The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Construction of the Quorum Sensing Mutants
2.3. N-Acylhomoserine Lactone Profiling
2.4. Quantitative Reverse Transcription PCR
2.5. Motility, Autoagglutination and Haemagglutination Assays
2.6. Cell Attachment Assays
2.7. Virulence Plasmid Loss and spyA Expression
3. Results
3.1. Yersinia enterocolitica 8081 Possesses an Orphan luxR Ortholog
3.2. Comprehensive N-Acylhomoserine Lactone Profiling of the Yersinia enterocolitica 8081 Parent and Quorum Sensing Mutants
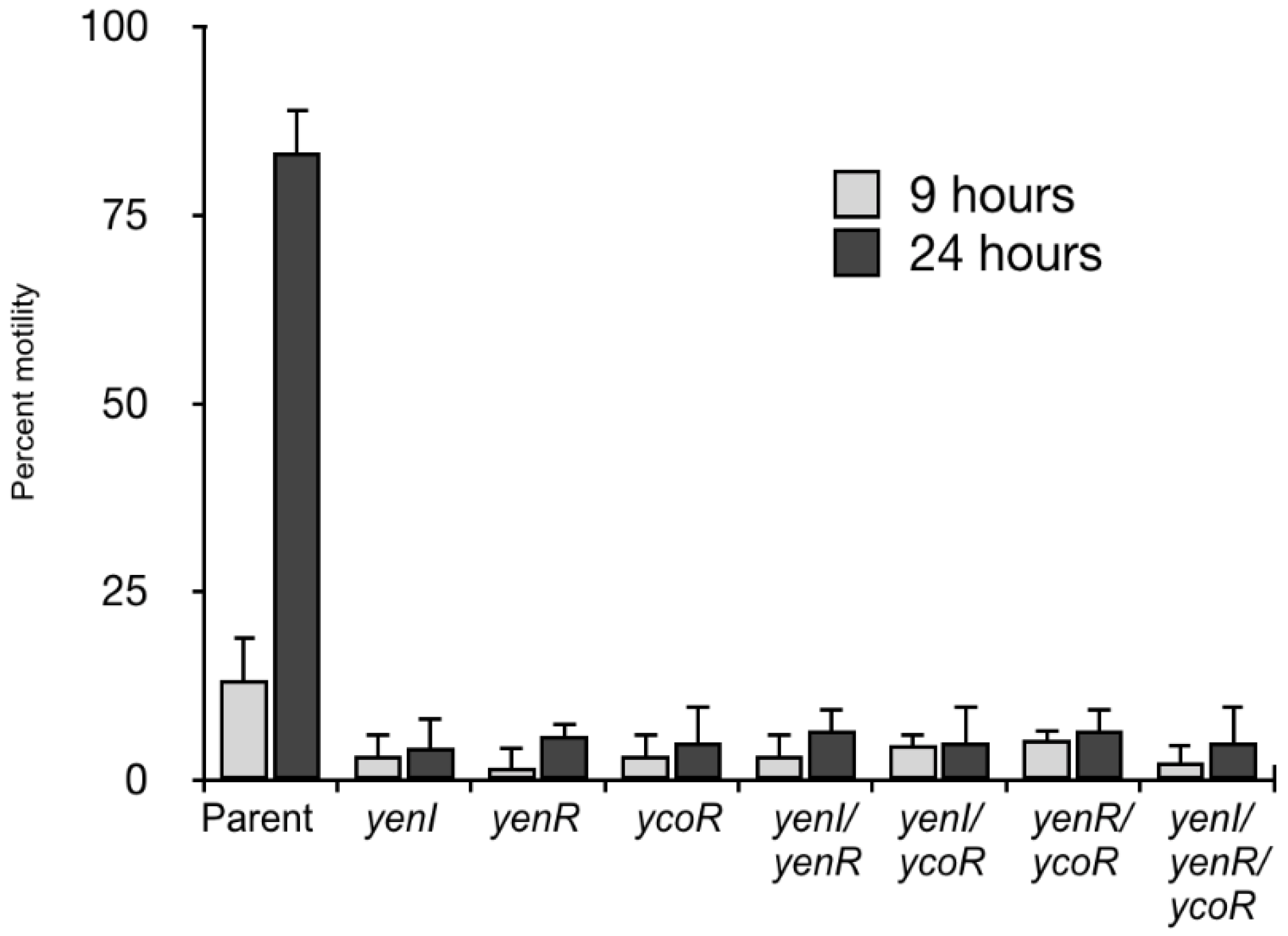
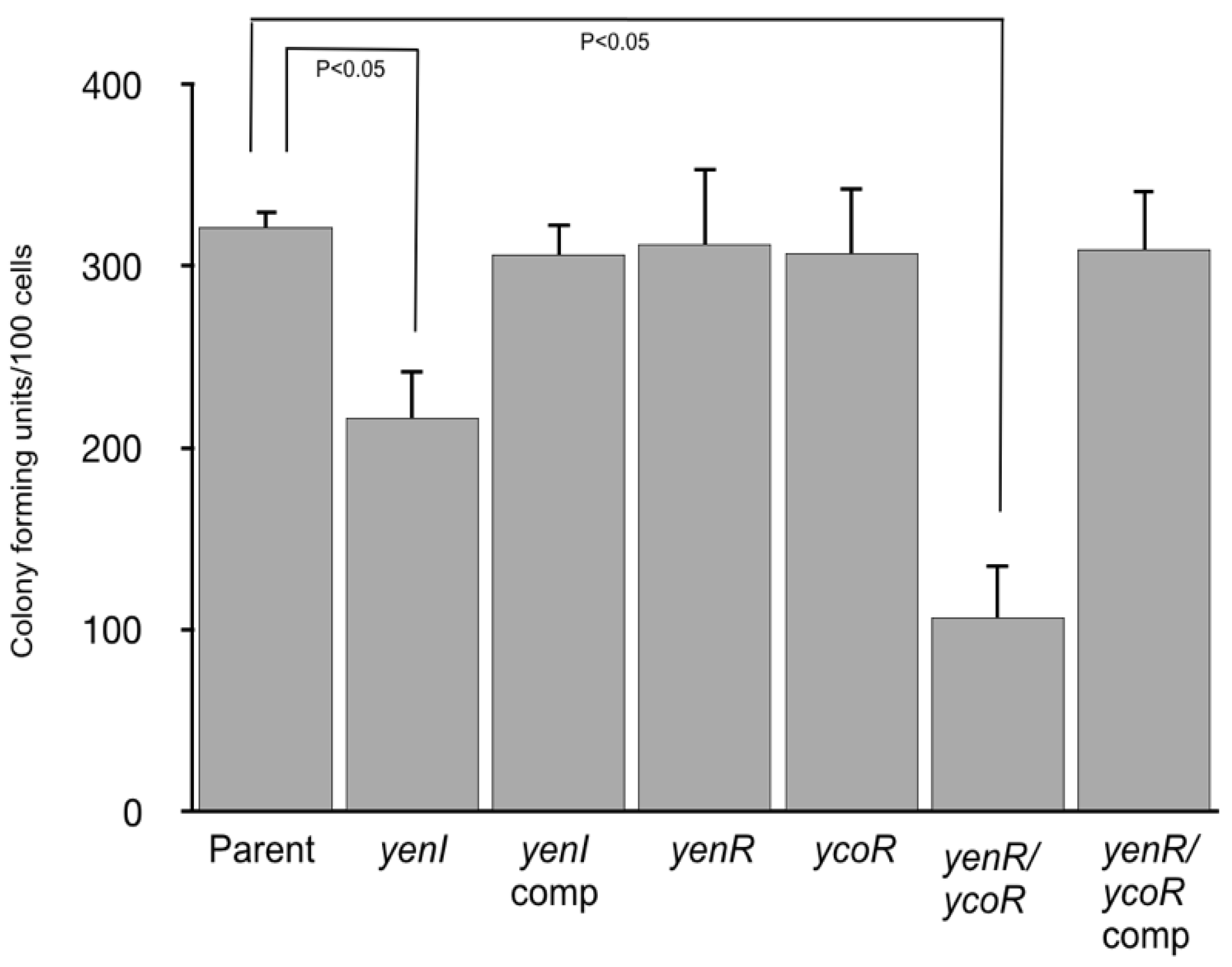
3.3. Phenotypic Analysis Reveals a Role for Quorum Sensing in Swimming Motility and Eukaryotic Cell Attachment but Not Type Three Secretion.
3.4. Quorum Sensing and Virulence Plasmid Maintenance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelis, G.R. The Yersinia Ysc-Yop “type III” weaponry. Nat. Rev. Mol. Cell Biol. 2002, 3, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Dewoody, R.S.; Merritt, P.M.; Marketon, M.M. Regulation of the Yersinia type III secretion system: Traffic control. Front. Cell. Infect. Microbiol. 2013, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Trosky, J.E.; Liverman, A.D.B.; Orth, K. Yersinia outer proteins: Yops. Cell. Microbiol. 2008, 10, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Ortori, C.A.; Atkinson, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Barrett, D.A. Comprehensive profiling of N-acylhomoserine lactones produced by Yersinia pseudotuberculosis using liquid chromatography coupled to hybrid quadrupole-linear ion trap mass spectrometry. Anal. Bioanal. Chem. 2007, 387, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, J.P.; Gould, T.A.; Schweizer, H.P.; Bearden, S.W.; Murphy, R.C.; Churchill, M.E.A. Quorum-sensing signal synthesis by the Yersinia pestis acyl-homoserine lactone synthase YspI. J. Bacteriol. 2006, 188, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Bobrov, A.G.; Bearden, S.W.; Fetherston, J.D.; Khweek, A.A.; Parrish, K.D.; Perry, R.D. Functional quorum sensing systems affect biofilm formation and protein expression in Yersinia pestis. Adv. Exp. Med. Biol. 2007, 603, 178–191. [Google Scholar] [PubMed]
- Atkinson, S.; Goldstone, R.J.; Joshua, G.W.P.; Chang, C.-Y.; Patrick, H.L.; Cámara, M.; Wren, B.W.; Williams, P. Biofilm development on Caenorhabditis elegans by Yersinia is facilitated by quorum sensing-dependent repression of type III secretion. PLoS Pathog. 2011, 7, e1001250. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.; Throup, J.P.; Stewart, G.S.; Williams, P. A hierarchical quorum-sensing system in Yersinia pseudotuberculosis is involved in the regulation of motility and clumping. Mol. Microbiol. 1999, 33, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.; Chang, C.-Y.; Sockett, R.E.; Cámara, M.; Williams, P. Quorum sensing in Yersinia enterocolitica controls swimming and swarming motility. J. Bacteriol. 2006, 188, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.S.; Winans, S.C. The quorum-hindered transcription factor YenR of Yersinia enterocolitica inhibits pheromone production and promotes motility via a small non-coding RNA. Mol. Microbiol. 2011, 80, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Kinder, S.A.; Badger, J.L.; Bryant, G.O.; Pepe, J.C.; Miller, V.L. Cloning of the YenI restriction endonuclease and methyltransferase from Yersinia enterocolitica serotype O8 and construction of a transformable R−M+ mutant. Gene 1993, 136, 271–275. [Google Scholar] [CrossRef]
- Thomson, N.R.; Howard, S.; Wren, B.W.; Holden, M.T.G.; Crossman, L.; Challis, G.L.; Churcher, C.; Mungall, K.; Brooks, K.; Chillingworth, T.; et al. The complete genome sequence and comparative genome analysis of the high pathogenicity Yersinia enterocolitica strain 8081. PLoS Genet. 2006, 2, 2039–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennox, E.S. Transduction of linked genetic characters of the host by bacteriophage P1. Virology 1955, 1, 190–206. [Google Scholar] [CrossRef]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y. N-Acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Maxson, M.E.; Darwin, A.J. Identification of inducers of the Yersinia enterocolitica phage shock protein system and comparison to the regulation of the RpoE and Cpx extracytoplasmic stress responses. J. Bacteriol. 2004, 186, 4199–4208. [Google Scholar] [CrossRef] [PubMed]
- Derbise, A.; Lesic, B.; Dacheux, D.; Ghigo, J.-M.; Carniel, E. A rapid and simple method for inactivating chromosomal genes in Yersinia. FEMS Immunol. Med. Microbiol. 2003, 38, 113–116. [Google Scholar] [CrossRef]
- Ng, Y.K. Novel Insights into the Quorum Sensing System of Yersinia enterocolitica 8081. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2010. [Google Scholar]
- Atkinson, S.; Sockett, R.E.; Cámara, M.; Williams, P. Quorum sensing and the lifestyle of Yersinia. Curr. Issues Mol. Biol. 2006, 8, 1–10. [Google Scholar] [PubMed]
- Laird, W.J.; Cavanaugh, D.C. Correlation of autoagglutination and virulence of Yersiniae. J. Clin. Microbiol. 1980, 11, 430–432. [Google Scholar] [PubMed]
- Di Biase, A.M.; Petrone, G.; Conte, M.P.; Seganti, L.; Ammendolia, M.G.; Tinari, A.; Iosi, F.; Marchetti, M.; Superti, F. Infection of human enterocyte-like cells with rotavirus enhances invasiveness of Yersinia enterocolitica and Yersinia pseudotuberculosis. J. Med. Microbiol. 2000, 49, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Pepe, J.C.; Badger, J.L.; Miller, V.L. Growth phase and low pH affect the thermal regulation of the Yersinia enterocolitica inv gene. Mol. Microbiol. 1994, 11, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Surgalla, M.J.; Beesley, E.D. Congo red-agar plating medium for detecting pigmentation in Pasteurella pestis. Appl. Microbiol. 1969, 18, 834–837. [Google Scholar] [PubMed]
- Riley, G.; Toma, S. Detection of pathogenic Yersinia enterocolitica by using Congo red-magnesium oxalate agar medium. J. Clin. Microbiol. 1989, 27, 213–214. [Google Scholar] [PubMed]
- Atkinson, S.; Chang, C.-Y.; Patrick, H.L.; Buckley, C.M.F.; Wang, Y.; Sockett, R.E.; Cámara, M.; Williams, P. Functional interplay between the Yersinia pseudotuberculosis YpsRI and YtbRI quorum sensing systems modulates swimming motility by controlling expression of flhDC and fliA. Mol. Microbiol. 2008, 69, 137–151. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, R.; Milton, D.L.; Wolf-Watz, H. Chemotactic motility is required for invasion of the host by the fish pathogen Vibrio anguillarum. Mol. Microbiol. 1996, 19, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Sitnikov, D.M.; Schineller, J.B.; Baldwin, T.O. Transcriptional regulation of bioluminescence genes from Vibrio fischeri. Mol. Microbiol. 1995, 17, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Subramoni, S.; Florez Salcedo, D.V.; Suarez-Moreno, Z.R. A bioinformatic survey of distribution, conservation, and probable functions of LuxR solo regulators in bacteria. Front. Cell. Infect. Microbiol. 2015, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.L.; Gaunt, M.W.; Hinds, J.; Witney, A.A.; Stabler, R.; Wren, B.W. Application of comparative phylogenomics to study the evolution of Yersinia enterocolitica and to identify genetic differences relating to pathogenicity. J. Bacteriol. 2006, 188, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Tahir, Y.E.; Skurnik, M. YadA, the multifaceted Yersinia adhesin. Int. J. Med. Microbiol. 2001, 291, 209–218. [Google Scholar] [PubMed]
- Kapperud, G.; Namork, E.; Skurnik, M.; Nesbakken, T. Plasmid-mediated surface fibrillae of Yersinia pseudotuberculosis and Yersinia enterocolitica: Relationship to the outer membrane protein YOP1 and possible importance for pathogenesis. Infect. Immun. 1987, 55, 2247–2254. [Google Scholar] [PubMed]
- Isberg, R.R.; Voorhis, D.L.; Falkow, S. Identification of invasin: A protein that allows enteric bacteria to penetrate cultured mammalian cells. Cell 1987, 50, 769–778. [Google Scholar] [CrossRef]
- Miller, V.L.; Falkow, S. Evidence for two genetic loci in Yersinia enterocolitica that can promote invasion of epithelial cells. Infect. Immun. 1988, 56, 1242–1248. [Google Scholar] [PubMed]
- Subramoni, S.; Venturi, V. LuxR-family ‘solos’: Bachelor sensors/regulators of signalling molecules. Microbiology 2009, 155, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; Wiehlmann, L.; Huber, B.; Jordan, D.; Lauber, J.; Salunkhe, P.; Limpert, A.S.; von Götz, F.; Steinmetz, I.; Eberl, L.; et al. Global regulation of quorum sensing and virulence by VqsR in Pseudomonas aeruginosa. Microbiology 2004, 150, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Chain, P.S.G.; Carniel, E.; Larimer, F.W.; Lamerdin, J.; Stoutland, P.O.; Regala, W.M.; Georgescu, A.M.; Vergez, L.M.; Land, M.L.; Motin, V.L.; et al. Insights into the evolution of Yersinia pestis through whole-genome comparison with Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 2004, 101, 13826–13831. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, J.K.; Wilkinson, A.; Hardman, A.; Rodelas, B.; Wisniewski-Dye, F.; Williams, P.; Downie, J.A. The regulatory locus cinRI in Rhizobium leguminosarum controls a network of quorum sensing loci. Mol. Microbiol. 2000, 37, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Horng, Y.-T.; Deng, S.-C.; Daykin, M.; Soo, P.-C.; Wei, J.-R.; Luh, K.-T.; Ho, S.-W.; Swift, S.; Lai, H.-C.; Williams, P. The LuxR family protein SpnR functions as a negative regulator of N-acylhomoserine lactone-dependent quorum sensing in Serratia marcescens. Mol. Microbiol. 2002, 45, 1655–1671. [Google Scholar] [CrossRef] [PubMed]
- Throup, J.P.; Cámara, M.; Bainton, N.J.; Briggs, G.S.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S. Characterisation of the yenI/yenR locus from Yersinia enterocolitica mediating the synthesis of two N-acyl homoserine lactone signal molecules. Mol. Microbiol. 1995, 17, 345–356. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Cámara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143 (Pt 12), 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Young, G.M. Flagella: Organelles for motility and protein secretion. In Yersinia Molecular and Cellular Biology; Carniel, E., Hinnebusch, B.J., Eds.; Horizon Bioscience: Wymondham, Norfolk, UK, 2004; pp. 243–256. [Google Scholar]
- Gaillard, J.L.; Berche, P.; Mounier, J.; Richard, S.; Sansonetti, P. In vitro model of penetration and intracellular growth of Listeria monocytogenes in the human enterocyte-like cell line Caco-2. Infect. Immun. 1987, 55, 2822–2829. [Google Scholar] [PubMed]
- Curfs, J.H.; Meis, J.F.; Fransen, J.A.; van der Lee, H.A.; Hoogkamp-Korstanje, J.A. Interactions of Yersinia enterocolitica with polarized human intestinal Caco-2 cells. Med. Microbiol. Immunol. 1995, 184, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Chugani, S.; Greenberg, E.P. LuxR homolog-independent gene regulation by acyl-homoserine lactones in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2010, 107, 10673–10678. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bhaduri, S.; Magee, W.E. Maximizing plasmid stability and production of released proteins in Yersinia enterocolitica. Appl. Environ. Microbiol. 1998, 64, 1812–1815. [Google Scholar] [PubMed]
- Wang, H.; Avican, K.; Fahlgren, A.; Erttmann, S.F.; Nuss, A.M.; Dersch, P.; Fallman, M.; Edgren, T.; Wolf-Watz, H. Increased plasmid copy number is essential for Yersinia T3SS function and virulence. Science 2016, 353, 492–495. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, Y.-K.; Grasso, M.; Wright, V.; Garcia, V.; Williams, P.; Atkinson, S. The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance. Genes 2018, 9, 307. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060307
Ng Y-K, Grasso M, Wright V, Garcia V, Williams P, Atkinson S. The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance. Genes. 2018; 9(6):307. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060307
Chicago/Turabian StyleNg, Yen-Kuan, Marco Grasso, Victoria Wright, Vanina Garcia, Paul Williams, and Steve Atkinson. 2018. "The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance" Genes 9, no. 6: 307. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9060307