; Knuutila, S. DNA sequence copy number increase at 8q: A potential new prognostic marker in high-grade osteosarcoma. Int. J. Cancer 1999, 84, 114–121. [Google Scholar] [CrossRef]
- Larramendy, M.L.; Tarkkanen, M.; Blomqvist, C.; Virolainen, M.; Wiklund, T.; Asko-Seljavaara, S.; Elomaa, I.; Knuutila, S. Comparative genomic hybridization of malignant fibrous histiocytoma reveals a novel prognostic marker. Am. J. Pathol. 1997, 151, 1153–1161. [Google Scholar] [PubMed]
- El-Rifai, W.; Sarlomo-Rikala, M.; Miettinen, M.; Knuutila, S.; Andersson, L.C. DNA copy number losses in chromosome 14: An early change in gastrointestinal stromal tumors. Cancer Res. 1996, 56, 3230–3233. [Google Scholar] [PubMed]
- Zhang, A.; Zheng, C.; Hou, M.; Lindvall, C.; Wallin, K.L.; Ångström, T.; Yang, X.; Hellström, A.C.; Blennow, E.; Björkholm, M. Amplification of the telomerase reverse transcriptase (hTERT) gene in cervical carcinomas. Genes Chromosomes Cancer 2002, 34, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Flacco, A.; Ludovini, V.; Bianconi, F.; Ragusa, M.; Bellezza, G.; Tofanetti, F.R.; Pistola, L.; Siggillino, A.; Vannucci, J.; Cagini, L.; et al. MYC and human telomerase gene (TERC) copy number gain in early-stage non-small cell lung cancer. Am. J. Clin. Oncol. 2015, 38, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.T.; Han, W.; Cho, J.; Lee, J.W.; Ko, E.; Kim, E.K.; Jung, S.Y.; Jeong, E.M.; Bae, J.Y.; Kang, J.J.; et al. Genomic copy number alterations as predictive markers of systemic recurrence in breast cancer. Int. J. Cancer 2008, 123, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Heselmeyer-Haddad, K.; Janz, V.; Castle, P.E.; Chaudhri, N.; White, N.; Wilber, K.; Morrison, L.E.; Auer, G.; Burroughs, F.H.; Sherman, M.E.; et al. Detection of genomic amplification of the human telomerase gene (TERC) in cytologic specimens as a genetic test for the diagnosis of cervical dysplasia. Am. J. Pathol. 2003, 163, 1405–1416. [Google Scholar] [CrossRef]
- Heselmeyer-Haddad, K.; Sommerfeld, K.; White, N.M.; Chaudhri, N.; Morrison, L.E.; Palanisamy, N.; Wang, Z.Y.; Auer, G.; Steinberg, W.; Ried, T. Genomic amplification of the human telomerase gene (TERC) in pap smears predicts the development of cervical cancer. Am. J. Pathol. 2005, 166, 1229–1238. [Google Scholar] [CrossRef]
- Baird, D.M. Variation at the TERT locus and predisposition for cancer. Exp. Rev. Mol. Med. 2010, 12, e16. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.D.; Hung, R.J.; Gaborieau, V.; Boffetta, P.; Chabrier, A.; Byrnes, G.; Zaridze, D.; Mukeria, A.; Szeszenia-Dabrowska, N.; Lissowska, J.; et al. Lung cancer susceptibility locus at 5p15.33. Nat. Genet. 2008, 40, 1404–1406. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Zhang, X.Y.; Jin, B. TERT genetic polymorphism rs2736100 was associated with lung cancer: A meta-analysis based on 14,492 subjects. Genet. Test. Mol. Biomark. 2013, 17, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jiao, S. Increased lung cancer risk associated with the TERT rs2736100 polymorphism: An updated meta-analysis. Tumor Biol. 2014, 35, 5763–5769. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.J.; Yoon, J.H.; Kim, O.; Choi, W.S.; Nam, S.W.; Lee, J.Y.; Park, W.S. Influence of the hTERT rs2736100 polymorphism on telomere length in gastric cancer. World J. Gastroenterol. 2015, 21, 9328–9336. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.T.; Meng, Y.; Li, P.; Ge, N.; Kong, F.; Yang, L.; Bjorkholm, M.; Zhao, S.T.; Xu, D.W. The association between the TERT rs2736100 AC genotype and reduced risk of upper tract urothelial carcinomas in a Han Chinese population. Oncotarget 2016, 7, 31972–31979. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yu, H.; Sun, J.; Qi, J.; Liu, Q.; Li, R.; Zheng, S.L.; Xu, J.; Kang, J. Association of genetic polymorphisms in the telomerase reverse transcriptase gene with prostate cancer aggressiveness. Mol. Med. Rep. 2015, 12, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Gu, A.; Ji, G.; Zhao, L.; Zhao, P.; Lu, A. The TERT rs2736100 polymorphism and cancer risk: A meta-analysis based on 25 case-control studies. BMC Cancer 2012, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yin, Z.; Wu, W.; Li, X.; Zhou, B. Genetic variants in TERT-CLPTM1L genetic region associated with several types of cancer: A meta-analysis. Gene 2013, 526, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, G.; Wei, S.; Niu, J.; Wang, L.E.; Sturgis, E.M.; Wei, Q. Genetic variations in TERT-CLPTM1L genes and risk of squamous cell carcinoma of the head and neck. Carcinogenesis 2010, 31, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Qiao, N.; Wang, Y.; Jiang, M.; Wang, S.; Wang, C.; Hu, L. Association between the telomerase reverse transcriptase (TERT) rs2736098 polymorphism and cancer risk: Evidence from a case-control study of non-small-cell lung cancer and a meta-analysis. PLoS ONE 2013, 8, e76372. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; McGrath, M.; Lee, I.M.; Buring, J.E.; De Vivo, I. Telomere length and genetic analyses in population-based studies of endometrial cancer risk. Cancer 2010, 116, 4275–4282. [Google Scholar] [CrossRef] [PubMed]
- Hofer, P.; Baierl, A.; Bernhart, K.; Leeb, G.; Mach, K.; Micksche, M.; Gsur, A. Association of genetic variants of human telomerase with colorectal polyps and colorectal cancer risk. Mol. Carcinog. 2012, 51 (Suppl. 1), E176–E182. [Google Scholar] [CrossRef] [PubMed]
- Pellatt, A.J.; Wolff, R.K.; Herrick, J.; Lundgreen, A.; Slattery, M.L. TERT’s role in colorectal carcinogenesis. Mol. Carcinog. 2013, 52, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.L.; Ogundiran, T.O.; Adebamowo, C.; Nathanson, K.L.; Domchek, S.M.; Rebbeck, T.R.; Simon, M.S.; John, E.M.; Hennis, A.; Nemesure, B.; et al. Lack of association between common single nucleotide polymorphisms in the TERT-CLPTM1L locus and breast cancer in women of African ancestry. Breast Cancer Res. Treat. 2012, 132, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Varadi, V.; Brendle, A.; Grzybowska, E.; Johansson, R.; Enquist, K.; Butkiewicz, D.; Pamula-Pilat, J.; Pekala, W.; Hemminki, K.; Lenner, P.; et al. A functional promoter polymorphism in the TERT gene does not affect inherited susceptibility to breast cancer. Cancer Genet. Cytogenet. 2009, 190, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Lee, S.H.; Kim, J.Y.; Kim, J.E.; Kim, T.M.; Lee, S.T.; Choi, S.H.; Park, S.H.; Kim, I.H. Expression level of hTERT is regulated by somatic mutation and common single nucleotide polymorphism at promoter region in glioblastoma. Oncotarget 2014, 5, 3399–3407. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Lu, Y.J.; Wang, X.; Peng, J.; Zhu, Y.W.; Cheng, L.M. Association between rs2853669 in TERT gene and the risk and prognosis of human cancer: A systematic review and meta-analysis. Oncotarget 2017, 8, 50864–50872. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Rozadilla, C.; Kartsonaki, C.; Woolley, C.; McClellan, M.; Whittington, D.; Horgan, G.; Leedham, S.; Kriaucionis, S.; East, J.; Tomlinson, I. Telomere length and genetics are independent colorectal tumour risk factors in an evaluation of biomarkers in normal bowel. Br. J. Cancer 2018, 118, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.J.; McBride, D.J.; Lin, M.-L.; Varela, I.; Pleasance, E.D.; Simpson, J.T.; Stebbings, L.A.; Leroy, C.; Edkins, S.; Mudie, L.J. Complex landscapes of somatic rearrangement in human breast cancer genomes. Nature 2009, 462, 1005. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, L.J.; Koster, J.; Zwijnenburg, D.A.; Hasselt, N.E.; van Sluis, P.; Volckmann, R.; van Noesel, M.M.; George, R.E.; Tytgat, G.A.; Molenaar, J.J.; et al. TERT rearrangements are frequent in neuroblastoma and identify aggressive tumors. Nat. Genet. 2015, 47, 1411–1414. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.; Alitalo, K.; Klempnauer, K.-H.; Varmus, H.E.; Bishop, J.M.; Gilbert, F.; Brodeur, G.; Goldstein, M.; Trent, J. Amplified DNA with limited homology to myc cellular oncogene is shared by human neuroblastoma cell lines and a neuroblastoma tumour. Nature 1983, 305, 245. [Google Scholar] [CrossRef] [PubMed]
- Poremba, C.; Hero, B.; Heine, B.; Scheel, C.; Schaefer, K.-L.; Christiansen, H.; Berthold, F.; Kneif, S.; Stein, H.; Juergens, H. Telomerase is a strong indicator for assessing the proneness to progression in neuroblastomas. Med. Pediatr. Oncol. 2000, 35, 651–655. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, S.; Popova, E.Y.; Grigoryev, S.A.; Zhu, J. Rearrangement of upstream sequences of the hTERT gene during cellular immortalization. Genes Chromosomes Cancer 2009, 48, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Alexandrov, L.B.; Wedge, D.C.; Van Loo, P.; Greenman, C.D.; Raine, K.; Jones, D.; Hinton, J.; Marshall, J.; Stebbings, L.A. Mutational processes molding the genomes of 21 breast cancers. Cell 2012, 149, 979–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L. Signatures of mutational processes in human cancer. Nature 2013, 500, 415. [Google Scholar] [CrossRef] [PubMed]
- Ohali, A.; Avigad, S.; Ash, S.; Goshen, Y.; Luria, D.; Feinmesser, M.; Zaizov, R.; Yaniv, I. Telomere length is a prognostic factor in neuroblastoma. Cancer 2006, 107, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Jeison, M.; Yaniv, I.; Ash, S. Genetic stratification of neuroblastoma for treatment tailoring. Future Oncol. 2011, 7, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Cahill, D.P.; Louis, D.N.; Cairncross, J.G. Molecular background of oligodendroglioma: 1p/19q, IDH, TERT, CIC and FUBP1. CNS Oncol. 2015, 4, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Rao, C.M. Epigenetics in cancer: Fundamentals and Beyond. Pharmacol. Ther. 2017, 173, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Falus, A.; Marton, I.; Borbényi, E.; Tahy, Á.; Karádi, P.; Aradi, J.; Stauder, A.; Kopp, M. A challenging epigenetic message: Telomerase activity is associated with complex changes in lifestyle. Cell Biol. Int. 2011, 35, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Entringer, S.; Epel, E.S.; Kumsta, R.; Lin, J.; Hellhammer, D.H.; Blackburn, E.H.; Wüst, S.; Wadhwa, P.D. Stress exposure in intrauterine life is associated with shorter telomere length in young adulthood. Proc. Natl. Acad. Sci. USA 2011, 108, E513–E518. [Google Scholar] [CrossRef] [PubMed]
- Price, L.H.; Kao, H.-T.; Burgers, D.E.; Carpenter, L.L.; Tyrka, A.R. Telomeres and early-life stress: An overview. Biol. Psychiatry 2013, 73, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Epel, E.; Blackburn, E. Telomeres and lifestyle factors: Roles in cellular aging. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2012, 730, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Tollefsbol, T.O. Regulation of the Telomerase Reverse Transcriptase Subunit through Epigenetic Mechanisms. Front. Genet. 2016, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic diet: Impact on the epigenome and cancer. Epigenomics 2011, 3, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Takai, D. The role of DNA methylation in mammalian epigenetics. Science 2001, 293, 1068–1070. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Han, Z.; Zhu, R.; Liu, P.; Liu, S. Association of p16 gene methylation with prostate cancer risk: A meta-analysis. J. BUON 2015, 20, 1074–1080. [Google Scholar] [PubMed]
- Herman, J.G.; Umar, A.; Polyak, K.; Graff, J.R.; Ahuja, N.; Issa, J.-P.J.; Markowitz, S.; Willson, J.K.; Hamilton, S.R.; Kinzler, K.W. Incidence and functional consequences of hMLH1 promoter hypermethylation in colorectal carcinoma. Proc. Natl. Acad. Sci. USA 1998, 95, 6870–6875. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, X.; Wang, Y.; Hu, F.; Wang, F.; Jiang, L.; Liu, Y.; Wang, D.; Sun, G.; Zhao, Y. MLH1 promoter methylation frequency in colorectal cancer patients and related clinicopathological and molecular features. PLoS ONE 2013, 8, e59064. [Google Scholar] [CrossRef] [PubMed]
- Szyf, M.; Pakneshan, P.; Rabbani, S.A. DNA methylation and breast cancer. Biochem. Pharmacol. 2004, 68, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Dessain, S.K.; Yu, H.-Y.; Reddel, R.R.; Beijersbergen, R.L.; Weinberg, R.A. Methylation of the human telomerase gene CpG island. Cancer Res. 2000, 60, 537–541. [Google Scholar] [PubMed]
- Guilleret, I.; Benhattar, J. Demethylation of the human telomerase catalytic subunit (hTERT) gene promoter reduced hTERT expression and telomerase activity and shortened telomeres. Exp. Cell Res. 2003, 289, 326–334. [Google Scholar] [CrossRef]
- Guilleret, I.; Yan, P.; Grange, F.; Braunschweig, R.; Bosman, F.T.; Benhattar, J. Hypermethylation of the human telomerase catalytic subunit (hTERT) gene correlates with telomerase activity. Int. J. Cancer 2002, 101, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Gigek, C.O.; Leal, M.F.; Silva, P.N.O.; Lisboa, L.C.F.; Lima, E.M.; Calcagno, D.Q.; Assumpcao, P.P.; Burbano, R.R.; Smith, M.D.A.C. hTERT methylation and expression in gastric cancer. Biomarkers 2009, 14, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, A.; Müller, H.M.; Hubalek, M.M.; Wiedemair, A.; Fiegl, H.; Goebel, G.; Mueller-Holzner, E.; Marth, C.; Widschwendter, M. Methylation status and expression of human telomerase reverse transcriptase in ovarian and cervical cancer. Gynecol. Oncol. 2004, 93, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, P.; Messinis, I.; Tsezou, A. DNA methylation is not likely to be responsible for hTERT expression in premalignant cervical lesions. Exp. Biol. Med. 2007, 232, 881–886. [Google Scholar]
- Fan, Y.; Lee, S.; Wu, G.; Easton, J.; Yergeau, D.; Dummer, R.; Vogel, P.; Kirkwood, J.M.; Barnhill, R.L.; Pappo, A.; et al. Telomerase Expression by Aberrant Methylation of the TERT Promoter in Melanoma Arising in Giant Congenital Nevi. J. Investig. Dermatol. 2016, 136, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; Loukinov, D.; Abdullaev, Z.; Guilleret, I.; Bosman, F.T.; Lobanenkov, V.; Benhattar, J. Dual role of DNA methylation inside and outside of CTCF-binding regions in the transcriptional regulation of the telomerase hTERT gene. Nucleic Acids Res. 2007, 35, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Katzenellenbogen, R.A.; Grandori, C.; Galloway, D.A. An unbiased in vivo screen reveals multiple transcription factors that control HPV E6-regulated hTERT in keratinocytes. Virology 2013, 446, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Valls-Bautista, C.; Bougel, S.; Piñol-Felis, C.; Viñas-Salas, J.; Benhattar, J. hTERT methylation is necessary but not sufficient for telomerase activity in colorectal cells. Oncol. Lett. 2011, 2, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Bechter, O.E.; Eisterer, W.; Dlaska, M.; Kühr, T.; Thaler, J. CpG island methylation of the hTERT promoter is associated with lower telomerase activity in B-cell lymphocytic leukemia. Exp. Hematol. 2002, 30, 26–33. [Google Scholar] [CrossRef]
- Choi, J.-H.; Park, S.H.; Park, J.; Park, B.G.; Cha, S.-J.; Kong, K.-H.; Lee, K.-H.; Park, A.J. Site-specific methylation of CpG nucleotides in the hTERT promoter region can control the expression of hTERT during malignant progression of colorectal carcinoma. Biochem. Biophys. Res. Commun. 2007, 361, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Srinivasan, R.; Vasishta, R.K.; Wig, J.D. Positive regulation of human telomerase reverse transcriptase gene expression and telomerase activity by DNA methylation in pancreatic cancer. Ann. Surg. Oncol. 2009, 16, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Petko, Z.; Dzieciatkowski, S.; Lin, L.; Ghiassi, M.; Stain, S.; Chapman, W.C.; Washington, M.K.; Willis, J.; Markowitz, S.D. CpG island methylation of genes accumulates during the adenoma progression step of the multistep pathogenesis of colorectal cancer. Genes Chromosomes Cancer 2006, 45, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Satra, M.; Drakaki, A.; Poultsides, G.A.; Tsezou, A. Epigenetic regulation of hTERT promoter in hepatocellular carcinomas. Int. J. Oncol. 2009, 34, 391–399. [Google Scholar] [PubMed]
- Castelo-Branco, P.; Leao, R.; Lipman, T.; Campbell, B.; Lee, D.; Price, A.; Zhang, C.; Heidari, A.; Stephens, D.; Boerno, S.; et al. A cancer specific hypermethylation signature of the TERT promoter predicts biochemical relapse in prostate cancer: A retrospective cohort study. Oncotarget 2016, 7, 57726–57736. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, I.; Apolonio, J.D.; Price, A.J.; De Mello, R.A.; Roberto, V.P.; Tabori, U.; Castelo-Branco, P. The TERT hypermethylated oncologic region predicts recurrence and survival in pancreatic cancer. Future Oncol. 2017, 13, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Lundblad, V.; Blackburn, E.H. An alternative pathway for yeast telomere maintenance rescues est1− senescence. Cell 1993, 73, 347–360. [Google Scholar] [CrossRef]
- Bryan, T.M.; Englezou, A.; Gupta, J.; Bacchetti, S.; Reddel, R.R. Telomere Elongation in Immortal Human-Cells without Detectable Telomerase Activity. EMBO J. 1995, 14, 4240–4248. [Google Scholar] [PubMed]
- Dunham, M.A.; Neumann, A.A.; Fasching, C.L.; Reddel, R.R. Telomere maintenance by recombination in human cells. Nat. Genet. 2000, 26, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Verma, P.; Cho, N.W.; Winters, H.D.; Wondisford, A.R.; Greenberg, R.A. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 2016, 539, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Bryan, T.M.; Reddel, R.R. Telomere dynamics and telomerase activity in in vitro immortalised human cells. Eur. J. Cancer 1997, 33, 767–773. [Google Scholar] [CrossRef]
- Perrem, K.; Colgin, L.M.; Neumann, A.A.; Yeager, T.R.; Reddel, R.R. Coexistence of alternative lengthening of telomeres and telomerase in hTERT-transfected GM847 cells. Mol. Cell. Biol. 2001, 21, 3862–3875. [Google Scholar] [CrossRef] [PubMed]
- Yeager, T.R.; Neumann, A.A.; Englezou, A.; Huschtscha, L.I.; Noble, J.R.; Reddel, R.R. Telomerase-negative immortalized human cells contain a novel type of promyelocytic leukemia (PML) body. Cancer Res. 1999, 59, 4175–4179. [Google Scholar] [PubMed]
- Osterwald, S.; Deeg, K.I.; Chung, I.; Parisotto, D.; Worz, S.; Rohr, K.; Erfle, H.; Rippe, K. PML induces compaction, TRF2 depletion and DNA damage signaling at telomeres and promotes their alternative lengthening. J. Cell Sci. 2015, 128, 1887–1900. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Griffith, J.D. Telomeric DNA in ALT cells is characterized by free telomeric circles and heterogeneous T-loops. Mol. Cell. Biol. 2004, 24, 9948–9957. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Reddel, R.R. Assaying and investigating Alternative Lengthening of Telomeres activity in human cells and cancers. FEBS Lett. 2010, 584, 3800–3811. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.P.; Santos, G.; Vinagre, J.; Soares, P. The Role of ATRX in the Alternative Lengthening of Telomeres (ALT) Phenotype. Genes 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Shi, C.; Edil, B.H.; de Wilde, R.F.; Klimstra, D.S.; Maitra, A.; Schulick, R.D.; Tang, L.H.; Wolfgang, C.L.; Choti, M.A.; et al. DAXX/ATRX, MEN1, and mTOR pathway genes are frequently altered in pancreatic neuroendocrine tumors. Science 2011, 331, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Sampl, S.; Pramhas, S.; Stern, C.; Preusser, M.; Marosi, C.; Holzmann, K. Expression of telomeres in astrocytoma WHO grade 2 to 4: TERRA level correlates with telomere length, telomerase activity, and advanced clinical grade. Transl. Oncol. 2012, 5, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.; Chen, Y.J.; Taha, A.; Olivecrona, M.; Boet, R.; Wiles, A.; Warr, T.; Shaw, A.; Eiholzer, R.; Baguley, B.C.; Eccles, M.R.; Braithwaite, A.W.; Macfarlane, M.; Royds, J.A.; Slatter, T. Increased paired box transcription factor 8 has a survival function in glioma. BMC Cancer 2014, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.A.; Eiholzer, R.A.; Kirs, S.; Zhou, J.; Ward-Hartstonge, K.; Wiles, A.K.; Frampton, C.M.; Taha, A.; Royds, J.A.; Slatter, T.L. Telomere profiles and tumor-associated macrophages with different immune signatures affect prognosis in glioblastoma. Mod. Pathol. 2016, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Ulaner, G.A.; Hoffman, A.R.; Otero, J.; Huang, H.Y.; Zhao, Z.Q.; Mazumdar, M.; Gorlick, R.; Meyers, P.; Healey, J.H.; Ladanyi, M. Divergent patterns of telomere maintenance mechanisms among human sarcomas: Sharply contrasting prevalence of the alternative lengthening of telomeres mechanism in Ewing’s sarcomas and osteosarcomas. Genes Chromosomes Cancer 2004, 41, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Gagos, S.; Papaioannou, G.; Chiourea, M.; Merk-Loretti, S.; Jefford, C.E.; Mikou, P.; Irminger-Finger, I.; Liossi, A.; Blouin, J.L.; Dahoun, S. Unusually stable abnormal karyotype in a highly aggressive melanoma negative for telomerase activity. Mol. Cytogenet. 2008, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Viceconte, N.; Dheur, M.S.; Majerova, E.; Pierreux, C.E.; Baurain, J.F.; van Baren, N.; Decottignies, A. Highly aggressive metastatic melanoma cells unable to maintain telomere length. Cell Rep. 2017, 19, 2529–2543. [Google Scholar] [CrossRef] [PubMed]
- Royds, J.A.; Al Nadaf, S.; Wiles, A.K.; Chen, Y.J.; Ahn, A.; Shaw, A.; Bowie, S.; Lam, F.; Baguley, B.C.; Braithwaite, A.W.; et al. The CDKN2A G500 allele is more frequent in GBM patients with no defined telomere maintenance mechanism tumors and is associated with poorer survival. PLoS ONE 2011, 6, e26737. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Lingner, J. TERRA: Telomeric repeat-containing RNA. EMBO J. 2009, 28, 2503–2510. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat–containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, D.E.; Bensoussan, H.J.; Autexier, C. Telomerase Regulation from Beginning to the End. Genes 2016, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.J.; Lopez de Silanes, I.; Grana, O.; Blasco, M.A. Telomeric RNAs are essential to maintain telomeres. Nat. Commun. 2016, 7, 12534. [Google Scholar] [CrossRef] [PubMed]
- Azhibek, D.; Skvortsov, D.; Andreeva, A.; Zatsepin, T.; Arutyunyan, A.; Zvereva, M.; Dontsova, O. TERRA mimicking ssRNAs prevail over the DNA substrate for telomerase in vitro due to interactions with the alternative binding site. J. Mol. Recognit. 2016, 29, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Chartrand, P. Telomeric repeat-containing RNA TERRA: A noncoding RNA connecting telomere biology to genome integrity. Front. Genet. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Redon, S.; Reichenbach, P.; Lingner, J. The non-coding RNA TERRA is a natural ligand and direct inhibitor of human telomerase. Nucleic Acids Res. 2010, 38, 5797–5806. [Google Scholar] [CrossRef] [PubMed]
- De Silanes, I.L.; Grana, O.; De Bonis, M.L.; Dominguez, O.; Pisano, D.G.; Blasco, M.A. Identification of TERRA locus unveils a telomere protection role through association to nearly all chromosomes. Nat. Commun. 2014, 5, 4723. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.L.; Cox, K.E.; Jeitany, M.; Wakimoto, H.; Bryll, A.R.; Ganem, N.J.; Bersani, F.; Pineda, J.R.; Suva, M.L.; Benes, C.H.; et al. Alternative lengthening of telomeres renders cancer cells hypersensitive to ATR inhibitors. Science 2015, 347, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 regulates TERRA-telomeric DNA hybrids and telomere maintenance in ALT tumour cells. Nat. Commun. 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; Van Beneden, A.; Decottignies, A. Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1alpha. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Porro, A.; Feuerhahn, S.; Delafontaine, J.; Riethman, H.; Rougemont, J.; Lingner, J. Functional characterization of the TERRA transcriptome at damaged telomeres. Nat. Commun. 2014, 5, 5379. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Romero, C.A.P.; Chartrand, P. Telomeric noncoding RNA TERRA is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol. Cell 2013, 51, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.J.; Cropley, J.E.; Pickett, H.A.; Reddel, R.R.; Suter, C.M. Telomerase activity is associated with an increase in DNA methylation at the proximal subtelomere and a reduction in telomeric transcription. Nucleic Acids Res. 2009, 37, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, C.A.; Li, W.; Reisenweber, S.; Thongthip, S.; Bruno, J.; de Lange, T.; De, S.; Petrini, J.H.; Sung, P.A.; Jasin, M.; et al. Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway. PLoS Genet. 2012, 8, e1002772. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Telomerase-dependent (A,B) and -independent (C) telomere maintenance mechanisms (TMM) in cancer. Telomerase reactivation depends on several mechanisms that imply modifications that can have a direct impact on TERT gene regulation, which is localized at the short arm of chromosome 5. (A) TERTp hotspot mutations (−124 bp and −146 bp upstream the ATG transcriptional start site) create binding sites for ETS transcription factors (red boxes); (B) Germline genetic variations of the TERTp and of intronic and exonic regions seem to associate with cancer risk; their genomic coordinates based on build 37 (GRCh 37, hg19/Human); (C) TERT structural variants comprise amplification and rearrangement of the gene. Hypermethylation of the TERTp or other regions, micro RNA (miRNA) regulation and post-translational histone modifications are epigenetic modifications involved in telomerase reactivation; (D) Alternative lengthening of telomeres (ALT) is a telomerase-independent mechanism that relies on the homologous recombination machinery of DNA repair to maintain telomere length. Mutation of the genes ATRX or DAXX and loss of protein expression are known events related to ALT. miRNAs and TERRA molecules are some epigenetic regulators of ALT. TERT: telomerase reverse transcriptase; TERTp: TERT promoter.
Figure 1.
Telomerase-dependent (A,B) and -independent (C) telomere maintenance mechanisms (TMM) in cancer. Telomerase reactivation depends on several mechanisms that imply modifications that can have a direct impact on TERT gene regulation, which is localized at the short arm of chromosome 5. (A) TERTp hotspot mutations (−124 bp and −146 bp upstream the ATG transcriptional start site) create binding sites for ETS transcription factors (red boxes); (B) Germline genetic variations of the TERTp and of intronic and exonic regions seem to associate with cancer risk; their genomic coordinates based on build 37 (GRCh 37, hg19/Human); (C) TERT structural variants comprise amplification and rearrangement of the gene. Hypermethylation of the TERTp or other regions, micro RNA (miRNA) regulation and post-translational histone modifications are epigenetic modifications involved in telomerase reactivation; (D) Alternative lengthening of telomeres (ALT) is a telomerase-independent mechanism that relies on the homologous recombination machinery of DNA repair to maintain telomere length. Mutation of the genes ATRX or DAXX and loss of protein expression are known events related to ALT. miRNAs and TERRA molecules are some epigenetic regulators of ALT. TERT: telomerase reverse transcriptase; TERTp: TERT promoter.

Figure 2.
Frequency of telomere maintenance mechanisms in tumours by organ/anatomical site. The rates presented correspond to tumour types in which the following mechanisms were studied: TERTp mutations (TERTp mut) (orange), TERT amplification (TERT amp) (yellow), TERC amplification (TERC amp) (green), and ALT (brown). The studied population for each TMM is composed of the following cohorts (when not specified the data is depicted in Table 1, Table 2 and Table 4): Glioblastoma (GB), NOS (TERTp mut, n = 523; ALT, n = 953); oligodendroglioma (OD), NOS (TERTp mut, n = 469; ALT, n = 40); anaplastic astrocytoma (AA), NOS (TERTp mut, n = 89; ALT, n = 143); medulloblastoma (MB), NOS (TERTp mut, n = 166; TERT amp, n = 15; ALT, n = 192); hepatocellular carcinoma (HCC) (TERTp mut, n = 3091; TERT amp, n = 181; ALT, n = 121); papillary thyroid carcinoma (PTC), NOS (TERTp mut, n = 3256); pancreatic neuroendocrine tumour, NOS (pNET) (TERTp mut, n = 123; ALT, n = 849); basal cell carcinoma (BCC) (TERTp mut, n = 292); cutaneous melanoma, NOS (CM) (TERTp mut, n = 1975; TERT amp, n = 10; ALT, n = 106); acral lentiginous melanoma (ALM) (TERTp mut, n = 452; TERT amp, n = 60); myxoid liposarcoma (MLS) (TERTp mut, n = 76; ALT, n = 107); pleomorphic liposarcoma (PLS) (ALT, n = 26); leiomyosarcoma (LMS) (ALT, n = 161); gastrointestinal stromal tumour (GIST) (TERTp mut, n = 251; ALT, n = 50); muscle invasive bladder carcinoma (MIBC) (TERTp mut, n = 679; TERT amp, n = 3; ALT, n = 150); non-muscle invasive bladder carcinoma (NMIBC) (TERTp mut, n = 1682; TERT amp, n = 15); clear cell renal cell carcinoma (ccRCC) (TERTp mut, n = 443; ALT, n = 117); breast carcinoma (BRC) (TERT amp, n = 19; ALT, n = 377); oesophageal carcinoma (OEC) (TERTp mut, n = 403; TERC amp, n = 168; ALT, n = 136); non-small cell lung cancer (NSCLC) (TERTp mut, n = 961; TERT amp, n = 769; TERC amp, n = 176; ALT, n = 353). We assume for the percentage in which no defined TMM was reported that a non-defined telomere maintenance mechanisms (NDTMM) may be operating (grey).
Figure 2.
Frequency of telomere maintenance mechanisms in tumours by organ/anatomical site. The rates presented correspond to tumour types in which the following mechanisms were studied: TERTp mutations (TERTp mut) (orange), TERT amplification (TERT amp) (yellow), TERC amplification (TERC amp) (green), and ALT (brown). The studied population for each TMM is composed of the following cohorts (when not specified the data is depicted in Table 1, Table 2 and Table 4): Glioblastoma (GB), NOS (TERTp mut, n = 523; ALT, n = 953); oligodendroglioma (OD), NOS (TERTp mut, n = 469; ALT, n = 40); anaplastic astrocytoma (AA), NOS (TERTp mut, n = 89; ALT, n = 143); medulloblastoma (MB), NOS (TERTp mut, n = 166; TERT amp, n = 15; ALT, n = 192); hepatocellular carcinoma (HCC) (TERTp mut, n = 3091; TERT amp, n = 181; ALT, n = 121); papillary thyroid carcinoma (PTC), NOS (TERTp mut, n = 3256); pancreatic neuroendocrine tumour, NOS (pNET) (TERTp mut, n = 123; ALT, n = 849); basal cell carcinoma (BCC) (TERTp mut, n = 292); cutaneous melanoma, NOS (CM) (TERTp mut, n = 1975; TERT amp, n = 10; ALT, n = 106); acral lentiginous melanoma (ALM) (TERTp mut, n = 452; TERT amp, n = 60); myxoid liposarcoma (MLS) (TERTp mut, n = 76; ALT, n = 107); pleomorphic liposarcoma (PLS) (ALT, n = 26); leiomyosarcoma (LMS) (ALT, n = 161); gastrointestinal stromal tumour (GIST) (TERTp mut, n = 251; ALT, n = 50); muscle invasive bladder carcinoma (MIBC) (TERTp mut, n = 679; TERT amp, n = 3; ALT, n = 150); non-muscle invasive bladder carcinoma (NMIBC) (TERTp mut, n = 1682; TERT amp, n = 15); clear cell renal cell carcinoma (ccRCC) (TERTp mut, n = 443; ALT, n = 117); breast carcinoma (BRC) (TERT amp, n = 19; ALT, n = 377); oesophageal carcinoma (OEC) (TERTp mut, n = 403; TERC amp, n = 168; ALT, n = 136); non-small cell lung cancer (NSCLC) (TERTp mut, n = 961; TERT amp, n = 769; TERC amp, n = 176; ALT, n = 353). We assume for the percentage in which no defined TMM was reported that a non-defined telomere maintenance mechanisms (NDTMM) may be operating (grey).


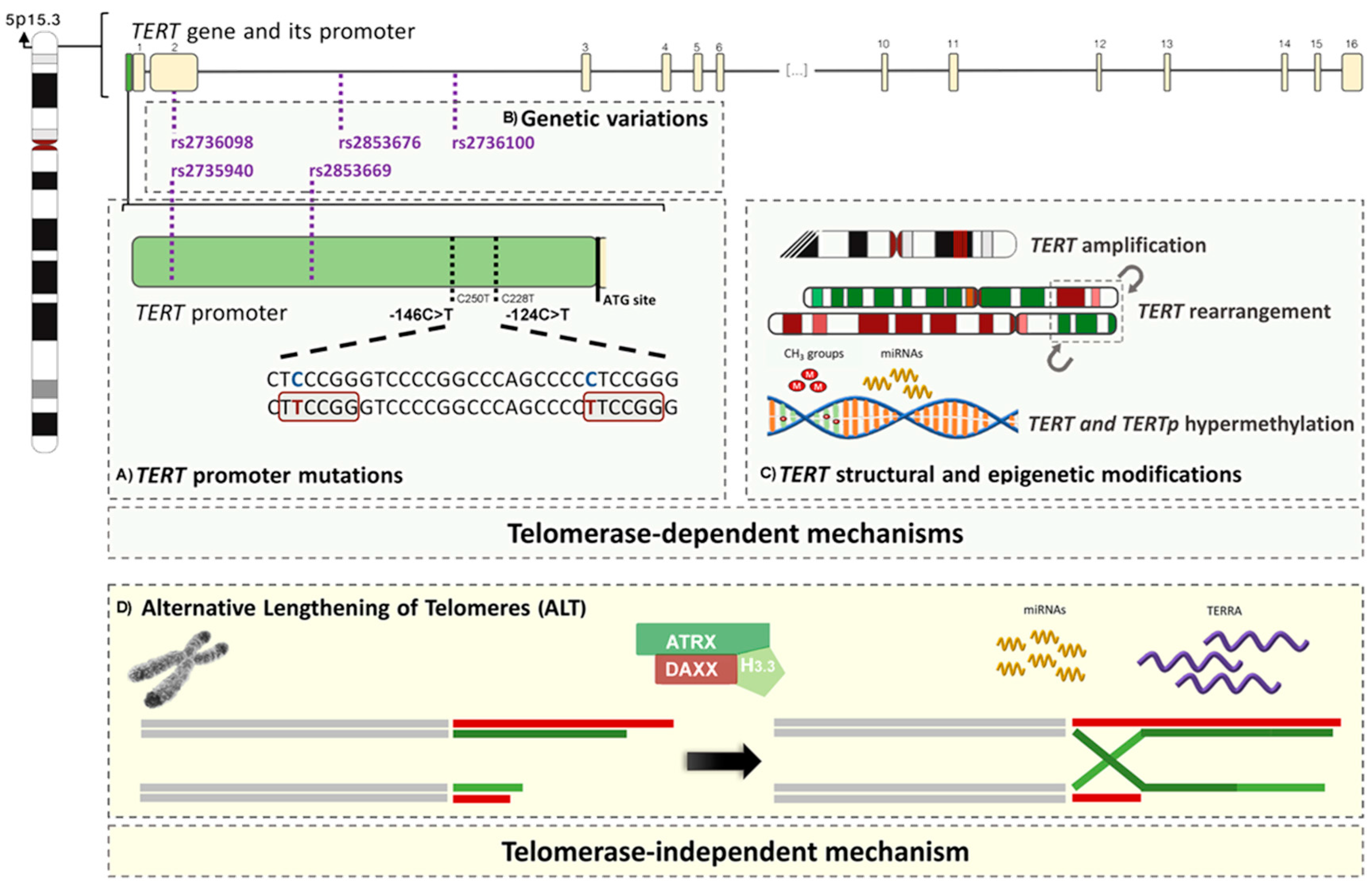{kind=link}
{kind=link}
Table 1.
Prevalence of TERTp mutations in human tumours. Only the tumour histotypes associated with a frequency of TERTp mutations ≥5% appear together with the respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of TERTp mutations (<5%) only the total of patients is shown (complete data available in Supplementary Table S1). The designations ‘NOS’ (not otherwise specified) and ‘SNS’ (site not specified) were applied to the tumours in which a specific histotype/site was not available.
Table 1.
Prevalence of TERTp mutations in human tumours. Only the tumour histotypes associated with a frequency of TERTp mutations ≥5% appear together with the respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of TERTp mutations (<5%) only the total of patients is shown (complete data available in Supplementary Table S1). The designations ‘NOS’ (not otherwise specified) and ‘SNS’ (site not specified) were applied to the tumours in which a specific histotype/site was not available.
| Tumour Type | Mutated Cases (Total) | Mutated Cases in % (Range) |
|---|---|---|
| Tumours of the breast | ||
| Fibroadenoma [58] | 4 (58) | 6.9 |
| Phyllodes tumour [58,59] | 41 (70) | 58.6 (45.8–65.2) |
| <5%: breast carcinoma, NOS (n = 88) and breast cancer, NOS (n = 122) [49,60]. | ||
| Tumours of the central nervous system | ||
| Astrocytoma (grades II-III), NOS [61,62,63] | 160 (699) | 22.9 (18.2–39.3) |
| Primary glioblastoma, NOS [50,64,65,66,67,68,69,70] | 1216 (1800) | 67.6 (46.3–85.9) |
| Secondary glioblastoma, NOS [50,64,65,67] | 25 (99) | 25.3 (5.0–35.7) |
| Paediatric glioblastoma, NOS [49,50] | 3 (51) | 5.9 (3.1–10.5) |
| Diffuse astrocytoma IDH-mutant [50,64,71,72,73] | 38 (242) | 15.7 (7.7–31.6) |
| Diffuse astrocytoma IDH-wildtype [50,64,71,72,73] | 23 (87) | 26.4 (7.1–58.3) |
| Diffuse astrocytoma, NOS [67,74,75,76,77] | 112 (574) | 19.5 (7.7–32.0) |
| Anaplastic astrocytoma, IDH-mutant [50,64,71,73,78,79] | 28 (248) | 11.3 (4.4–20.0) |
| Anaplastic astrocytoma, IDH-wildtype [50,64,71,73,78,79] | 104 (220) | 47.3 (20.0–80.0) |
| Anaplastic astrocytoma, NOS [67,74,75,76] | 29 (89) | 32.6 (10.0–40.0) |
| Glioblastoma, IDH-wildtype [61,62,63,64,67,68,71,74,77,78,79,80,81,82] | 2082 (2899) | 71.8 (27.6–91.7) |
| Giant cell [50] | 6 (17) | 35.3 |
| Gliosarcoma [50] | 21 (26) | 80.8 |
| Glioblastoma, IDH-mutant [61,62,63,64,67,68,71,74,77,78,79,80,81] | 114 (470) | 24.3 (0.0–76.9) |
| Glioblastoma, NOS a [66,75,76,83] | 328 (523) | 62.7 (46.3–84.0) |
| Oligodendroglioma, IDH-mutant and 1p/19q-codeleted [50,63] | 296 (311) | 95.2 |
| Oligodendroglioma, NOS [49,62,64,67,68,73,74,75,77] | 251 (469) | 53.5 (7.7–75.7) |
| Anaplastic oligodendroglioma, IDH-mutant and 1p/19q-codeleted [50] | 8 (38) | 21.1 |
| Anaplastic oligodendroglioma, NOS [49,50,64,67,73,74,76] | 171 (273) | 62.6 (0.0–88.5) |
| Oligodendroglioma (grades II–III), NOS [61,78] | 132 (152) | 86.8 (79.3–96.9) |
| Oligoastrocytoma, NOS b [49,50,61,62,64,67,73,74,76] | 222 (618) | 35.9 (3.8–57.9) |
| Anaplastic oligoastrocytoma, NOS [49,50,64,67,73,74,77] | 170 (415) | 41.0 (26.7–52.3) |
| Ependymoma c [49,50] | 6 (118) | 5.1 (0.0–6.9) |
| Choroid plexus carcinoma [50] | 1 (6) | 16.7 |
| Dysembryoplastic neuroepithelial tumour [49,50] | 1 (15) | 6.7 (0.0–33.3) |
| Desmoplastic infantile astrocytoma and ganglioglioma [50] | 1 (8) | 12.5 |
| Paraganglioma [50] | 1 (12) | 8.3 |
| Medulloblastoma, adult (>18 y) WNT-activated [50,84] | 5 (15) | 33.3 (30.8–50.0) |
| Medulloblastoma, adult (>18 y) SHH-MB [50,84,85,86] | 119 (134) | 88.8 (72.7–100.0) |
| Medulloblastoma, adult (>18 y), NOS [50] | 15 (23) | 65.2 |
| Medulloblastoma, paediatric SHH-MB d [50,84,86] | 49 (146) | 33.6 (20.0–31.9) |
| Medulloblastoma, paediatric, NOS [50,84,86] | 22 (121) | 18.2 (3.5–56.0) |
| Medulloblastoma, NOS [49,75,87] | 40 (166) | 24.1 (20.9–33.3) |
| Meningioma with malignant histology [88] | 5 (18) | 27.8 |
| <5%: pilocytic astrocytoma (n = 167) [50,75,76], pilomyxoid astrocytoma (n = 3) [50], subependymal giant cell astrocytoma (n = 11) [50,75], pleomorphic xanthoastrocytoma (n = 26) [50,75], mixopapillary ependymoma (n = 14) [50], subependymoma (n = 12) [50], anaplastic ependymoma (n = 48) [50], angiocentric glioma (n = 9) [50], choroid plexus papiloma (n = 13) [50], gangliocytoma (n = 2) [50], ganglioglioma (n = 40) [50], anaplastic ganglioglioma (n = 3) [50], papillary glioneuronal tumour (n = 1) [50], rosette-forming glioneuronal tumour (n = 6) [50], central neurocytoma (n = 28) [50], pineocytoma (n = 2) [50], pineal parenchymal tumours of intermediate differentiation (n = 9) [51], pineoblastoma (n = 5) [50], papillary tumours of the pineal region (n = 2) [50]), medulloblastoma, paediatric WNT-activated (n = 66) [50,84,86] and meningioma (n = 349) [49,75,89]. | ||
| Tumours of the conjunctiva and uvea | ||
| Primary acquired melanosis with atypia [90] | 2 (25) | 8.0 |
| Conjunctival melanoma [90,91,92] | 32 (84) | 38.1 (31.6–57.1) |
| Perilimbal squamous carcinoma [93] | 21 (48) | 43.8 |
| <5%: primary acquired melanosis without atypia (n = 13) [90], conjunctival naevi (n = 56) [90] and uveal melanomas (n = 202) [90,91,92,94]. | ||
| Tumours of the digestive system | ||
| Borderline hepatocellular adenoma/hepatocellular carcinoma [95] | 3 (18) | 16.7 |
| Hepatocellular carcinoma derived from hepatocellular adenoma [95,96] | 12 (25) | 48.0 (43.8–55.6) |
| Fibrolamellar hepatocellular carcinoma [49,97] | 1 (20) | 5.0 (0.0–12.5) |
| Hepatocellular carcinomas e [45,49,75,96,97,98,99,100,101,102,103,104,105,106,107,108,109] | 1263 (3093) | 40.8 (26.3–63.3) |
| Gallbladder carcinoma, NOS [49,75,110] | 15 (166) | 9.0 (0.0–50.0) |
| <5%: oesophageal squamous cell carcinoma (n = 313) [111], oesophageal adenocarcinoma, NOS (n = 90) [112], gastric moderately differentiated adenocarcinoma, NOS (n = 29) [113], moderately to poorly differentiated gastric adenocarcinoma, NOS (n = 39) [113], poorly differentiated gastric adenocarcinoma, NOS (n = 119) [113], gastric mucous adenocarcinoma (n = 13) [113], well-differentiated gastric cancer, NOS (n = 90) [110], poorly-differentiated gastric cancer, NOS (n = 178) [110], gastric carcinomas of both intestinal and diffuse types, NOS (n = 74) [75], colorectal adenocarcinoma, NOS (n = 22) [49], hepatoblastoma (n = 18) [49,114], hepatocellular adenoma, NOS (n = 238) [95,97], intrahepatic cholangiocarcinoma (n = 65) [97,99]), extrahepatic cholangiocarcinoma (n = 6) [75], extrahepatic cholangiosarcoma (n = 28) [49], pancreatic ductal adenocarcinoma (n = 24) [49], pancreatic acinar cell carcinoma (n = 25) [49] and pancreatic cancer, NOS (n = 46) [49]. | ||
| Tumours of the endocrine organs | ||
| Benign thyroid tumour, atypical follicular adenoma * [115] | 3 (18) | 16.7 |
| Papillary thyroid carcinoma, conventional variant [51,116,117,118,119,120,121,122,123,124,125] | 178 (1838) | 9.7 (2.9–15.2) |
| Papillary thyroid carcinoma, follicular variant [51,116,117,118,120,122,123,124,125] | 27 (481) | 5.6 (0.0–13.3) |
| Papillary thyroid carcinoma, tall cell variant [116,117,118,120,124,125,126] | 40 (214) | 18.7 (0.0–60.0) |
| Papillary thyroid carcinoma, NOS [51,118,119,120,123,125,127,128,129,130,131,132,133,134,135,136,137,138,139] | 361 (3288) | 11.0 (0.0–40.0) |
| Follicular thyroid carcinoma [51,115,116,119,120,122,125,128,129,132,135,138,139] | 102 (564) | 18.1 (13.9–50.0) |
| Hürthle cell carcinoma f [120,127,140] | 11 (146) | 7.5 (4.8–33.3) |
| Poorly differentiated thyroid carcinoma [51,116,120,122,125,127,133,135] | 88 (217) | 40.6 (21.4–51.7) |
| Anaplastic thyroid carcinoma [51,116,122,125,127,129,133,135,141,142] | 150 (326) | 46.0 (12.5–81.5) |
| Metastases of well-differentiated papillary and follicular thyroid carcinomas, NOS [118,125,138,143] | 90 (437) | 20.6 (15.0–50.0) |
| Adrenal cortical carcinoma [144] | 4 (34) | 11.8 |
| Extra-adrenal paraganglioma g [144] | 1 (9) | 11.1 |
| <5%: pituitary adenoma (n = 15) [145], follicular thyroid adenoma (n = 263) [51,115,122,128,139], benign thyroid tumour, NOS (n = 44) [146], papillary thyroid carcinoma, hobnail variant (n = 10) [147], papillary microcarcinoma (n = 435) [148,149], paediatric papillary carcinoma (n = 105) [150,151], minimally invasive Hürthle cell tumour (n = 52) [127,140], medullary thyroid carcinoma (n = 135) [49,51,116,119,129], adrenal cortical adenoma (n = 47) [144], phaechromocytoma (n = 122) [51,144], extra-adrenal benign paraganglioma * (n = 4) [144], pancreatic neuroendocrine tumour, NOS (n = 123) [49,57]. | ||
| Tumours of the female reproductive organs | ||
| Ovarian low-grade serous carcinoma [49,152] | 2 (41) | 5.0 (3.0–12.5) |
| Ovarian clear cell carcinoma [49,152,153] | 48 (301) | 15.9 (15.9–16.7) |
| Uterine endometrial carcinoma, NOS [49] | 2 (19) | 10.5 |
| Uterine clear cell carcinoma, NOS [153] | 3 (14) | 21.4 |
| Cervical squamous cell carcinoma [49,152,154,155] | 33 (225) | 14.7 (0.0–21.4) |
| Vulvar melanoma [156] | 2 (23) | 8.7 |
| <5%: high-grade ovarian serous carcinoma (n = 80) [152], ovarian endometrioid carcinoma (n = 43) [152], uterine endometrioid carcinoma (n = 24) [152], uterine leiomyosarcoma (n = 22) [152], uterine serous carcinoma (n = 12) [152], cervical adenocarcinoma, NOS (n = 25) [152] and vulvar melanoma (n = 23) [156]. | ||
| Tumours of the head and neck | ||
| Laryngeal carcinoma, NOS [157] | 64 (235) | 27.2 |
| Oral squamous cell carcinoma h [49,154,155,158] | 146 (295) | 49.5 (2.4–67.7) |
| Tongue squamous cell carcinoma [49] | 10 (28) | 35.7 |
| <5%: olfactory neuroblastoma (n = 11) [49] and salivary gland tumours (adenoid cystic carcinoma (n = 1) [159], adenocarcinoma, NOS (n = 1) [159], salivary duct carcinoma, NOS (n = 1) [159], epithelial-myoepithelial carcinoma (n = 1) [159], adenosquamous carcinoma (n = 1) [159], pleomorphic adenoma (n = 19) [159], basal cell adenoma (n = 1) [159], Warthin tumour (n = 8) [159]). | ||
| Tumours of haematopoietic and lymphoid tissues | ||
| Mantle cell lymphoma [160,161] | 8 (36) | 22.2 (0.0–33.3) |
| <5%: chronic myeloid leukaemia, NOS (n = 6) [49], acute myeloid leukaemia, NOS (n = 274) [49,162], B-cell acute lymphoblastic leukaemia, NOS (n = 12) [160], chronic lymphocytic leukaemia (n = 26) [49,160], marginal zone lymphoma (n = 16) [160], lymphoplasmacytic lymphoma (n = 7) [160], myeloma/plasmacytoma (n = 9) [160], follicular lymphoma (n = 13) [160], diffuse large B-cell lymphoma (DLBCL), NOS (n = 9) [160], plasmablastic lymphoma (n = 7) [160] and Burkitt lymphoma (n = 9) [160]. | ||
| Tumours of the lung, pleura and thymus | ||
| Epithelioid mesothelioma [163] | 10 (95) | 10.5 |
| Sarcomatoid, desmoplastic, and biphasic mesothelioma [163] | 6 (15) | 15.2 |
| <5%: lung adenocarcinoma, NOS (n = 533) [164,165,166,167], lung squamous cell carcinoma, NOS (n = 384) [164,165,166,167,168], lung adenosquamous carcinoma (n = 44) [154,164,165], mesothelioma, NOS (n = 4) [49], thymoma, NOS (n = 47) [75], type C thymic cancer * (n = 5) [75] and thymic neuroendocrine atypical carcinoma (n = 2) [75]. | ||
| Tumours of the peripheral nervous system | ||
| Absence of histotypes with a frequency of TERT promoter mutations equal or higher than 5%. | ||
| <5%: neuroblastoma, NOS (n = 153) [49,169]. | ||
| Tumours of the skin | ||
| Basal cell carcinoma [170,171,172,173] | 143 (292) | 49.0 (38.8–81.8) |
| Squamous cell carcinoma [49,154,170,172,173] | 57 (102) | 55.9 (20.0–74.1) |
| Bowen disease [172,173] | 2 (13) | 15.4 (9.1–50.0) |
| Superficial spreading melanoma [51,92,171,174,175] | 129 (380) | 33.9 (23.0–40.9) |
| Nodular melanoma [51,92,171,174,175] | 101 (184) | 54.9 (42.9–85.7) |
| Lentigo maligna [51,171,175] | 10 (41) | 24.4 (0.0–52.9) |
| Acral-lentiginous melanoma [51,92,171,174,175,176,177,178,179] | 40 (452) | 8.9 (0.0–27.0) |
| Desmoplastic melanoma i [180,181] | 43 (96) | 44.8 (22.9–85.0) |
| Cutaneous melanoma j [45,51,92,156,171,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190] | 826 (1975) | 41.8 (7.1–85.0) |
| Metastatic melanoma of unknown primary site k [178,179,182] | 36 (73) | 49.3 (25.0–66.7) |
| Metastatic cutaneous melanoma l [185,188] | 168 (221) | 76.0 (58.8–81.2) |
| Metastatic melanoma of other primary locations m [188] | 7 (13) | 53.8 |
| Mucosal melanoma, NOS [178,179,182] | 21 (211) | 10.0 (8.7–13.2) |
| Atypical fibroxanthoma [191] | 25 (27) | 92.6 |
| Pleomorphic dermal sarcoma ** [191] | 26 (34) | 76.5 |
| Merkel cell carcinoma [192,193] | 6 (63) | 9.5 (6.7–10.4) |
| <5%: cutaneous naevi (n = 9) [51]. | ||
| Tumours of the soft tissues and bone | ||
| Myxoid liposarcoma [49,53,194] | 51 (76) | 67.1 (23.1–79.2) |
| Solitary fibrous tumour [49,53,194,195] | 37 (175) | 21.1 (12.5–27.7) |
| Fibrosarcoma [49] | 1 (3) | 33.3 |
| Malignant peripheral nerve sheath tumours [49,53,194,196] | 11 (139) | 7.9 (0.0–9.6) |
| Malignant granular cell tumour [53] | 1 (2) | 50.0 |
| <5%: lipoma (n = 8) [49], atypical lipomatous tumour (n = 10) [49], dedifferentiated liposarcoma (n = 61) [194], pleomorphic liposarcoma (n = 15) [194], well differentiated liposarcoma (n = 18) [53], dermatofibrosarcoma protuberans (n = 16) [53,194], myxofibrosarcoma (n = 33) [49,53,194], low-grade fibromyxoid sarcoma (n = 18) [49,194], gastrointestinal leiomyoma (n = 5) [75], leiomyosarcoma (n = 49) [49,53,194] pleomorphic leiomyosarcoma (n = 5) [53], rhabdomyosarcoma, NOS (n = 5) [53], embryonal rhabdomyosarcoma (n = 8) [49], alveolar rhabdomyosarcoma (n = 7) [49], angiosarcoma of soft tissue (n = 9) [194], gastrointestinal stromal tumour (n = 251) [49,51,75,197], gastric schwannoma (n = 1) [75], synovial sarcoma (n = 48) [49,53,194], epithelioid sarcoma (n = 4) [194], alveolar soft part sarcoma (n = 9) [53,194], clear cell sarcoma of soft tissue (n = 6) [53,194], extraskeletal myxoid chondrosarcoma (n = 12) [49,53,194], chondrosarcoma, NOS (n = 21) [49,53], Ewing sarcoma (n = 6) [53] and undifferentiated high-grade pleomorphic sarcoma (n = 70) [49,50,53]. | ||
| Tumours of the urinary system and male genital organs | ||
| Clear cell renal cell carcinoma [198,199,200] | 39 (443) | 8.8 (0.0–12.2) |
| Chromophobe renal cell carcinoma [51,200] | 1 (12) | 8.3 (0–12.5) |
| Urothelial muscle invasive bladder carcinoma [201,202,203,204,205,206,207] | 463 (679) | 68.2 (48.8–85.2) |
| Urothelial non-muscle invasive bladder carcinoma [51,201,202,203,204,205,206,208,209] | 959 (1395) | 68.7 (44.3–85.4) |
| Urothelial bladder carcinoma [45,49,70,75,204,207,210,211] | 377 (591) | 63.8 (46.4–84.6) |
| Squamous cell carcinoma of the bladder [168,211,212] | 52 (79) | 65.8 (54.7–100.0) |
| Bladder adenocarcinoma n [210,213] | 4 (54) | 7.4 (0.0–28.6) |
| Papillary urothelial neoplasm of low malignant potential [204,214] | 26 (43) | 60.5 (28.6–75.9) |
| Urothelial carcinoma of upper urinary tract o [49,75,200,201,207,215,216] | 146 (293) | 49.8 (35.3–76.9) |
| Urothelial carcinoma of the ureter [200,214,215] | 26 (135) | 19.3 (11.1–50.0) |
| Micropapillary urothelial carcinoma p [217] | 33 (33) | 100.0 |
| Urothelial carcinoma q [154,218] | 71 (96) | 75.9 (70.0–100) |
| <5%: papillary renal cell carcinoma (n = 10) [51], nephrogenic adenoma (n = 10) [210], prostate squamous cell carcinoma (n = 26) [219], prostate carcinoma, NOS (n = 47) [49,201], ‘testicular carcinoma’ ***, NOS (n = 17) [201]. | ||
a includes cerebellar glioblastoma (n = 14) [50], gliomatosis (n = 10) [50], glioblastoma with oligodendroglial differentiation (n = 6) [50]; b includes oligoastrocytoma (grades II-III) (n = 58) [61]; c includes spinal ependymoma (n = 9); d these cases were not analysed for the TP53 status. There is a partial overlap of the study populations of Eckel-Passow et al. [71] and Pekmezci et al. [63]; e includes grade I-IV hepatocellular carcinoma and clear cell (n = 57) [107] and non-clear cell hepatocellular carcinoma, NOS (n = 259) [107]; f includes widely invasive (n = 126) [140] and minimaly invasive (n = 44) [140] Hürthle cell carcinomas; g these cases were classified according to the AFIP criteria [220]. It does not seem to exist an overlap of the study populations of Landa et al. [127] and Landa et al. [133]; it may exist an overlap of the study populations of Kim et al. [136] and Kim et al. [137] and the study populations of Melo et al. [125] and Melo et al. [138]; h includes buccal (n = 84) [158], gum (n = 34) [158], lip (n = 6) [158], tongue (n = 63) [158], floor of mouth (n = 22) [49,158], alveolar ridge (n = 1) [49], mandibule (n = 1) [49], hard palate (n = 2) [49], supraglottis (n = 4) [49], glottis (n = 1) [49], tonsil (n = 18) [49], larynx (n = 2) [49], oropharynx/hypopharynx (n = 1) [49] and hypopharynx (n = 1) [49]; i includes pure (n = 48) [49] and mixed (n = 28) [180] desmoplastic melanoma; j includes cutaneous melanomas, NOS and spitzoid melanocytic neoplasms (n = 56) [183], occult melanoma (n = 34), chronically sun-damaged (CSD) (n = 18) [179] and non-CSD melanomas (n = 12) [179]; k includes brain (n = 11) [182], skin (n = 9) [182], bone (n = 9) [182], liver (n = 13) [182], lung (n = 9) [182], visceral lymph nodes (n = 16) [182] metastases, and other SNS (site not-specified) metastases (n = 34) [178,179]; l includes superficial spreading melanoma (n = 100) [185], nodular melanoma (n = 56) [185], lentigo maligna (n = 1) [185] and other non-specified metastases (n = 156) [185,186]; m includes lymph node (n = 6) [188], brain (n = 3) [188], soft tissues (n = 2) [188], lung (n = 1) [188] and liver (n = 1) [188] metastases; n includes primary (n = 24) [212] and metastatic (n = 30) [210] bladder adenocarcinoma; o includes sarcomatoid urothelial carcinoma of upper urinary tract (n = 17) [216] and urothelial carcinoma of the renal pelvis (n = 205) [200,201,207,215]; p includes pure micropapillary urothelial carcinoma (n = 18) [217] and urothelial carcinoma with focal micropapillary features (n = 15) [217]; q includes SNS cases, low-grade (n = 28) [218] and high-grade urothelial carcinoma (n = 58) [218], and urothelial carcinoma with squamous differentiation (n = 10) [154]; * these designations are not in line with the current World Health Organization (WHO) classification; ** this designation is not in line with the current WHO classification. It may exist an overlap between the study populations of Heidenreich et al. [221], Nagore et al. [175] and Nagore et al. [186] and the study populations of Egberts et al. [182] and Egberts et al. [184]; it does not seem to exist an overlap of the study populations of Griewank et al. [91] and Griewank et al. [92]; there is a partial overlap of the study populations of Vinagre et al. [51] and Pópulo et al. [171]; *** this designation is not in line with the current WHO classification. It may exist an overlap between the study populations of Rachakonda et al. [204] and Hosen et al. [222].
Table 2.
Prevalence of TERT and TERC amplifications in human tumours. Only the tumour histotypes associated with a frequency of TERT and TERC amplifications ≥5% will appear in the following table together with its respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of TERT and TERC amplifications (<5%) only the total of patients will be shown (complete data available in Supplementary Table S1); the percentages of amplified cases here presented are the same reported by its respective authors, therefore the readers should note that the applied cut-off of copy number alterations may vary among references.
Table 2.
Prevalence of TERT and TERC amplifications in human tumours. Only the tumour histotypes associated with a frequency of TERT and TERC amplifications ≥5% will appear in the following table together with its respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of TERT and TERC amplifications (<5%) only the total of patients will be shown (complete data available in Supplementary Table S1); the percentages of amplified cases here presented are the same reported by its respective authors, therefore the readers should note that the applied cut-off of copy number alterations may vary among references.
| TERT Amplifications | ||
|---|---|---|
| Tumour Type | Amplified Cases (Total) | Amplified Cases in % (Range) |
| Tumours of the breast | ||
| Breast carcinoma, NOS a [52] | 5 (19) | 26.3 |
| <5%: phyllodes tumour (n = 73) [223]. | ||
| Tumours of the central nervous system | ||
| Pineoblastoma [224] | 1 (1) | 100 |
| Classic medulloblastoma [224] | 5 (13) | 38.5 |
| Nodular medulloblastoma [224] | 1 (10) | 10.0 |
| Anaplastic medulloblastoma [224] | 2 (5) | 40.0 |
| Medulloepithelyoma [224] | 2 (2) | 100.0 |
| Medullomyoepithelyoma [224] | 1 (2) | 50.0 |
| Ewing sarcoma/peripheral primitive neuroectodermal tumour [224] | 4 (8) | 50.0 |
| <5%: medullomyoblastoma (n = 1) [224]. | ||
| Tumours of the digestive system | ||
| Colorectal carcinoma, NOS [225] | 31 (64) | 48.4 |
| Hepatocellular carcinoma, NOS b [99,103,226] | 27 (181) | 14.9 (3.4–72.2) |
| Tumours of the endocrine system | ||
| Adrenal cortical carcinoma [45] | 11 (75) | 14.7 |
| Tumours of the female reproductive organs | ||
| Ovarian serous cystadenocarcinoma [45] | 6 (27) | 22.2 |
| Cervical intraepithelial neoplasia (CIN) 2 [227] | 6 (10) | 60 |
| Cervical intraepithelial neoplasia (CIN) 3 [227] | 7 (8) | 87.5 |
| Cervical carcinoma, NOS c [52,227] | 7 (14) | 50.0 |
| <5%: cervical intraepithelial neoplasia (CIN) 1 (n = 5) [227]. | ||
| Tumours of the head and neck | ||
| Pharyngeal/laryngeal squamous cell carcinoma, NOS [228] | 8 (81) | 9.9 |
| <5%: oral squamous cell carcinoma (n = 191) [228]. | ||
| Tumours of the lung | ||
| Lung carcinoma, NOS d [52] | 8 (21) | 38.1 |
| Lung adenocarcinoma, NOS [45,229,230] | 97 (529) | 18.3 (13.2–75.0) |
| Lung squamous cell carcinoma, NOS [45,229,230] | 59 (240) | 24.6 (13.8–63.9) |
| Lung large cell carcinoma [230] | 2 (5) | 40.0 |
| Mixed histology lung tumours [230] | 1 (5) | 20.0 |
| Tumours of the peripheral nervous system | ||
| Neuroblastoma, NOS [52] | 1 (8) | 12.5 |
| Tumours of the skin | ||
| Acral-lentiginous melanoma [231,232] | 14 (60) | 23.3 (20.1–29.4) |
| Melanoma e [233] | 4 (10) | 40.0 |
| Merkel cell carcinoma [192] | 11 (14) | 78.6 |
| <5%: desmoplastic melanoma (n = 62) [181]. | ||
| Tumours of the urinary system | ||
| Urothelial invasive bladder carcinoma [234] | 2 (3) | 66.7 |
| <5%: urothelial non-invasive bladder cancer (n = 15) [234]. | ||
| TERC amplifications | ||
| Tumours of the digestive system | ||
| Oesophageal carcinoma, NOS [45] | 36 (168) | 21.4 |
| Tumours of the female reproductive organs | ||
| Ovarian carcinoma f [45,235] Cervical intraepithelial neoplasia (CIN) 1 [227,236,237] Cervical intraepithelial neoplasia (CIN) 2 [227,236,237] Cervical intraepithelial neoplasia (CIN) 3 [227,236,237] Cervical carcinoma g [227,236,237,238,239,240] | 13 (35) 10 (41) 29 (44) 51 (58) 53 (90) | 37.1 (22.2–87.5) 24.4 (21.1–40.0) 69.0 (50.0–90.0) 87.9 (81.5–100.0) 58.9 (6.1–100.00) |
| Tumours of head and neck | ||
| Absence of histotypes with a frequency of TERC amplifications equal or higher than 5%. | ||
| <5%: squamous cell carcinoma of the head and neck, site not-specified (n = 31) [238]. | ||
| Tumours of the lung | ||
| Lung carcinoma, NOS [238] Lung squamous cell carcinoma, NOS [45] | 1 (9) 68 (167) | 11.1 40.7 |
a Includes poorly differentiated (n = 11) [52] and moderately differentiated (n = 8) [52] carcinomas; b includes moderately to highly differentiated hepatocellular carcinoma (n = 34) [226] and poorly differentiated hepatocellular carcinoma (n = 12) [226]; c includes invasive squamous carcinoma (n = 9) and in-situ adenocarcinoma (n = 1), of which one is well-differentiated, six are moderately differentiated and two are poorly differentiated [52]; d includes non-small cell lung cancer (n = 13) [52] and small cell lung cancer (n = 11) [52]; only 21 were analysed; e includes conventional (n = 7), fatal spitzoid (n = 1) and melanoma arising in giant congenital naevi (n = 2); f includes serous cystadenocarcinoma (n = 27) [45] and carcinoma, NOS (n = 8) [235]; g includes squamous cell carcinoma (n = 20) [236,237], adenocarcinoma, NOS (n = 12) [240], mucinous adenocarcinoma (n = 2) [236], minimal deviation adenocarcinoma (n = 1) [236], squamous cell carcinoma and adenocarcinoma (n = 4) [227], and carcinoma, NOS (n = 33) [238]. It does not seem to exist an overlap of the study populations of Andersson et al. (2006) [240] and Andersson et al. (2009) [237]. The designation ‘NOS’ (not otherwise specified) was applied to the tumours in which a specific histotype was not available.
Table 3.
Association of the most common TERT and TERTp polymorphisms with the risk of developing cancer.
Table 3.
Association of the most common TERT and TERTp polymorphisms with the risk of developing cancer.
| TERT Polymorphism | Cancers with Higher Risk of Development |
| rs2736100 | Acute lymphoblastic leukaemia (paediatric); myeloproliferative neoplasms; bladder, cervical, colorectal, gastric, lung and pancreas (exocrine) cancers, NOS; gliomas; oral squamous cell and papillary thyroid carcinomas [241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270] |
| rs2736098 | Bladder, breast, cervical, colorectal, lung, pancreas (exocrine) and prostate cancers, NOS; basal cell, hepatocellular and nasopharyngeal carcinomas, NOS [252,270,271,272,273,274,275,276,277] |
| rs2853676 | Breast, gastric, lung, prostate and ovary cancers, NOS; gliomas, NOS; melanomas, NOS [243,247,278,279,280,281,282,283,284,285,286,287] |
| TERT Promoter Polymorphism | Cancer Types with Higher Risk of Development |
| rs2853669 | Bladder, breast, gastric, lung and prostate cancers, NOS; gliomas, NOS; hepatocellular carcinomas, NOS; melanomas, NOS [68,69,76,80,162,188,222,265,275,288,289,290,291,292,293] |
| rs2735940 | Acute lymphoblastic leukaemia (paediatric); gastric and lung cancers, NOS [256,265,294] |
The designation ‘NOS’ (not otherwise specified) were applied to the tumours in which a specific histotype was not available.
Table 4.
Prevalence of alternative lengthening of telomeres (ALT) in human tumours. Only the tumour histotypes associated with ALT-positive phenotype in a frequency ≥5% will appear in the following table together with its respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of ALT-positive phenotype (<5%) only the total of patients will be shown (complete data available in Supplementary Table S1).
Table 4.
Prevalence of alternative lengthening of telomeres (ALT) in human tumours. Only the tumour histotypes associated with ALT-positive phenotype in a frequency ≥5% will appear in the following table together with its respective number, percentage, and range of mutated cases. Whenever the tumour histotypes were associated with a low rate of ALT-positive phenotype (<5%) only the total of patients will be shown (complete data available in Supplementary Table S1).
| Tumour Type | Positive Cases (Total) | Positive Cases in % (Range) |
|---|---|---|
| Tumours of the breast | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: invasive lobular carcinoma (n = 27) [55], tubular carcinoma (n = 9) [55], carcinoma with medullary features (n = 55) [55], mucinous carcinoma (n = 15) [55] and ductal carcinoma (n = 271) [55]. | ||
| Tumours of the central nervous system | ||
| Diffuse astrocytoma, NOS [55,295,296] | 22 (42) | 52.4 (27.3–63.0) |
| Anaplastic astrocytoma, NOS [55,295,296,297,298] | 63 (143) | 53.8 (26.4–100.0) |
| Astrocytoma, NOS [299] | 17 (50) | 34.0 |
| Anaplastic paediatric astrocytoma, NOS [296] | 26 (88) | 29.5 |
| Paediatric glioblastoma, NOS [55,300] | 17 (57) | 29.8 (12.0–43.8) |
| Glioblastoma, NOS a [55,295,297,298,301,302,303] | 147 (953) | 15.4 (11.4–50.0) |
| Oligodendroglioma, NOS [55] | 8 (40) | 20.0 |
| Anaplastic pleomorphic xanthoastrocytoma [295] | 2 (2) | 100.0 |
| Choroid plexus carcinoma b [300] | 7 (31) | 22.6 |
| Ewing sarcoma/peripheral primitive neuroectodermal tumour b [300] | 5 (43) | 11.6 |
| <5%: paediatric anaplastic astrocytoma (n = 24) [296], pilocytic astrocytoma (n = 45) [295,300], pleomorphic xanthoastrocytoma (n = 8) [300], ependymoma (n = 95) [300], choroid plexus papilloma (n = 24) [300], ganglioglioma (n = 8) [300], medulloblastoma, NOS [55,300], atypical teratoid/rhabdoid tumour (n = 38) [300], schwannoma (n = 44) [55], meningioma (n = 46) [55]. | ||
| Tumours of the digestive system | ||
| Hepatocellular carcinoma, NOS [55] | 8 (121) | 6.6 |
| Chromophobe hepatocellular carcinoma with abrupt anaplasia [304] | 11 (12) | 91.7 |
| Gastric adenocarcinoma, NOS [55,305] | 16 (197) | 8.1 (0.0–38.1) |
| <5%: oesophageal adenocarcinoma (n = 106) [55], oesophageal small cell neuroendocrine carcinoma (n = 1) [55], oesophageal squamous cell carcinoma (n = 29) [55], adenocarcinoma of the small intestine, NOS (n = 215) [55], colon adenoma, NOS (n = 136) [55], colon adenocarcinoma, NOS (n = 126) [55], hepatocellular adenoma, NOS (n = 17) [55], extrahepatic cholangiocarcinoma (n = 33) [55], adenocarcinoma of the gallbladder, NOS (n = 60) [55], pancreatic ductal adenocarcinoma (n = 448) [55], gastrointestinal carcinoid tumours, NOS/SNS (n = 47) [306] and carcinoid tumour, NOS/SNS (n = 32) [55]. | ||
| Tumours of the endocrine organs | ||
| Medullary thyroid carcinoma [307] | 11 (42) | 26.2 |
| Adrenal cortical carcinoma [308] | 3 (24) | 12.5 |
| Extra-adrenal paranganglioma [55] | 1 (8) | 12.5 |
| Pancreatic neuroendocrine tumour, NOS [306,309,310,311,312,313,314] | 255 (849) | 30.0 (14.9–61.0) |
| <5%: thyroid adenoma (n = 34) [55], papillary thyroid carcinoma, NOS (n = 47) [55,299], follicular thyroid carcinoma (n = 52) [55], parathyroid adenoma (n = 38) [55], adrenal adenoma (n = 14) [55], and phaeochromocytoma (n = 67) [55]. | ||
| Tumours of the female reproductive system | ||
| Diffuse malignant peritoneal mesothelioma c [315] | 10 (38) | 26.3 |
| Uterine carcinosarcoma [316] | 8 (16) | 50.0 |
| Uterine leiomyosarcoma [316] | 7 (8) | 87.5 |
| Uterine serous carcinoma [55] | 3 (41) | 7.3 |
| Uterine stromal sarcoma [316] | 4 (17) | 23.5 |
| <5%: ovarian serous carcinoma, NOS (n = 205) [55], ovarian mucinous carcinoma (n = 21) [55], ovarian endometrioid carcinoma (n = 72) [55], ovarian clear cell carcinoma (n = 56) [55], uterine endometrioid carcinoma (n = 64) [55], uterine clear cell carcinoma (n = 3) [55], uterine mixed mesodermal tumour (n = 4) [55], cervical squamous cell carcinoma (n = 152) [55] and cervical adenocarcinoma, NOS (n = 19) [55]. | ||
| Tumours of the head and neck | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: laryngeal squamous cell carcinoma (n = 29) [55], oral squamous cell carcinoma, NOS (n = 41) [55], salivary gland cylindroma (n = 28) [55], salivary gland carcinoma, NOS (n = 98) [55], pleomorphic adenoma (n = 45) [55] and Warthin tumour (n = 23) [55]. | ||
| Tumours of haematopoietic and lymphoid tissues | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: diffuse large B-cell lymphoma (DLBCL), NOS (n = 10) [55], nodular sclerosis classic Hodgkin lymphoma (n = 23) [55], mixed-cellularity classic Hodgkin lymphoma (n = 17) [55] and other subtypes of non-Hodgkin lymphoma, NOS (n = 54) [55]. | ||
| Tumours of the lung, pleura, thymus and heart | ||
| Absence of histotypes with a frequency of ALT equal or higher than 5%. | ||
| <5%: lung tumours (adenocarcinoma, NOS (n = 153) [55], papillary carcinoma (n = 15) [55], bronchoalveolar carcinoma, NOS (n = 40) [55], squamous cell carcinoma (n = 100) [55], small cell neuroendocrine carcinoma (n = 63) [55], carcinoid tumour, NOS (n = 3) [55], large cell carcinoma (n = 35) [55], other hystotypes, NOS (n = 15) [55] and thymoma, NOS (n = 37) [55]. | ||
| Tumours of the peripheral nervous system | ||
| Neuroblastoma, NOS [55,317] | 62 (124) | 50.0 (9.1–58.8) |
| <5%: ganglioneuroma (n = 3) [55]. | ||
| Tumours of the skin | ||
| Malignant melanoma, NOS [55] | 7 (106) | 6.6 |
| <5%: basal cell carcinoma (n = 57) [55], squamous cell carcinoma (n = 56) [55], benign naevus, NOS (n = 12) [55] and benign adnexal tumour (n = 15) [55]. | ||
| Tumours of the soft tissues | ||
| Liposarcoma, NOS [55,299,318] | 21 (84) | 25.0 (23.5–33.3) |
| Dedifferentiated liposarcoma [319,320,321] | 35 (106) | 33.0 (26.9–47.6) |
| Myxoid liposarcoma d [319,320,321] | 20 (107) | 18.5 (5.0–30.0) |
| Pleomorphic liposarcoma [320] | 21 (26) | 80.8 (72.7–100) |
| Fibrosarcoma and variants [55,299] | 5 (23) | 21.7 (14.3–100.0) |
| Myxofibrosarcoma [322] | 19 (25) | 76.0 |
| Leiomyosarcoma [55,299,323] | 91 (161) | 56.5 (33.3–61.5) |
| Rhabdomyosarcoma, NOS [55,299] | 2 (39) | 5.1 (0.0–5.7) |
| Embryonal rhabdomyosarcoma [322,324] | 7 (24) | 29.2 (12.5–37.5) |
| Epithelioid haemangioendothelioma [325] | 1 (7) | 14.3 |
| Angiosarcoma [55,325] | 18 (79) | 22.8 (11.1–24.3) |
| Neurofibroma [55] | 2 (2) | 100.0 |
| Malignant peripheral nerve sheath tumours [55,320,326] | 47 (75) | 62.7 (0.0–79.2) |
| Epithelioid sarcoma [55,299,322] | 2 (14) | 14.3 (0.0–33.3) |
| Alveolar soft part sarcoma [299,322] | 1 (8) | 12.5 (0.0–25.0) |
| Chondrosarcoma, NOS [55,299] | 1 (7) | 14.3 (0.0–33.3) |
| Osteosarcoma e [299,327,328] | 109 (173) | 63.0 (46.6–79.6) |
| Malignant fibrous histiocytic tumour * [299,329] | 39 (75) | 52.0 (32.6–80.0) |
| Undifferentiated pleomorphic sarcoma f [55,322] | 55 (86) | 64.0 (63.5–64.7) |
| Radiation-associated sarcoma, NOS [322] | 3 (15) | 20.0 |
| <5%: lipoma (n = 1) [55], solitary fibrous tumours (n = 7), dermatofibrosarcoma protuberans (n = 9) [322], benign fibrous hystiocytoma (n = 16) [55], giant cell tumour of the tendon sheath (n = 22) [55], alveolar rhabdomyosarcoma (n = 23) [322,324], capillary haemangioma (n = 32) [55], Kaposi’s sarcoma (n = 55) [55], gastrointestinal stromal tumours (n = 50) [55,322], Ewing sarcoma (n = 63) [55,322,327], synovial sarcoma (n = 24) [299,322], clear cell sarcoma (n = 5) [55], extraskeletal myxoid chondrosarcoma (n = 2) [322]. | ||
| Tumours of the urinary system and male genital system | ||
| Chromophobe renal cell carcinoma [55] | 4 (47) | 8.5 |
| Nephroblastoma [330] | 26 (32) | 81.3 |
| Sarcomatoid renal carcinoma [55] | 2 (27) | 7.4 |
| Bladder small cell neuroendocrine carcinoma [55] | 3 (13) | 23.1 |
| Non-seminomatous germ cell tumour, NOS [55] | 7 (46) | 15.2 |
| <5%: clear cell renal cell carcinoma (n = 117) [55], papillary renal cell carcinoma (n = 86) [55], renal oncocytoma (n = 18) [55], urothelial muscle invasive (n = 150) [55] and non-muscle invasive bladder carcinoma (n = 38) [55], non-invasive papillary urothelial carcinoma (n = 5) [55], sarcomatoid bladder carcinoma (n = 1) [55], squamous cell carcinoma of the bladder (n = 2) [55], prostate adenocarcinoma (n = 1152) [55], prostate small cell neuroendocrine carcinoma (n = 24) [55] and seminoma (n = 48) [55]. | ||
a Does not include paediatric glioblastoma; b includes paediatric patients; c includes epithelial tubulopapillary (n = 5) [315], epithelial solid (n = 30) [315] and biphasic (mixed epithelial and sarcomatoid) (n = 3) [315]; d includes round-cell myxoid liposarcoma (n = 45) [319,320]; e includes osteoblastic (n = 8) [55], chondroblastic (n = 3) [55] and periosteal (n = 1) [55] osteosarcoma, and osteosarcoma, NOS (n = 32) [55]; f includes cases classified as malignant fibrous histiocytic tumour [55]; * this designation is not in line with the current WHO classification. The designation ‘NOS’ (not otherwise specified) and ‘SNS’ (site not specified) were applied to the tumours in which a specific histotype was not available.
Table 5.
Molecular associations, prognostic and clinical implications of telomere maintenance mechanisms in human tumours.
Table 5.
Molecular associations, prognostic and clinical implications of telomere maintenance mechanisms in human tumours.
| Hepatocellular Carcinomas |
| TERTp mutations: marker of malignant progression [95,331,332]. |
| Well-differentiated thyroid carcinomas |
| TERTp mutations: association with larger tumours, older patient age, higher tumour stage, tumour recurrence, and distant metastases [51,116,119,126,333]; association with BRAFV600E mutations [117,126,141,146]. |
| Pancreatic neuroendocrine tumours |
| TERTp mutations: association with hereditary syndromes [57]. |
| ALT: association with protein loss and mutation of ATRX/DAXX [311,312,314]; indicator of more aggressive disease, metastases and worse patient survival [309,311,312,334]. |
| Diffuse astrocytic and oligodendroglial tumours |
| TERTp mutations: correlation with older patient age, higher tumour grade, tumour progression and worse overall survival [62,65,80,98]; combination of TERTp and IDH mutations for survival assessment: TERTp and IDH concomitant mutations confer the longest overall patient survival; TERTp mutations alone confer the lowest [61,71,73,74,78,79]. |
| TERT and TERTp polymorphisms: rs2736100 [335] and rs2853676 [336] affect the risk of tumour development; association of TERTp rs2853669 status with worse prognosis and worse survival in GB patients [68,76,80]; modifying effect of rs2853669 on TERTp mutations [65,68,76,80]. |
| ALT: association with ATRX loss [295,296,298,337]; identification of less aggressive GBs with longer patient survival [298,301,303]; mutual exclusivity with TERTp mutations [49,50]. |
| Epigenetic mechanisms: methylation of TERT promoter as a potential biomarker for malignancy in paediatric gliomas [338]. |
| Neuroblastomas |
| TERT rearrangements: indicator of poor prognosis, particularly in combination with MYCN amplification [54,339,340,341]. |
| ALT: association with chemoresistant tumours with unfavourable prognosis [342]. |
| Uterine cervical lesions |
| TERT and TERC amplifications: early identification of patients with low-grade lesions and higher progression risk in routinely liquid based cytology by Pap smears [227,236,237]. |
| Cutaneous melanomas |
| TERTp mutations: association with male gender, older patient age, tumour ulceration, higher Breslow’s thickness, and worse overall survival [92,175,179,182,184,221]; association with BRAFV600E mutations; combination used to identify tumours with aggressive behaviour [175,179,186]. |
| TERT and TERTp polymorphisms: association of TERTp polymorphism rs2853669 with TERTp mutations identify patients at risk of aggressive disease [175,188,293]. |
| Liposarcomas |
| ALT: association with ATRX loss, disease progression and poor clinical outcome [319,320]. |
| Urothelial bladder carcinomas |
| TERTp mutations: association with increased disease recurrence and reduced patient survival [209,215,222]; useful biomarker for patient screening as a non-invasive diagnostic and follow-up tool [206,209]; combination with FGFR3 mutations to identify tumours with poor prognosis [222]. |
| TERT amplification: potential biomarker to identify high-risk patients with disease progression [234]. |
| TERT and TERTp polymorphisms: association of TERTp rs2853669 with tumour recurrence and worse patient survival [204]. |
CIN: cervical intraepithelial neoplasia; GB: glioblastoma; ALT: alternative lengthening of telomeres; TERC: telomerase RNA component; TERT: telomerase reverse transcriptase.
Table 6.
Distribution of absent/low frequency telomere maintenance mechanisms (TMMs) in prevalent human tumours.
Table 6.
Distribution of absent/low frequency telomere maintenance mechanisms (TMMs) in prevalent human tumours.
| TERT Promoter Mutations, % (Total) | TERT and TERC Amplifications % (Total) | ALT, % (Total) | |
|---|---|---|---|
| Tumours of the breast | |||
| 0 (210) | 26 (19) | 0–3.7 (377) | |
| Tumours of the digestive system | |||
| Stomach | 0 (543) | N.A. | 0–8 (197) |
| Small intestine | N.A. | N.A. | 0 (215) |
| Colon and rectum | 0 (22) | 48 (64) | 0 (126) |
| Exocrine pancreas | 0 (95) | N.A. | 0 (448) |
| Tumours of haematopoietic and lymphoid tissues | |||
| 2 (424) | N.A. | 0 (104) | |
| Tumours of the lung | |||
| 1 (611) | 9 (976) | 0 (424) | |
| Tumours of the male genital organs | |||
| Prostate | 0 (99) | N.A. | 0 (1176) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gaspar, T.B.; Sá, A.; Lopes, J.M.; Sobrinho-Simões, M.; Soares, P.; Vinagre, J. Telomere Maintenance Mechanisms in Cancer. Genes 2018, 9, 241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9050241
AMA Style
Gaspar TB, Sá A, Lopes JM, Sobrinho-Simões M, Soares P, Vinagre J. Telomere Maintenance Mechanisms in Cancer. Genes. 2018; 9(5):241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9050241
Chicago/Turabian StyleGaspar, Tiago Bordeira, Ana Sá, José Manuel Lopes, Manuel Sobrinho-Simões, Paula Soares, and João Vinagre. 2018. "Telomere Maintenance Mechanisms in Cancer" Genes 9, no. 5: 241. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9050241
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.