Effects of Asian Dust and Phosphorus Input on Abundance and Trophic Structure of Protists in the Southern Yellow Sea

 ,
,
Abstract
:1. Introduction
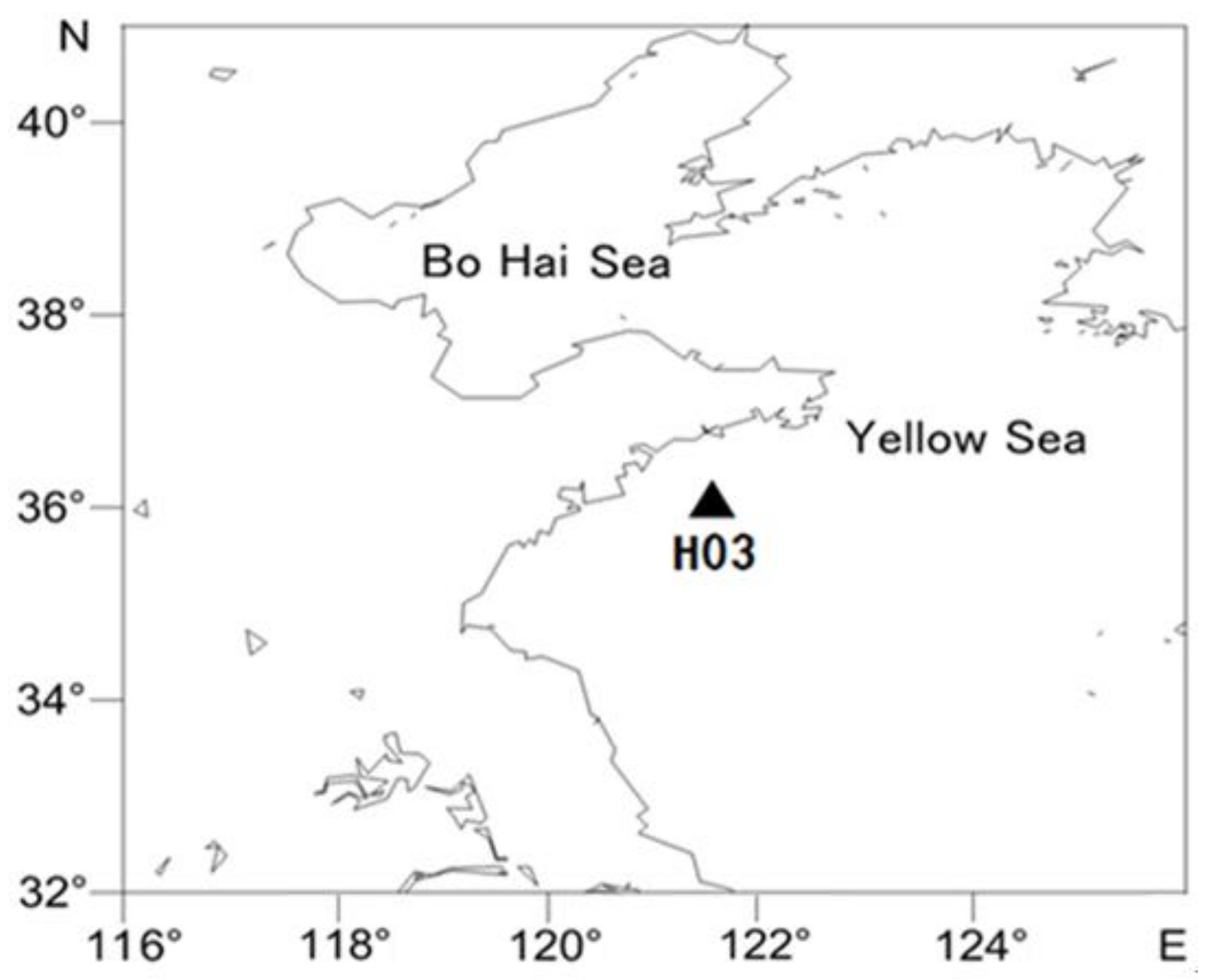
2. Materials and Methods
2.1. Sand-Dust Preparation
2.2. Onboard Culturing Experiment
2.3. Sample Collection and Measurement
2.3.1. Nanoplanktonic Protists Samples
2.3.2. Dissolved Organic Carbon (DOC)
2.3.3. Inorganic Nutrients
2.4. Data Processing and Analysis
3. Results and Analysis
3.1. Effects of Dust and Phosphorus Addition on the Abundance of Nanoplanktonic Protists
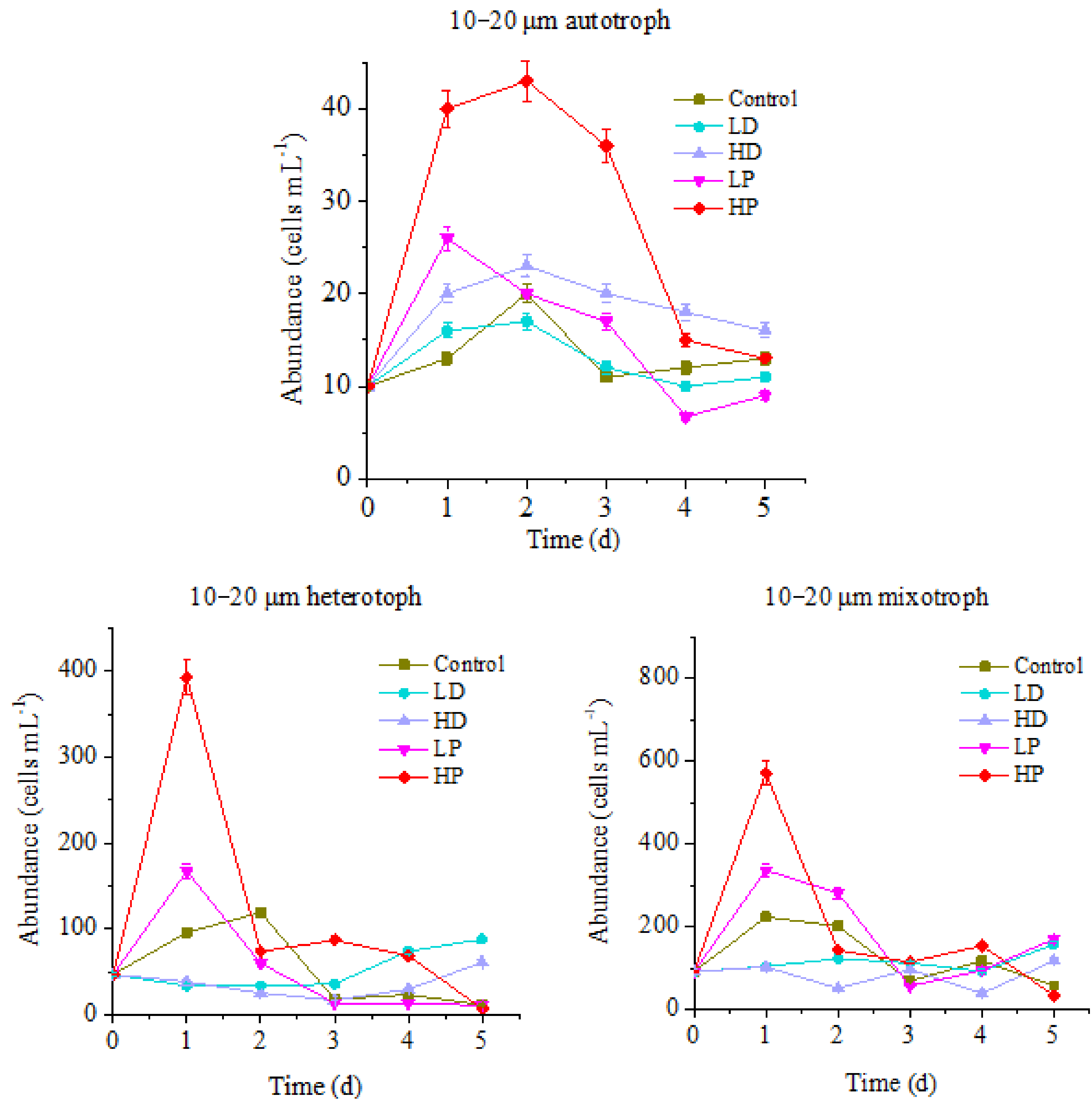
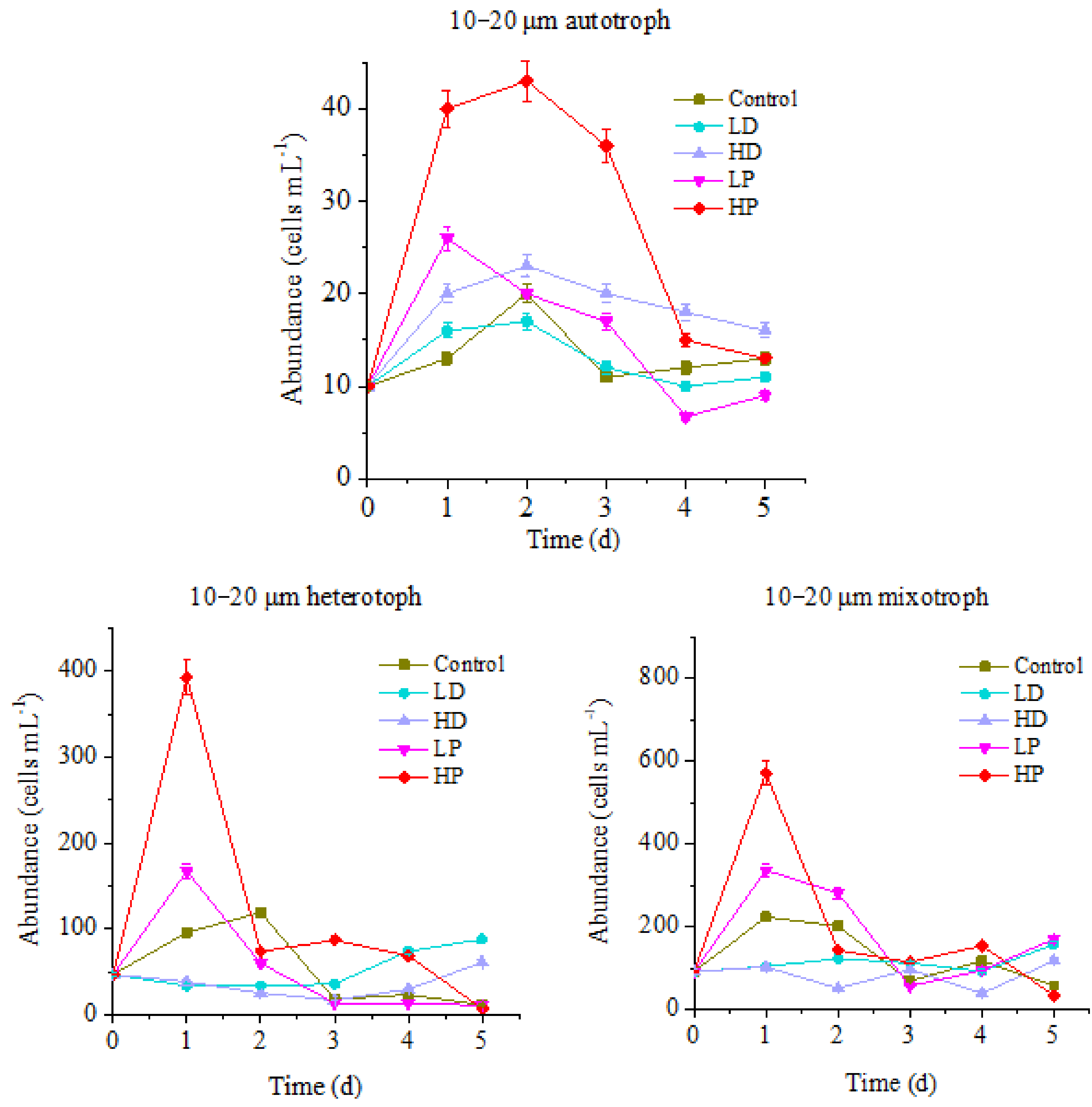
3.1.1. Changes of 10–20 µm Protists
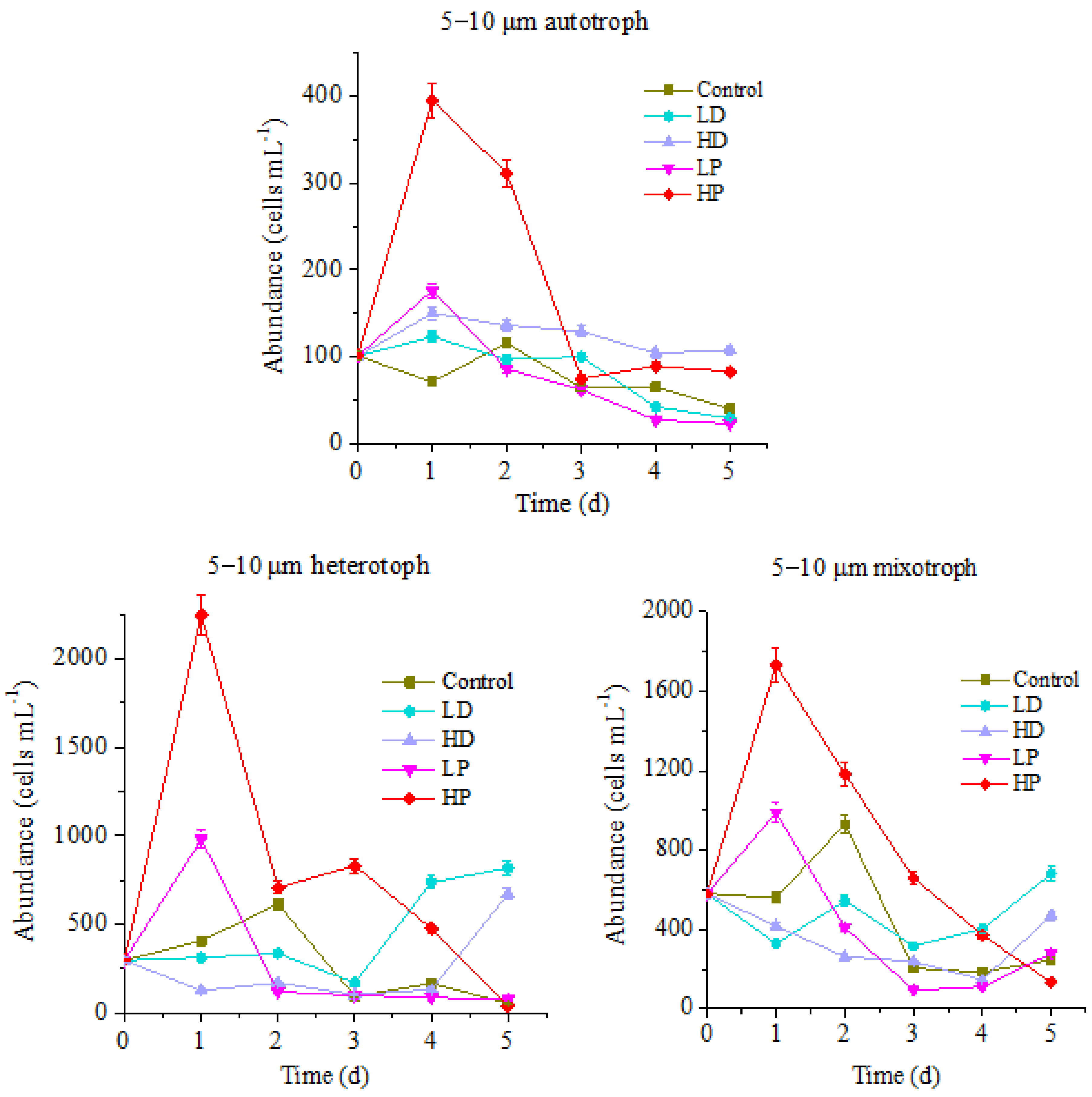
3.1.2. Response of 5–10 µm of Protists to Sand-Dust and Phosphorus Addition
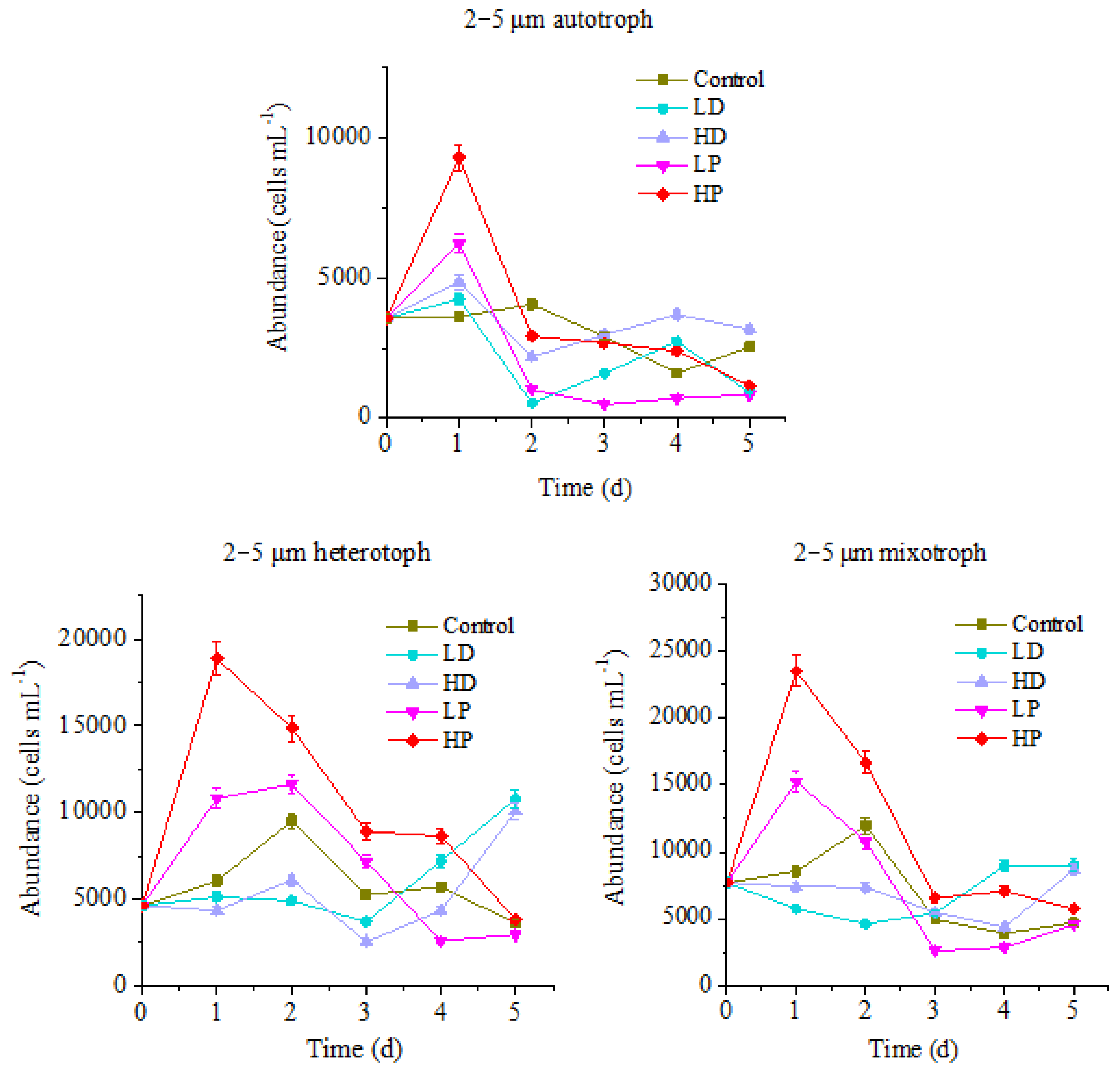
3.1.3. Response of 2–5 µm of Protists to Sand-Dust and Phosphorus Addition
3.2. Effects of Sand-Dust and Phosphorus Addition on the Composition of Protists
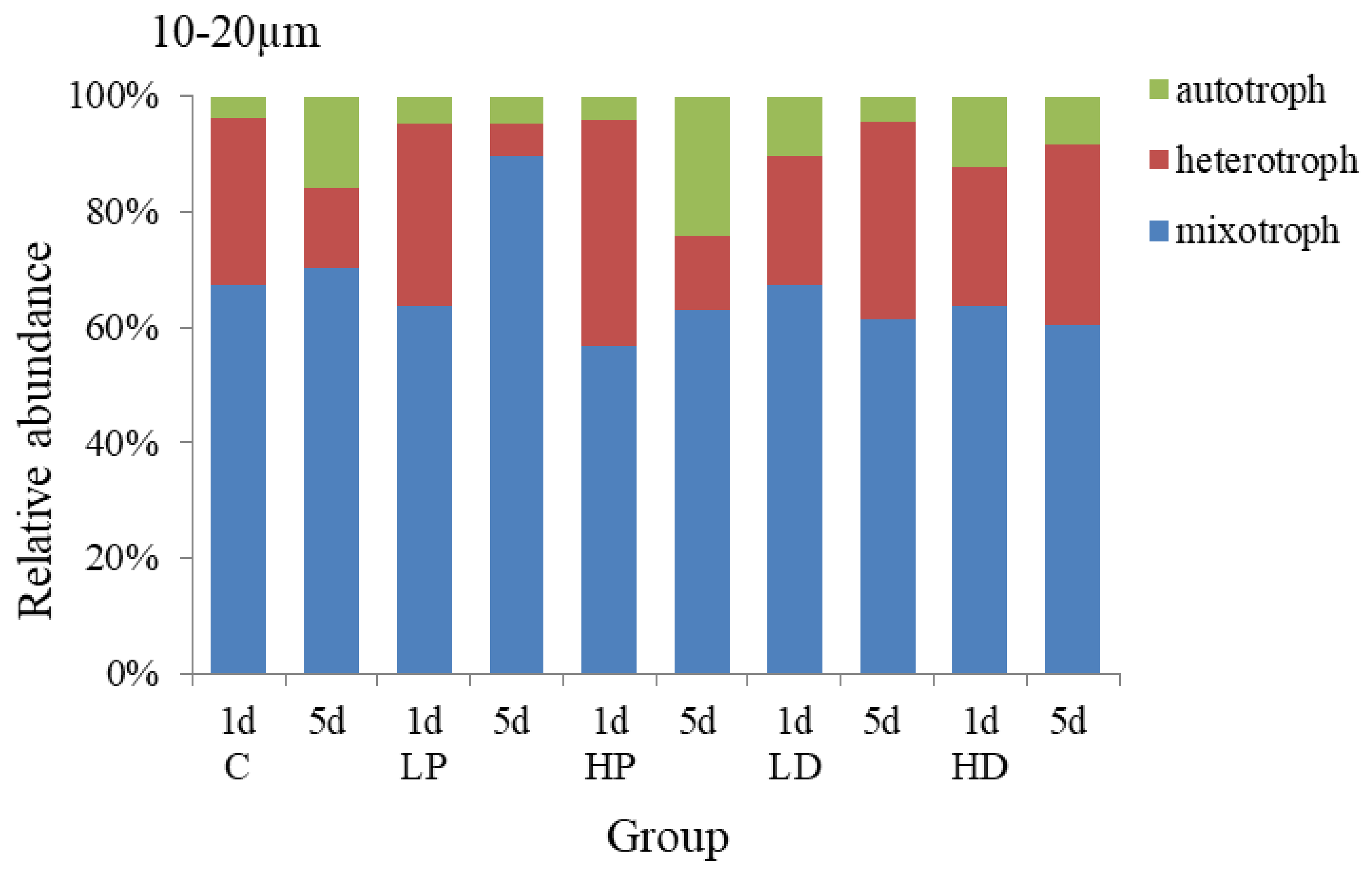
3.2.1. Effects of Dust and Phosphorus Dosage on Trophic Structure of 10–20 µm Protists
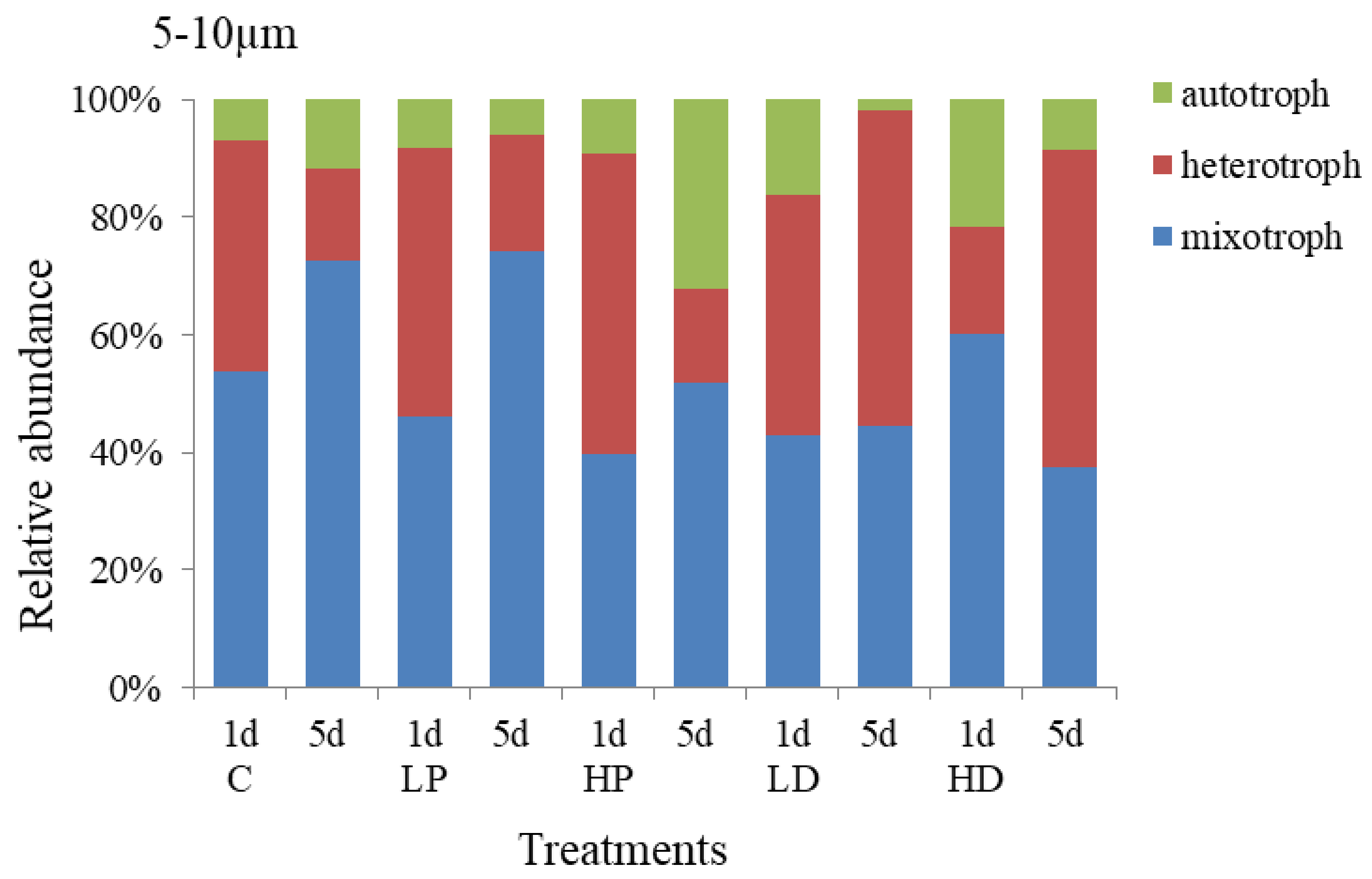
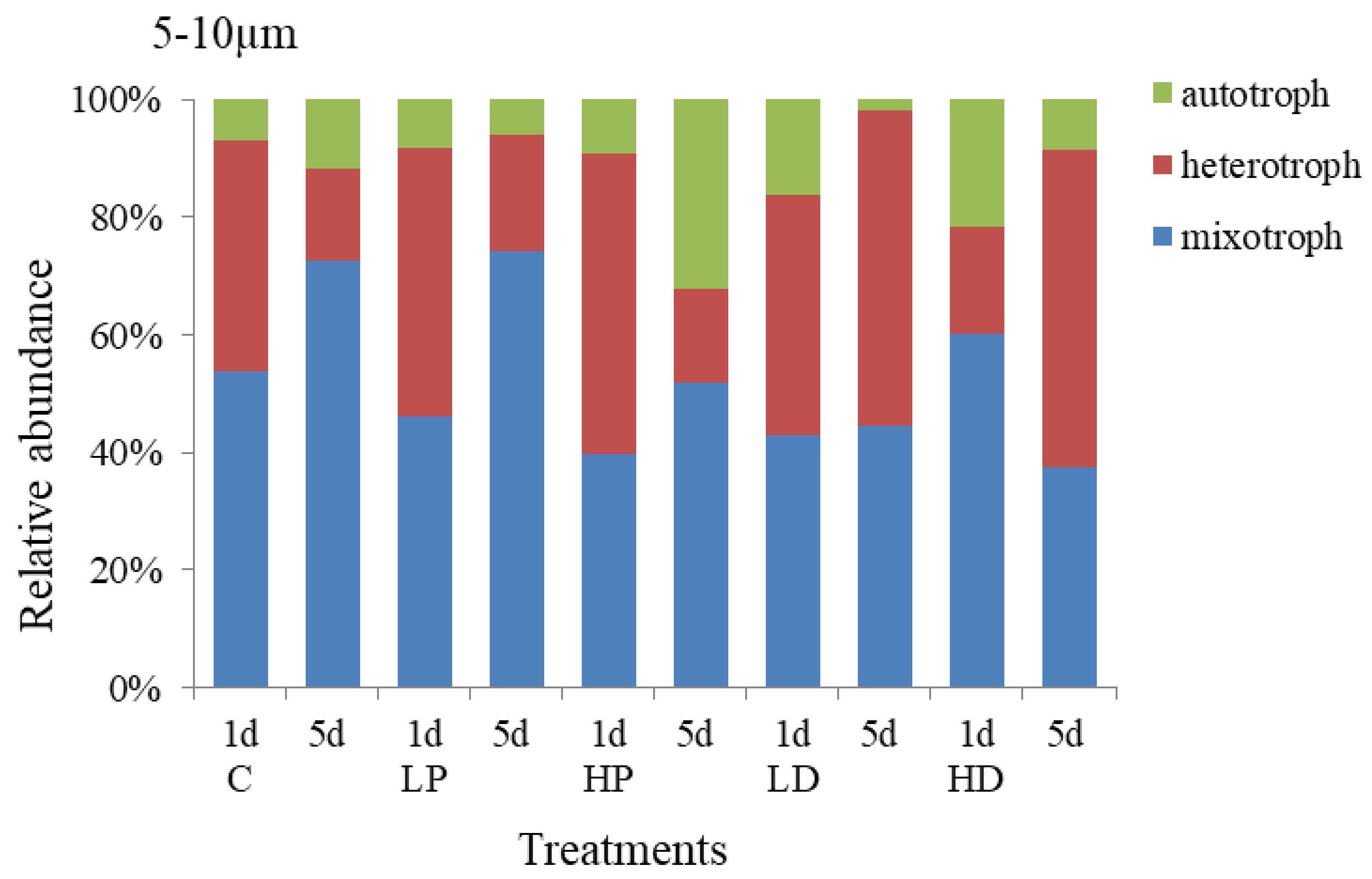
3.2.2. Effects of Dust and Phosphorus Dosage on Trophic Structure of 5–10 µm Protists
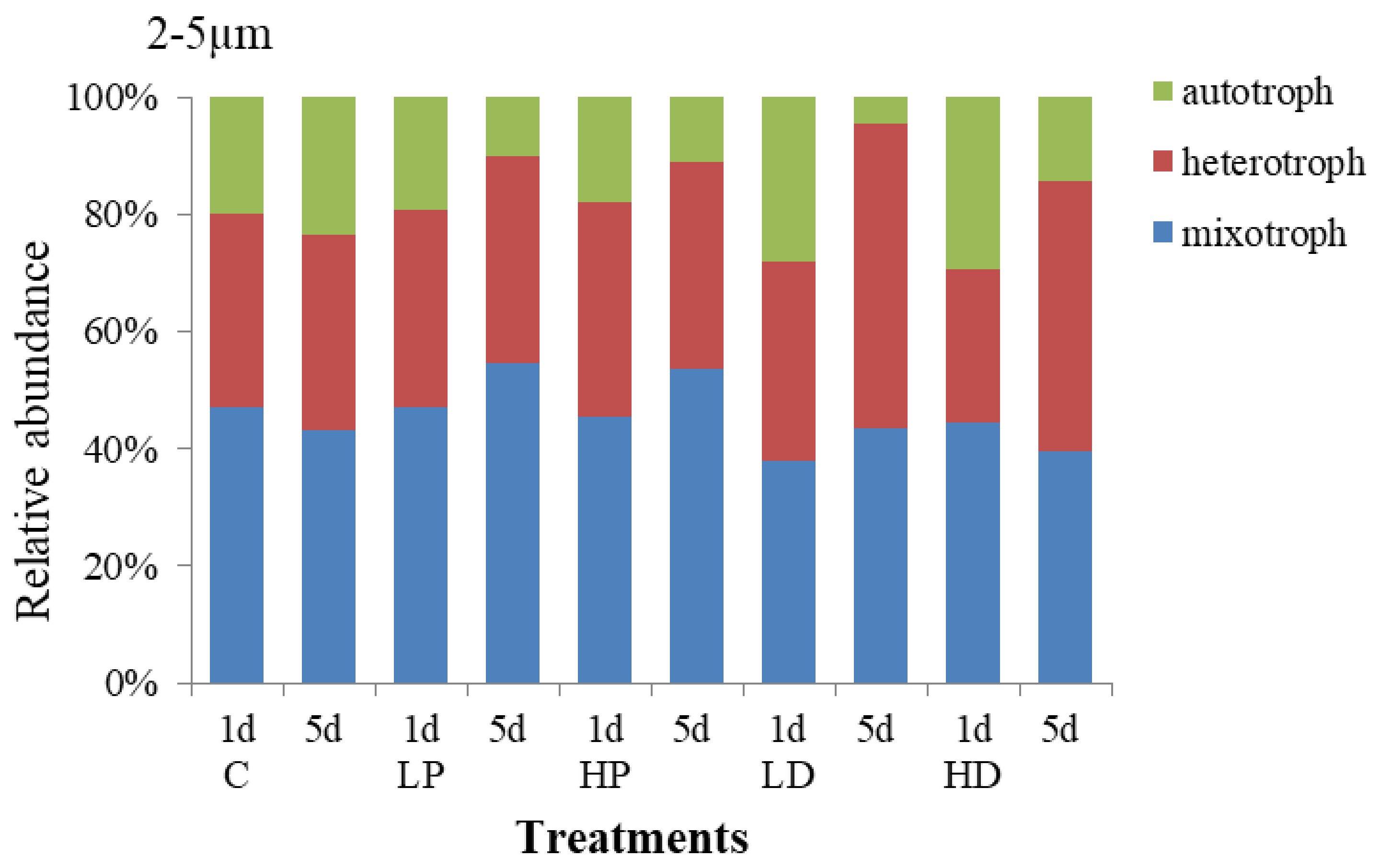
3.2.3. Effects of Dust and Phosphorus Dosage on Trophic Structure of 2–5 µm Protists
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, B.D.; Wang, G.Y. Horizontal distributions and transportation of nutrients in the southern Huanghai Sea. Acta Oceanol. Sin. 1999, 21, 124–129. [Google Scholar]
- Xie, L.P.; Sun, X.; Wang, B.D.; Xin, M. Temporal and spatial distributions of trophic structure and potential nutrient limitation in the Bohai Sea and the Yellow Sea. Mar. Sci. 2012, 36, 45–53. [Google Scholar]
- Ghyoot, C.; Lancelot, C.; Flynn, K.J.; Mitra, A.; Gypens, N. Introducing mixotrophy into a biogeochemical model describing an eutrophied coastal ecosystem. Prog. Oceanogr. 2017, 157, 1–11. [Google Scholar] [CrossRef]
- Stoecker, D.K. Conceptual models of mixotrophy in planktonic protists and some ecological and evolutionary implications. Eur. J. Protistol. 1998, 34, 281–290. [Google Scholar] [CrossRef]
- Griffin, D.W.; Kellogg, C.A. Dust storms and their impact on ocean and human health: Dust in earth’s atmosphere. EcoHealth 2004, 1, 284–295. [Google Scholar] [CrossRef]
- Richon, C.; Dutay, J.; Dulac, F.; Dulac, F.; Wang, R.; Balkanski, Y.; Nabat, P.; Aumont, O.; Desboeufs, K.; Laurent, B.; et al. Modeling the impacts of atmospheric deposition of nitrogen and desert dust-derived phosphorus on nutrients and biological budgets of the Mediterranean Sea. Prog. Oceanogr. 2018, 163, 21–39. [Google Scholar] [CrossRef]
- Zhang, K.; Gao, H.W. The characteristics of Asian-dust storms during 2000–2002: From the source to the sea. Atmos. Environ. 2007, 41, 9136–9145. [Google Scholar]
- Tan, S.C.; Shi, G.Y.; Wang, H. Long-range transport of spring dust storms in Inner Mongolia and impact on the China seas. Atmos. Environ. 2012, 46, 299–308. [Google Scholar] [CrossRef]
- Tan, S.C.; Shi, G.Y.; Shi, J.H.; Gao, H.W.; Yao, X.H. Correlation of Asian dust with chlorophyll and primary productivity in the coastal seas of China during the period from 1998 to 2008. J. Geophys. Res. 2011, 116, G02029. [Google Scholar] [CrossRef]
- Tan, S.C.; Shi, G.Y. The relationship between satellite-derived primary production and vertical mixing and atmospheric inputs in the Yellow Sea cold water mass. Cont. Shelf. Res. 2012, 48, 138–145. [Google Scholar] [CrossRef]
- Tan, S.C.; Wang, H. The transport and deposition of dust and its impact on phytoplankton growth in the Yellow Sea. Atmos. Environ. 2014, 99, 491–499. [Google Scholar] [CrossRef]
- Gao, Y.; Arimoto, R.; Duce, R.A.; Lee, D.S.; Zhou, M.Y. Input of atmospheric trace elements and mineral matter to the Yellow Sea during the spring of a low-dust year. J. Geophys. Res. 1992, 97, 3767–3777. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Hagstrǒm, A.; Azam, F.; Anderson, A.; Wikner, J.; Rassoulzadegan, F. Microbial loop in an oligotrophic pelagic marine ecosystem: Possible roles of cyanobacteria and nanoflagellates in the organic fluxes. Mar. Ecol. Prog. Ser. 1988, 49, 171–178. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Bacterivory and herbivory: Key roles of phagotrophic protists in pelagic food webs. Microb. Ecol. 1994, 28, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Christaki, U.; Courties, C.; Massana, R.; Catala, P.; Lebaron, P.; Gasol, J.M.; Zubkov, M. Optimized routine flow cytometric enumeration of heterotrophic flagellates using SYBR Green I. Limnol. Oceanogr. Meth. 2011, 9, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Tsai, A.Y.; Chiang, K.P. Trophic coupling between Synechococcus and pigmented nanoflagellates in the coastal waters of Taiwan, Western Subtropical Pacific. J. Oceanogr. 2009, 65, 781–789. [Google Scholar]
- Mitra, A.; Flynn, K.J.; Tillmann, U.; Raven, J.A.; Caron, D.; Stoecker, D.K.; Not, F.; Hansen, P.J.; Hallegraeff, G.; Sander, R.W.; et al. Defining planktonic protist functional groups on mechanisms for energy and nutrient acquisition: Incorporation of diverse mixotrophic strategies. Protists 2016, 167, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Sherr, E.B.; Sherr, B.F. Significance of predation by protists in aquatic microbial food webs. Antonie Van Leeuwenhoek 2002, 81, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Stoecker, D.K.; Capuzzo, J.M. Predation on protozoa: Its importance to zooplankton. J. Plankton Res. 1990, 12, 891–908. [Google Scholar] [CrossRef]
- Jeong, H.J.; Song, J.E.; Kang, N.S.; Kim, S.; Yoo, Y.D.; Park, J.Y. Feeding by heterotrophic dinoflagellates on the common marine heterotrophic nanoflagellate Cafeteria sp. Mar. Ecol. Prog. Ser. 2007, 333, 151–160. [Google Scholar] [CrossRef]
- Sato, M.; Yoshikawa, T.; Takeda, S.; Furuya, K. Application of the size-fractionation method to simultaneous estimation of clearance rates by heterotrophic flagellates and ciliates of pico- and nanophytoplankton. J. Exp. Mar. Bio. Ecol. 2007, 349, 334–343. [Google Scholar] [CrossRef]
- Sinistro, R. Top-down and bottom-up regulation of planktonic communities in a warm temperate wetland. J. Plankton Res. 2010, 32, 209–220. [Google Scholar] [CrossRef]
- Cheng, T.T.; Lü, D.R.; Chen, H.B.; Wang, G.C. Size distribution and element composition of dust aerosol in Chinese Otindag Sandland. Chin. Sci. Bull. 2005, 50, 788–792. [Google Scholar] [CrossRef]
- Duan, J.C.; Tan, J.H. Atmospheric heavy metals and Arsenic in China: Situation, sources and control policies. Atmos. Environ. 2013, 4, 93–101. [Google Scholar] [CrossRef]
- Sherr, B.F.; Sherr, E.B. Proportional distribution of total numbers, biovolume, and bacterivory among size classes of 2-20µm nonpigmented marine flagellates. Mar. Microb. Food Webs 1991, 5, 227–237. [Google Scholar]
- Christaki, U.; Courties, C.; Joux, F.; Jeffrey, W.H.; Neveux, J.; Naudin, J. Community structure and trophic role of ciliates and heterotrophic nanoflagellates in Rhone River diluted mesoscale structures (NW Mediterranean Sea). Aquat. Microb. Ecol. 2009, 57, 263–277. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Flynn, K.J.; Burkholder, J.; Berge, T.; Calber, A.; Raven, J.A.; Granéli, E.; Gilvert, P.M.; Hansen, P.J.; Stoecker, D.K.; et al. The role of mixotrophic protists in the biological carbon pump. Biogeosciences 2014, 11, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- Caron, D.A.; Alexander, C.H.; Allen, A.E.; Archibald, J.M.; Arrmbrust, E.V.; Bachy, C.; Bell, C.J.; Bharti, A.; Dyhrman, S.T.; Guida, S.M.; et al. Probing the evolution, ecology and physiology of marine protists using transcriptomics. Nat. Rev. Microbiol. 2017, 15, 6–20. [Google Scholar] [CrossRef]
- Stibor, H.; Stockenreiter, M.; Nejstgaard, J.C.; Ptacnik, R.; Sommer, U. Trophic switches in pelagic systems. Curr. Opin. Syst. Biol. 2019, 13, 108–114. [Google Scholar] [CrossRef]
- Guieu, C.; Dulac, F.; Desboeufs, K.; Wagener, T.; Pulido-Villena, E.; Grisoni, J.; Louis, F.; Ridame, C.; Blain, S.; Brunet, C.; et al. Large clean mesocosms and simulated dust deposition: A new methodology to investigate responses of marine oligotrophic ecosystems to atmospheric inputs. Biogeosciences 2010, 7, 2765–2784. [Google Scholar] [CrossRef]
- Shi, J.H.; Gao, H.W.; Zhang, J.; Tan, S.C.; Ren, J.L.; Liu, C.G.; Liu, Y.; Yao, X.H. Examination of causative link between a spring bloom and dry/wet deposition of Asian dust in the Yellow Sea. Chin. J. Geophys. Res. 2012, 117, D17304. [Google Scholar] [CrossRef]
- Zubkov, M.V.; Burkill, P.H.; Topping, J.N. Flow cytometric enumeration of DNA-stained oceanic planktonic protists. J. Plankton Res. 2007, 29, 79–86. [Google Scholar] [CrossRef]
- Knap, A.; Michaels, A.; Close, A. Protocols for the Joint Global Ocean Flux Study (JGOFS) Core Measurement. In Scientific Committee on Oceanic Research Manual and Guides; UNESCO: Paris, France, 1994; Volume 29, pp. 210–243. [Google Scholar]
- Pearce, I.; Davidson, A.T.; Thomson, P.G.; Wright, S.; Enden, R. Marine microbial ecology in the sub-Antarctic Zone: Rates of bacterial and phytoplankton growth and grazing by heterotrophic protists. Deep Sea Res. Part II 2011, 58, 2248–2259. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Nutrients and DOC (μmol mg−1) | Dissolved Trace Metals (μg g−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NO3− | NO2− | NH4+ | PO43− | DOC | Fe | Cu | Zn | Pb | As | Ni | Cd | Cr | |
| Concentration | 0.53 | 0.03 | 0.01 | 0.004 | 7.20 | 473.12 | 0.23 | 4.27 | 0.24 | 0.09 | 7.08 | 0.04 | 0.13 |
| Groups | Treatment | Concentration |
|---|---|---|
| Control | none | none |
| Low dust group (LD) | dust | 2 mg/L |
| High dust group (HD) | dust | 20 mg/L |
| Low phosphorus group (LP) | NaH2PO4 | 0.2 μmol/L |
| High phosphorus group (HP) | NaH2PO4 | 1 μmol/L |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Liu, G.-X.; Huang, X.; Chen, H.-J.; Zhang, C.; Zhao, Y.-G. Effects of Asian Dust and Phosphorus Input on Abundance and Trophic Structure of Protists in the Southern Yellow Sea. Water 2019, 11, 1188. https://0-doi-org.brum.beds.ac.uk/10.3390/w11061188
Chen X, Liu G-X, Huang X, Chen H-J, Zhang C, Zhao Y-G. Effects of Asian Dust and Phosphorus Input on Abundance and Trophic Structure of Protists in the Southern Yellow Sea. Water. 2019; 11(6):1188. https://0-doi-org.brum.beds.ac.uk/10.3390/w11061188
Chicago/Turabian StyleChen, Xi, Guang-Xing Liu, Xiao Huang, Hong-Ju Chen, Chao Zhang, and Yang-Guo Zhao. 2019. "Effects of Asian Dust and Phosphorus Input on Abundance and Trophic Structure of Protists in the Southern Yellow Sea" Water 11, no. 6: 1188. https://0-doi-org.brum.beds.ac.uk/10.3390/w11061188