Sex Chromosomes of the Iconic Moth Abraxas grossulariata (Lepidoptera, Geometridae) and Its Congener A. sylvata


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Chromosome and Polyploid Nuclei Preparations
2.3. Comparative Genomic Hybridization
2.4. Preparation of W-Chromosome Painting Probes
2.5. Preparation of Telomeric and rDNA Probes
2.6. Fluorescence In Situ Hybridization with W-Chromosome Painting Probes
2.7. Reprobing
2.8. FISH with Biotin-Labeled 18S rDNA Probe
2.9. FISH with Tyramide Signal Amplification (TSA-FISH)
2.10. Microscopy and Image Processing
2.11. Dating of Split between Abraxas grossulariata and A. sylvata
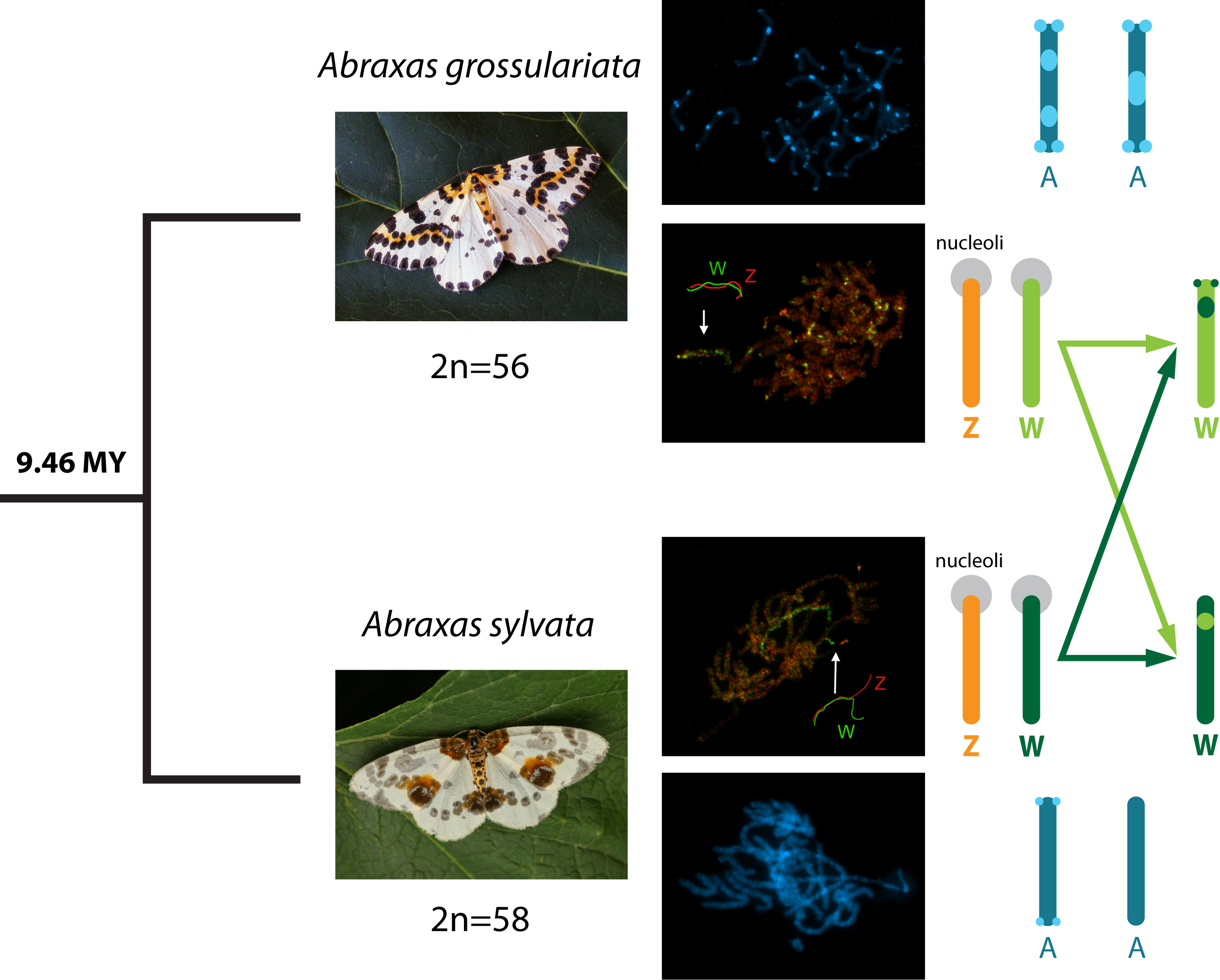
3. Results
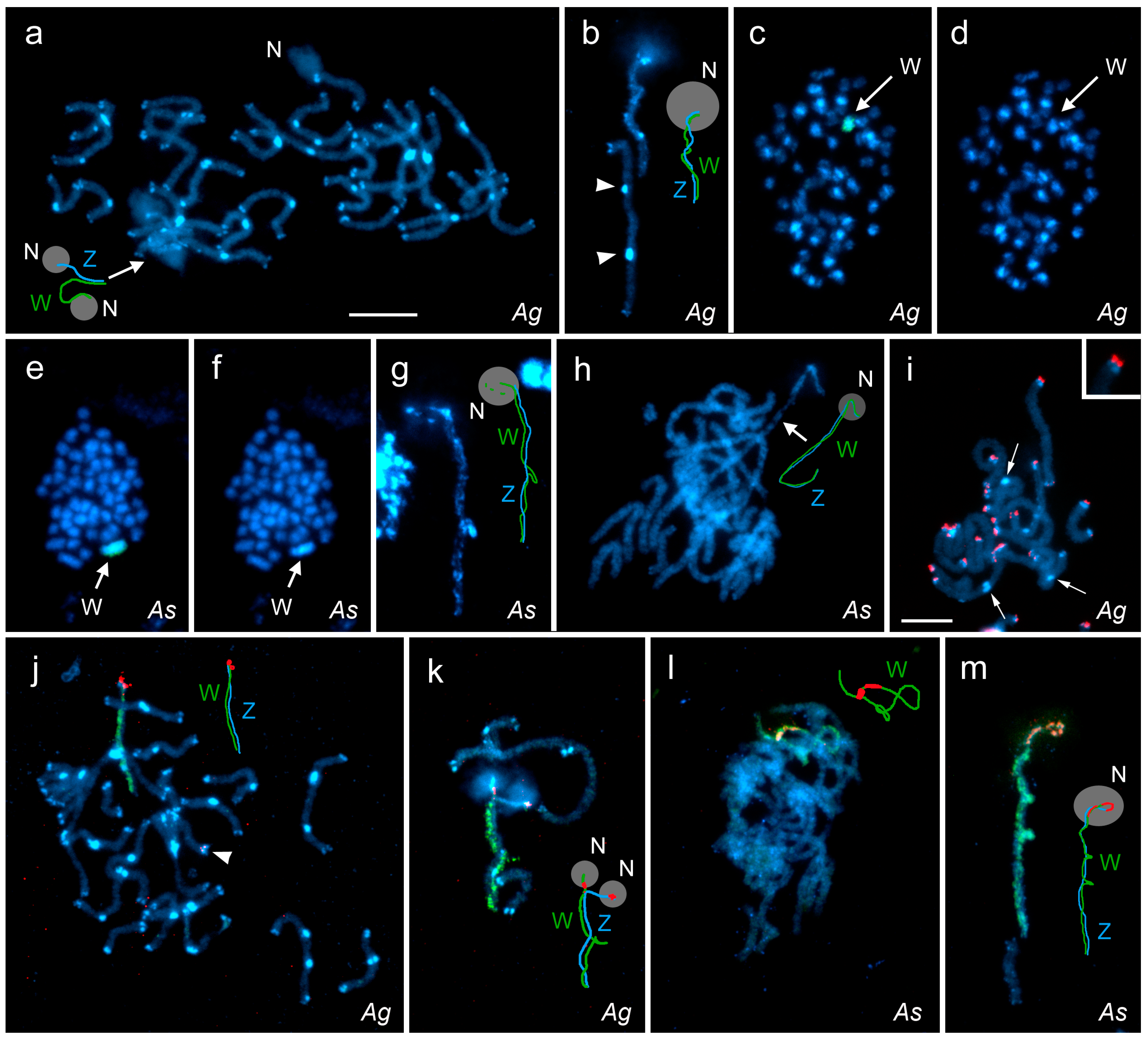
3.1. Basic Karyotype Characteristics
3.2. FISH with W-Chromosome Painting Probes and Localization of Major rDNA
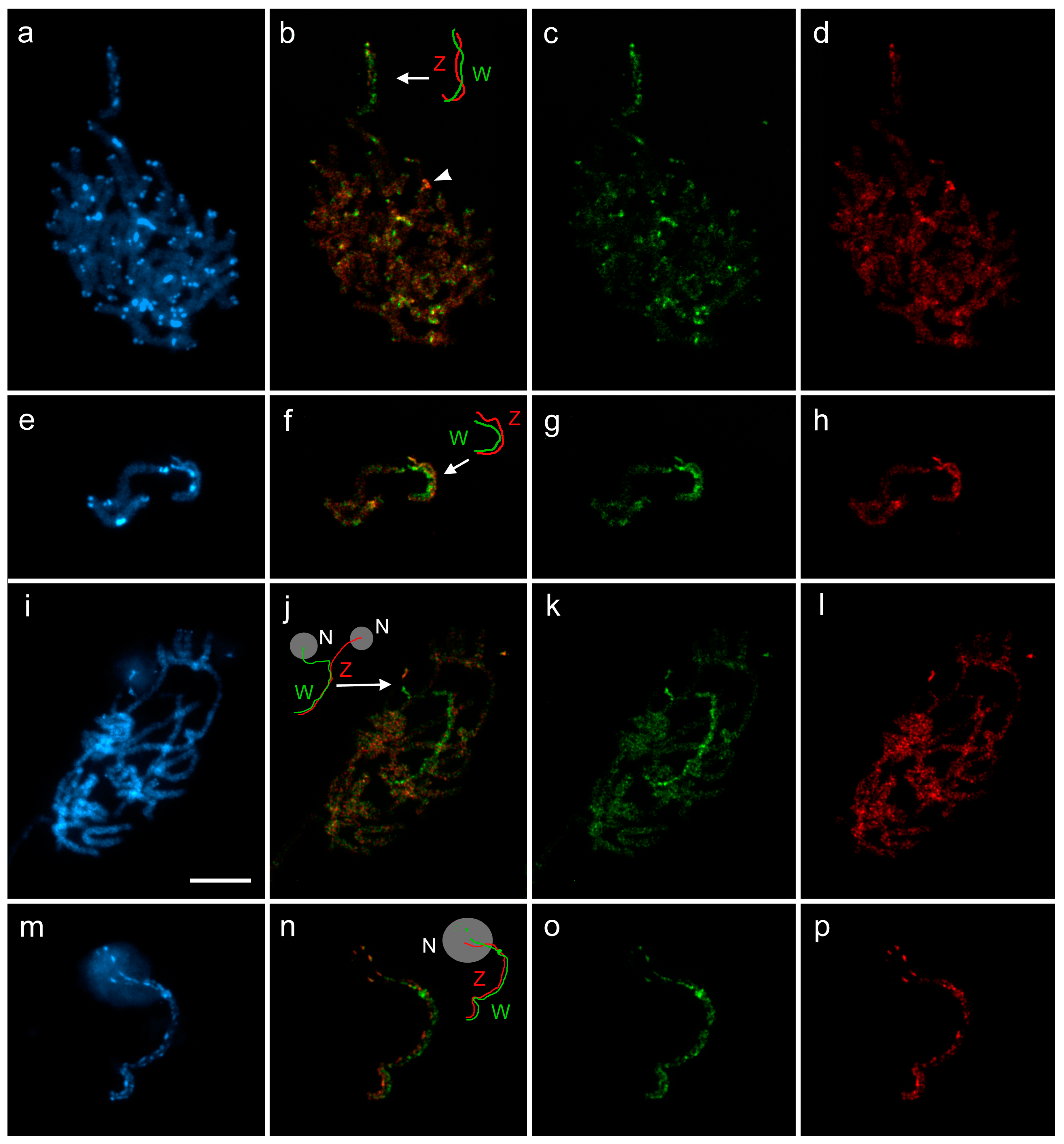
3.3. Cross-Species W-Chromosome Painting
3.4. Differentiation of W-Chromosomes by CGH
3.5. Dating of Split between Abraxas grossulariata and A. sylvata
4. Discussion
4.1. Sex Chromosome System in Abraxas Moths and Molecular Divergence of Their W-Chromosomes
4.2. Karyotype Diversification in Abraxas grossulariata and A. sylvata
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stevens, N.M. Studies in spermatogenesis with especial reference to the “accessory chromosome”. In Carnegie Institution of Washington, Publication No. 36; Henry E. Wilkens Printing Co.: Washington, DC, USA, 1905; 33p. [Google Scholar]
- Morgan, T.H.; Sturtevant, A.H.; Muller, H.J.; Bridges, C.B. The Mechanism of Mendelian Heredity; Henry Holt and Company: New York, NY, USA, 1915; 262p. [Google Scholar]
- Doncaster, L.; Raynor, G.H. On breeding experiments with Lepidoptera. Proc. Zool. Soc. Lond. 1906, 1, 125–133. [Google Scholar] [CrossRef]
- Doncaster, L. The chromosomes in the oogenesis and spermatogenesis of Pieris brassicae, and in the oogenesis of Abraxas grossulariata. J. Genet. 1912, 2, 189–200. [Google Scholar] [CrossRef]
- Seiler, J. Geschlechtschromosomen bei Lepidopteren. Zool. Anz. 1913, 41, 246–251. [Google Scholar]
- Traut, W.; Sahara, K.; Marec, F. Sex chromosomes and sex determination in Lepidoptera. Sex. Dev. 2007, 1, 332–346. [Google Scholar] [CrossRef] [PubMed]
- Marec, F.; Sahara, K.; Traut, W. Rise and fall of the W chromosome in Lepidoptera. In Molecular Biology and Genetics of the Lepidoptera; Goldsmith, M.R., Marec, F., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 49–63. [Google Scholar]
- Sahara, K.; Yoshido, A.; Traut, W. Sex chromosome evolution in moths and butterflies. Chromosome Res. 2012, 20, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Dopman, E.B.; Perez, L.; Bogdanowicz, S.M.; Harrison, R.G. Consequences of reproductive barriers for genealogical discordance in the European corn borer. Proc. Natl. Acad. Sci. USA 2005, 102, 14706–14711. [Google Scholar] [CrossRef] [PubMed]
- Putnam, A.S.; Scriber, J.M.; Andolfatto, P. Discordant divergence times among Z-chromosome regions between two ecologically distinct swallowtail butterfly species. Evolution 2007, 61, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Sýkorová, M.; Šíchová, J.; Kůta, V.; Dalíková, M.; Čapková Frydrychová, R.; Neven, L.G.; Sahara, K.; Marec, F. Neo-sex chromosomes and adaptive potential in tortricid pests. Proc. Natl. Acad. Sci. USA 2013, 110, 6931–6936. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Kost, S.; Heckel, D.G.; Yoshido, A.; Marec, F.; Groot, A.T. A Z-linked sterility locus causes sexual abstinence in hybrid females and facilitates speciation in Spodoptera frugiperda. Evolution 2016, 70, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Evolution of vertebrate sex chromosomes and dosage compensation. Nat. Rev. Genet. 2016, 17, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Marec, F.; Novák, K. Absence of sex chromatin corresponds with a sex-chromosome univalent in females of Trichoptera. Eur. J. Entomol. 1998, 95, 197–209. [Google Scholar]
- Bush, G.L. Female heterogamety in the family Tephritidae (Acalyptratae, Diptera). Am. Nat. 1966, 100, 119–126. [Google Scholar] [CrossRef]
- Kaiser, V.B.; Bachtrog, D. Evolution of sex chromosomes in insects. Annu. Rev. Genet. 2010, 44, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, T.; Juchault, P.; Mocquard, J.P. The evolution of sex determination in isopod crustaceans. BioEssays 1997, 19, 409–416. [Google Scholar] [CrossRef]
- Špakulová, M.; Casanova, J.C. Current knowledge on B chromosomes in natural populations of helminth parasites: A review. Cytogenet. Genome Res. 2004, 106, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Kongim, B.; Sutcharit, C.; Tongkerd, P.; Tan, S.H.A.; Quynh, N.X.; Naggs, F.; Panha, S. Karyotype variations in the genus Pollicaria (Caenogastropoda: Pupinidae). Zool. Stud. 2010, 49, 125–131. [Google Scholar]
- Parnes, S.; Khalaila, I.; Hulata, G.; Sagi, A. Sex determination in crayfish: Are intersex Cherax quadricarinatus (Decapoda, Parastacidae) genetically females? Genet. Res. 2003, 82, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.H.; Qiu, G.F. Female-only sex-linked amplified fragment length polymorphism markers support ZW/ZZ sex determination in the giant freshwater prawn Macrobrachium rosenbergii. Anim. Genet. 2013, 44, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kumari, R.; Sharma, V. Genetics of dioecy and causal sex chromosomes in plants. J. Genet. 2014, 93, 241–277. [Google Scholar] [CrossRef] [PubMed]
- Van Nieukerken, E.J.; Kaila, L.; Kitching, I.J.; Kristensen, N.P.; Lees, D.C.; Minet, J.; Mitter, C.; Mutanen, M.; Regier, J.C.; Simonsen, T.J.; et al. Order Lepidoptera Linnaeus, 1758. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.Q., Ed.; Magnolia Press: Auckland, New Zealand, 2011. [Google Scholar]
- Šíchová, J.; Voleníková, A.; Dincă, V.; Nguyen, P.; Vila, R.; Sahara, K.; Marec, F. Dynamic karyotype evolution and unique sex determination systems in Leptidea wood white butterflies. BMC Evol. Biol. 2015, 15, 89. [Google Scholar] [CrossRef] [PubMed]
- Šíchová, J.; Ohno, M.; Dincă, V.; Watanabe, M.; Sahara, K.; Marec, F. Fissions, fusions, and translocations shaped the karyotype and multiple sex chromosome constitution of the northeast-Asian wood white butterfly, Leptidea amurensis. Biol. J. Linn. Soc. 2016, 118, 457–471. [Google Scholar] [CrossRef]
- Šíchová, J.; Nguyen, P.; Dalíková, M.; Marec, F. Chromosomal evolution in tortricid moths: Conserved karyotypes with diverged features. PLoS ONE 2013, 8, e64520. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Carabajal Paladino, L. On the neo-sex chromosomes of Lepidoptera. In Evolutionary Biology; Pontarotti, P., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 171–185. [Google Scholar]
- Traut, W.; Sahara, K.; Otto, T.D.; Marec, F. Molecular differentiation of sex chromosomes probed by comparative genomic hybridization. Chromosoma 1999, 108, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Vítková, M.; Fuková, I.; Kubíčková, S.; Marec, F. Molecular divergence of the W chromosomes in pyralid moths (Lepidoptera). Chromosome Res. 2007, 15, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Van’t Hof, A.E.; Nguyen, P.; Dalíková, M.; Edmonds, N.; Marec, F.; Saccheri, I.J. Linkage map of the peppered moth, Biston betularia (Lepidoptera, Geometridae): A model of industrial melanism. Heredity 2013, 110, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Dalíková, M.; Zrzavá, M.; Hladová, I.; Nguyen, P.; Šonský, I.; Flegrová, M.; Kubíčková, S.; Voleníková, A.; Kawahara, A.Y.; Peters, R.S.; et al. New insights into the evolution of the W chromosome in Lepidoptera. J. Hered. 2017, 108, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Fraïsse, C.; Picard, M.A.L.; Vicoso, B. The deep conservation of the Lepidoptera Z chromosome suggests a non-canonical origin of the W. Nat. Commun. 2017, 8, 1486. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Nam, K.; Ellegren, H. Faster-Z evolution is predominantly due to genetic drift. Mol. Biol. Evol. 2010, 27, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Sackton, T.B.; Corbett-Detig, R.B.; Nagaraju, J.; Vaishna, L.; Arunkumar, K.P.; Hartl, D.L. Positive selection drives faster-Z evolution in silkmoths. Evolution 2014, 68, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Mita, K.; Yasukochi, Y.; Oshiki, T.; Shimada, T. Retrotransposable elements on the W chromosome of the silkworm, Bombyx mori. Cytogenet. Genome Res. 2005, 110, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Fuková, I.; Traut, W.; Vítková, M.; Nguyen, P.; Kubíčková, S.; Marec, F. Probing the W chromosome of the codling moth, Cydia pomonella, with sequences from microdissected sex chromatin. Chromosoma 2007, 116, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Vogel, H.; Glöckner, G.; Hartmann, E.; Heckel, D.G. High-throughput sequencing of a single chromosome: A moth W chromosome. Chromosome Res. 2013, 110, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Yoshido, A.; Šíchová, J.; Kubíčková, S.; Marec, F.; Sahara, K. Rapid turnover of the W chromosome in geographical populations of wild silkmoths, Samia cynthia ssp. Chromosome Res. 2013, 21, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Yoshido, A.; Marec, F.; Sahara, K. The fate of W chromosomes in hybrids between wild silkmoths, Samia cynthia ssp.: No role in sex determination and reproduction. Heredity 2016, 116, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Mediouni, J.; Fuková, I.; Frydrychová, R.; Dhouibi, M.H.; Marec, F. Karyotype, sex chromatin and sex chromosome differentiation in the carob moth, Ectomyelois ceratoniae (Lepidoptera: Pyralidae). Caryologia 2004, 57, 184–194. [Google Scholar] [CrossRef]
- Lockwood, A.P.M. “Ringer” solutions and some notes on the physiological basis of their ionic composition. Comp. Biochem. Physiol. 1961, 2, 241–289. [Google Scholar] [CrossRef]
- Kubickova, S.; Cernohorska, H.; Musilova, P.; Rubes, J. The use of laser microdissection for the preparation of chromosome-specific painting probes in farm animals. Chromosome Res. 2002, 10, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Drosopoulou, E.; Nakou, I.; Šíchová, J.; Kubíčková, S.; Marec, F.; Mavragani-Tsipidou, P. Sex chromosomes and associated rDNA form a heterochromatic network in the polytene nuclei of Bactrocera oleae (Diptera: Tephritidae). Genetica 2012, 140, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Sahara, K.; Marec, F.; Traut, W. TTAGG telomeric repeats in chromosomes of some insects and other arthropods. Chromosome Res. 1999, 7, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Albert, P.S.; Vega, J.M.; Bichler, J.A. Sensitive fluorescence in situ hybridization signal detection in maize using directly labeled probes produced by high concentration DNA polymerase nick translation. Biotech. Histochem. 2006, 81, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Fuková, I.; Nguyen, P.; Marec, F. Codling moth cytogenetics: Karyotype, chromosomal location of rDNA, and molecular differentiation of sex chromosomes. Genome 2005, 1092, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Shibata, F.; Sahara, K.; Naito, Y.; Yasukochi, Y. Reprobing multicolor FISH preparations in lepidopteran chromosome. Zool. Sci. 2009, 26, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Carabajal Paladino, L.Z.; Nguyen, P.; Šíchová, J.; Marec, F. Mapping of single-copy genes by TSA-FISH in the codling moth, Cydia pomonella. BMC Genet. 2014, 15 (Suppl. 2), S15. [Google Scholar] [CrossRef] [PubMed]
- Regier, J.C.; Mitter, C.; Zwick, A.; Bazinet, A.L.; Cummings, M.P.; Kawahara, A.Y.; Sohn, J.-C.; Zwickl, D.J.; Cho, S.; Davis, D.R.; et al. A large-scale, higher-level, molecular phylogenetic study of the insect order Lepidoptera (moths and butterflies). PLoS ONE 2013, 8, e58568. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, N.; Wheat, C.W.; Peña, C. Timing and patterns in the taxonomic diversification of Lepidoptera (butterflies and moths). PLoS ONE 2013, 8, e80875. [Google Scholar] [CrossRef] [PubMed]
- GenBank. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 12 March 2018).
- Õunap, E.; Viidalepp, J.; Truuverk, A. Phylogeny of the subfamily Larentiinae (Lepidoptera: Geometridae): Integrating molecular data and traditional classifications. Syst. Entomol. 2016, 41, 824–843. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Eickbush, T.H.; Eickbush, D.G. Finely orchestrated movements: Evolution of the ribosomal RNA genes. Genetics 2007, 175, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Skou, P.; Sihvonen, P.; Ennominae, I. The Geometrid Moths of Europe; Hausmann, A., Ed.; E. J. Brill: Leiden, The Netherlands, 2015; Volume 5, p. 657. [Google Scholar]
- Õunap, E.; Javoiš, J.; Viidalepp, J.; Tammaru, T. Phylogenetic relationships of selected European Ennominae (Lepidoptera: Geometridae). Eur. J. Entomol. 2011, 108, 267–273. [Google Scholar] [CrossRef]
- Sihvonen, P.; Mutanen, M.; Kaila, L.; Brehm, G.; Hausmann, A.; Staude, H.S. Comprehensive molecular sampling yields a robust phylogeny for geometrid moths (Lepidoptera: Geometridae). PLoS ONE 2011, 6, e20356. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Mosbacher, C. Geschlechtschromatin bei Lepidopteren. Chromosoma 1968, 25, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Sahara, K.; Marec, F.; Eickhoff, U.; Traut, W. Moth sex chromatin probed by comparative genomic hybridization (CGH). Genome 2003, 46, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Hallast, P.; Jobling, M.A. The Y chromosomes of the great apes. Hum. Genet. 2017, 136, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, H.P.; Ellegren, H. Old but not (so) degenerated—Slow evolution of largely homomorphic sex chromosomes in ratites. Mol. Biol. Evol. 2014, 31, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Sex-chromosome evolution: Recent progress and the influence of male and female heterogamety. Nat. Rev. Genet. 2011, 12, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.; Griffin, D.K.; Graves, J.A.M. Comparative painting reveals strong chromosome homology over 80 million years of bird evolution. Chromosome Res. 1999, 7, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Umehara, C.; Tsuda, Y.; Ishijima, J.; Ando, J.; Fujiwara, A.; Matsuda, Y.; Griffin, D.K. The molecular basis of chromosome orthologies and sex chromosomal differentiation in palaeognathous birds. Chromosome Res. 2007, 15, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Ellegren, H. Parallel divergence and degradation of the avian W sex chromosome. Trends Ecol. Evol. 2007, 22, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Poltronieri, J.; Bertollo, L.A.; Artoni, R.F.; Liehr, T.; Cioffi, M.B. Chromosomal mapping of repetitive DNAs in Triportheus trifurcatus (Characidae, Characiformes): Insights into the differentiation of the Z and W chromosomes. PLoS ONE 2014, 9, e90946. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Liehr, T.; Troy, W.P.; Cioffi, M.B. W chromosome dynamics in Triportheus species (Characiformes, Triportheidae): An ongoing process narrated by repetitive sequences. J. Hered. 2016, 107, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Mariguela, T.C.; Roxo, F.F.; Foresti, F.; Oliveira, C. Phylogeny and biogeography of Triportheidae (Teleostei: Characiformes) based on molecular data. Mol. Phylogenet. Evol. 2016, 96, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Sahara, K.; Yoshido, A.; Marec, F. Evolutionary dynamics of rDNA clusters on chromosomes of moths and butterflies (Lepidoptera). Genetica 2010, 138, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M. Satellite DNA: An evolving topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- López-Flores, I.; Garrido-Ramos, M.A. The repetitive DNA content of eukaryotic genomes. Genome Dyn. 2012, 7, 1–28. [Google Scholar] [PubMed]
- Talla, V.; Suh, A.; Kalsoom, F.; Dincă, V.; Vila, R.; Friberg, M.; Wiklund, C.; Backström, N. Rapid increase in genome size as a consequence of transposable element hyperactivity in wood-white (Leptidea) butterflies. Genome Biol. Evol. 2017, 9, 2491–2505. [Google Scholar] [CrossRef] [PubMed]
- Van’t Hof, A.E.; Marec, F.; Saccheri, I.J.; Brakefield, P.M.; Zwaan, B.J. Cytogenetic characterization and AFLP-based genetic linkage mapping for the butterfly Bicyclus anynana, covering all 28 karyotyped chromosomes. PLoS ONE 2008, 3, e3882. [Google Scholar] [CrossRef] [PubMed]
- Yoshido, A.; Marec, F.; Sahara, K. Resolution of sex chromosome constitution by genomic in situ hybridization and fluorescence in situ hybridization with (TTAGG)n telomeric probe in some species of Lepidoptera. Chromosoma 2005, 114, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zrzavá, M.; Hladová, I.; Dalíková, M.; Šíchová, J.; Õunap, E.; Kubíčková, S.; Marec, F. Sex Chromosomes of the Iconic Moth Abraxas grossulariata (Lepidoptera, Geometridae) and Its Congener A. sylvata. Genes 2018, 9, 279. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060279
Zrzavá M, Hladová I, Dalíková M, Šíchová J, Õunap E, Kubíčková S, Marec F. Sex Chromosomes of the Iconic Moth Abraxas grossulariata (Lepidoptera, Geometridae) and Its Congener A. sylvata. Genes. 2018; 9(6):279. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060279
Chicago/Turabian StyleZrzavá, Magda, Irena Hladová, Martina Dalíková, Jindra Šíchová, Erki Õunap, Svatava Kubíčková, and František Marec. 2018. "Sex Chromosomes of the Iconic Moth Abraxas grossulariata (Lepidoptera, Geometridae) and Its Congener A. sylvata" Genes 9, no. 6: 279. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060279