
Potential for Natural Attenuation of Domestic and Agricultural Pollution in Karst Groundwater Environments

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
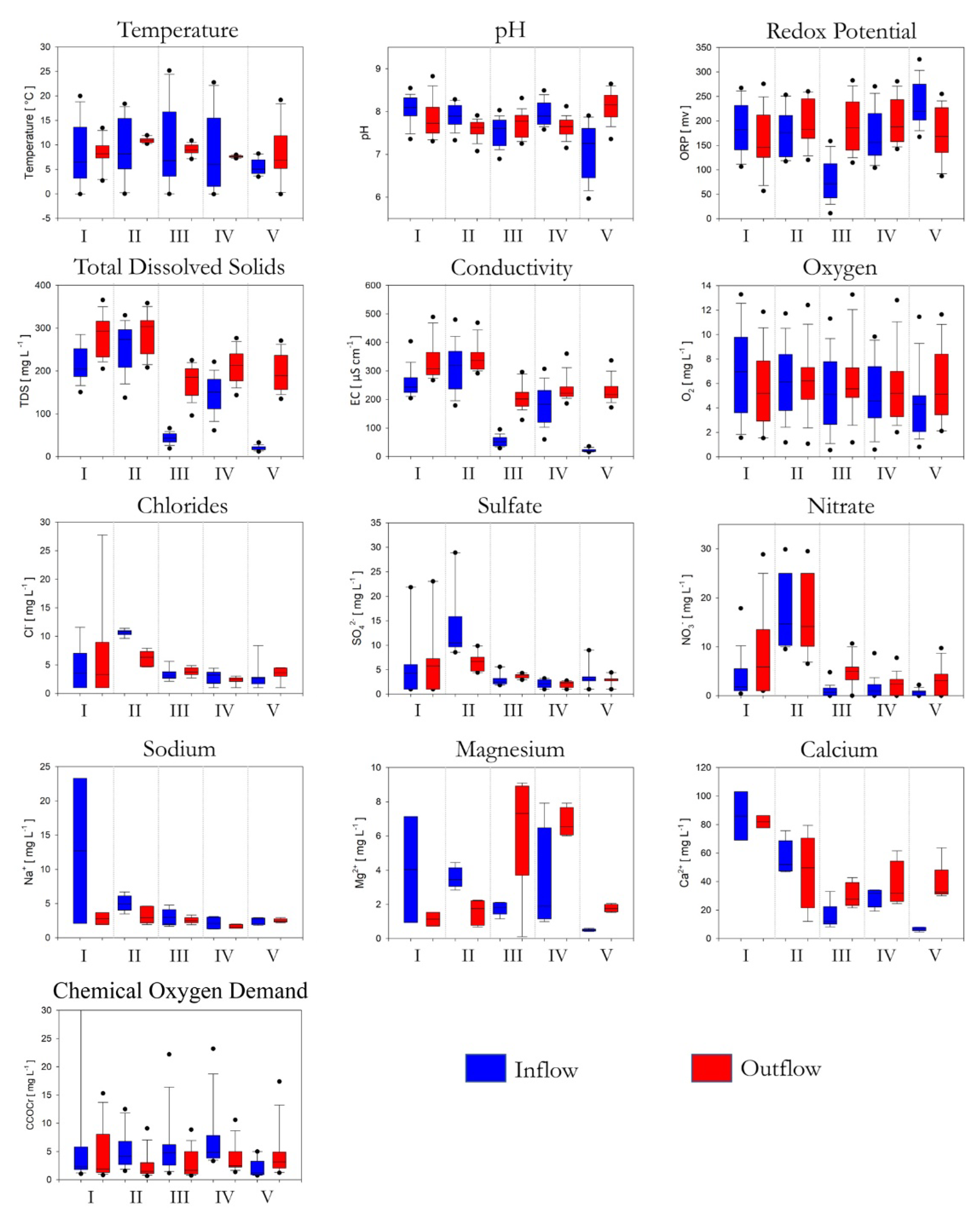
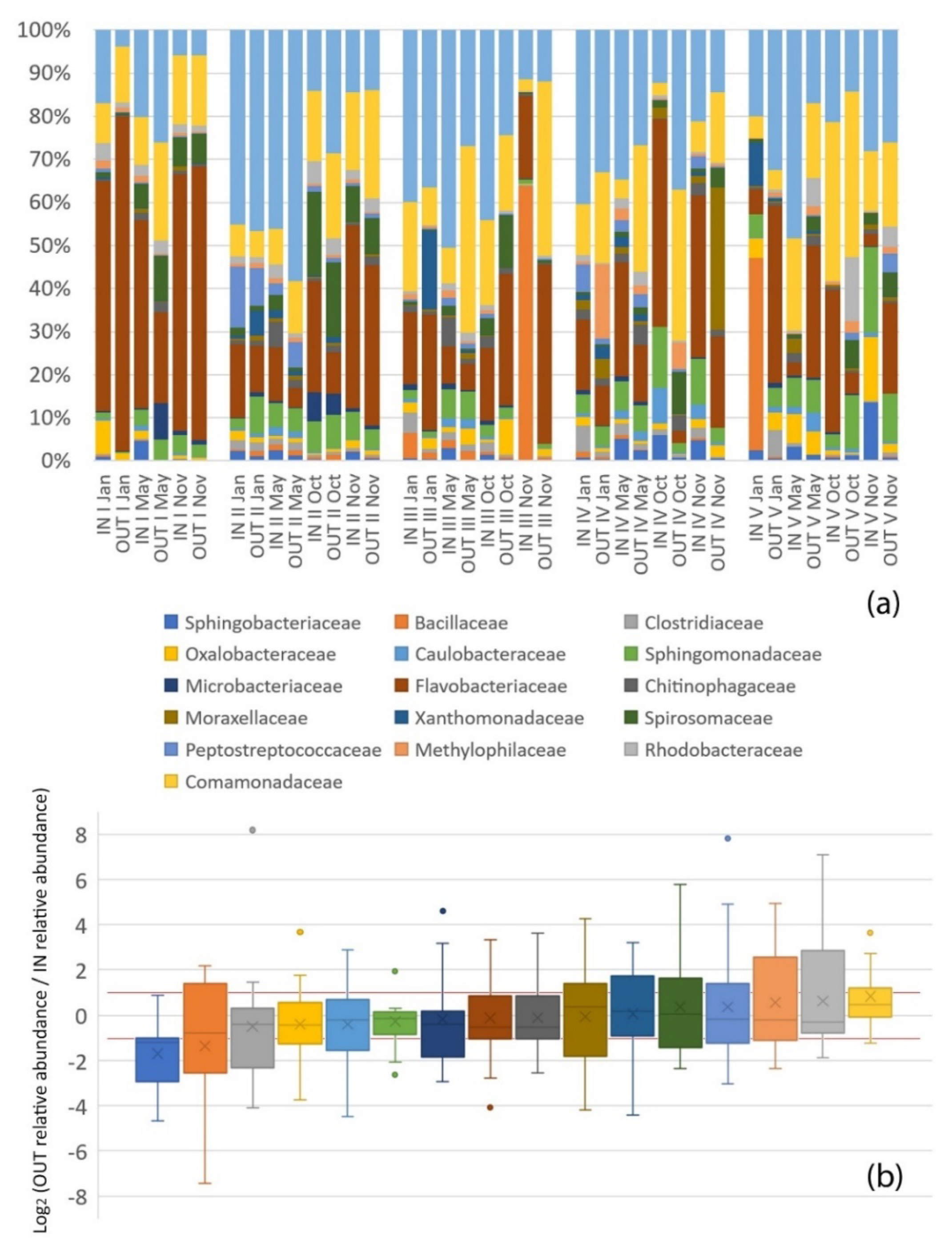
3.1. Environmental Characteristics
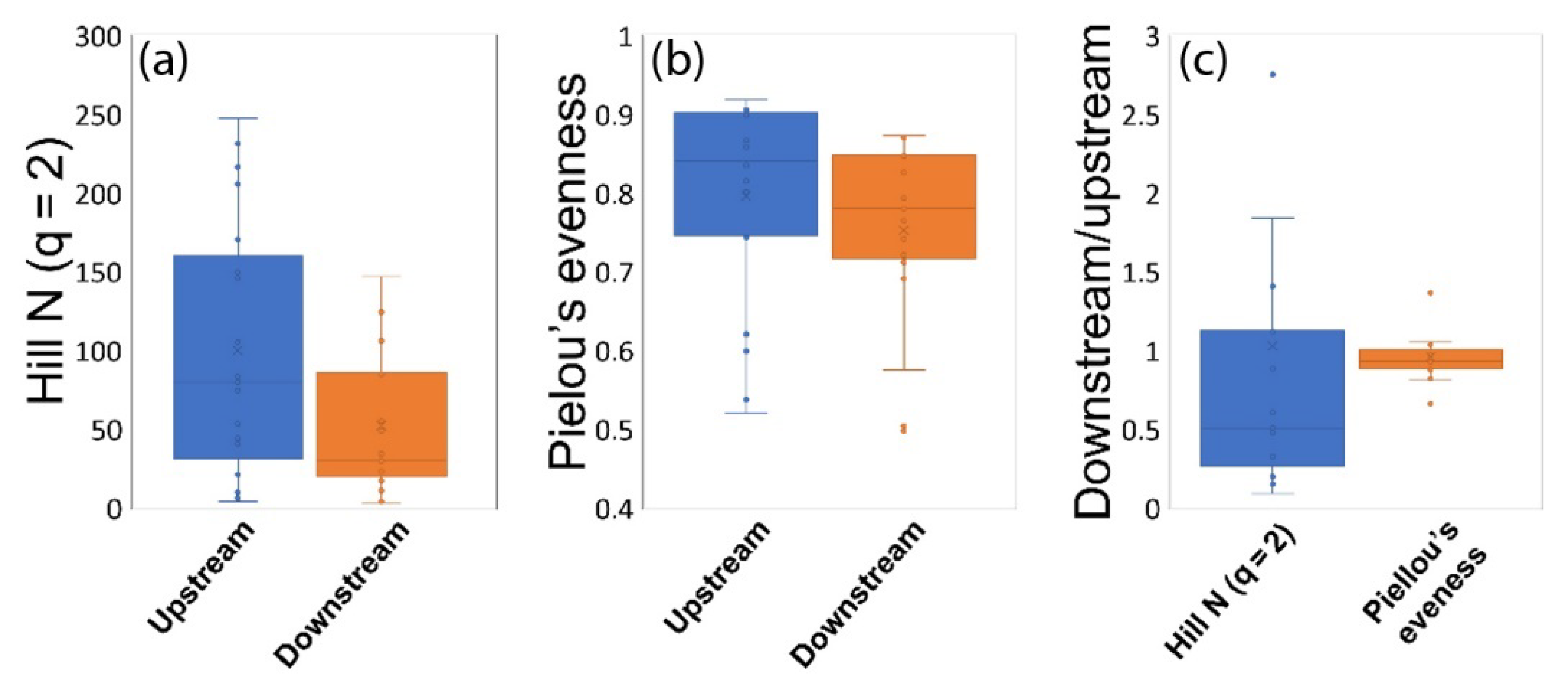
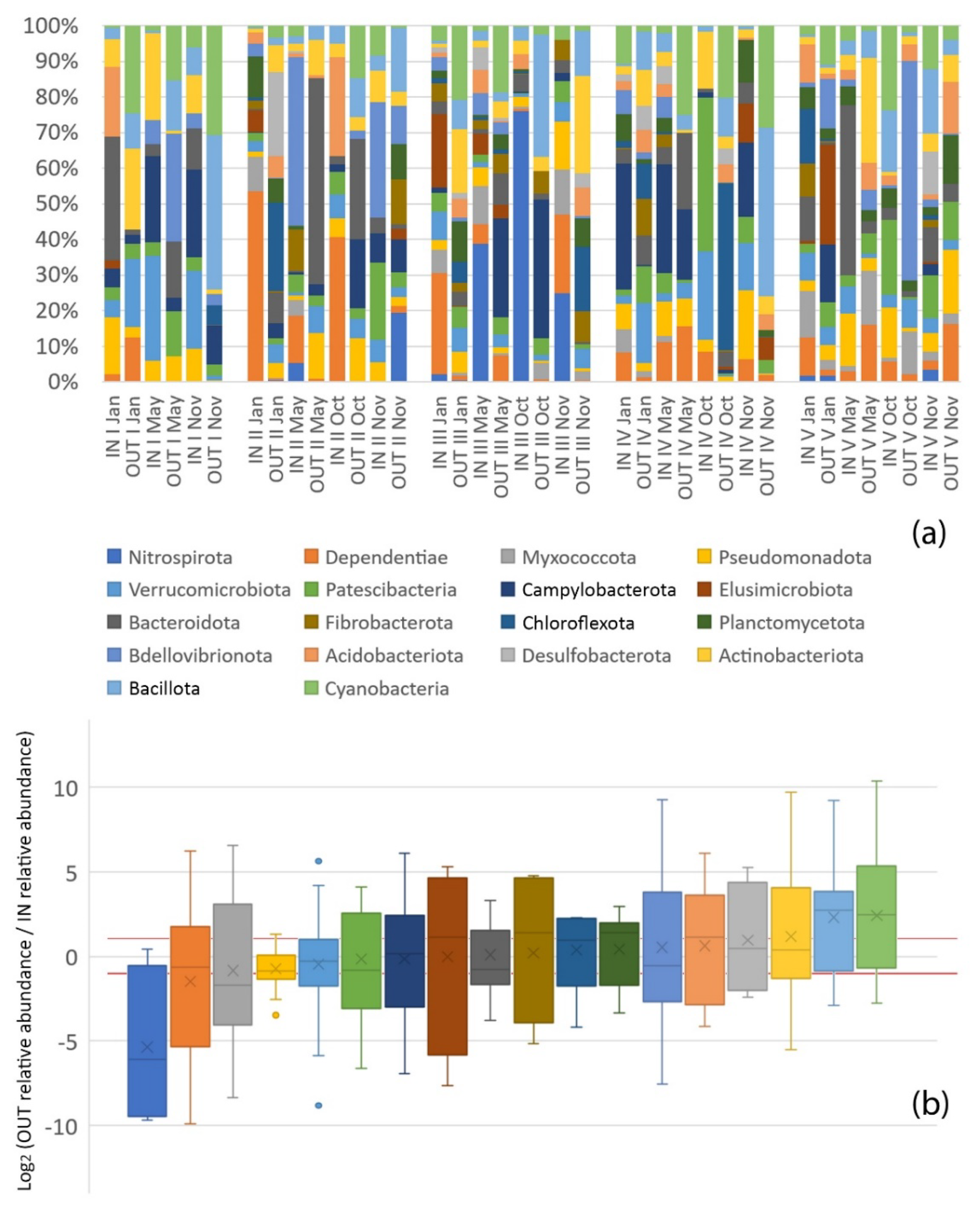
3.2. Microbial Diversity and Community Structure
4. Discussion
4.1. Water Chemistry
4.2. Microbial Community Structure
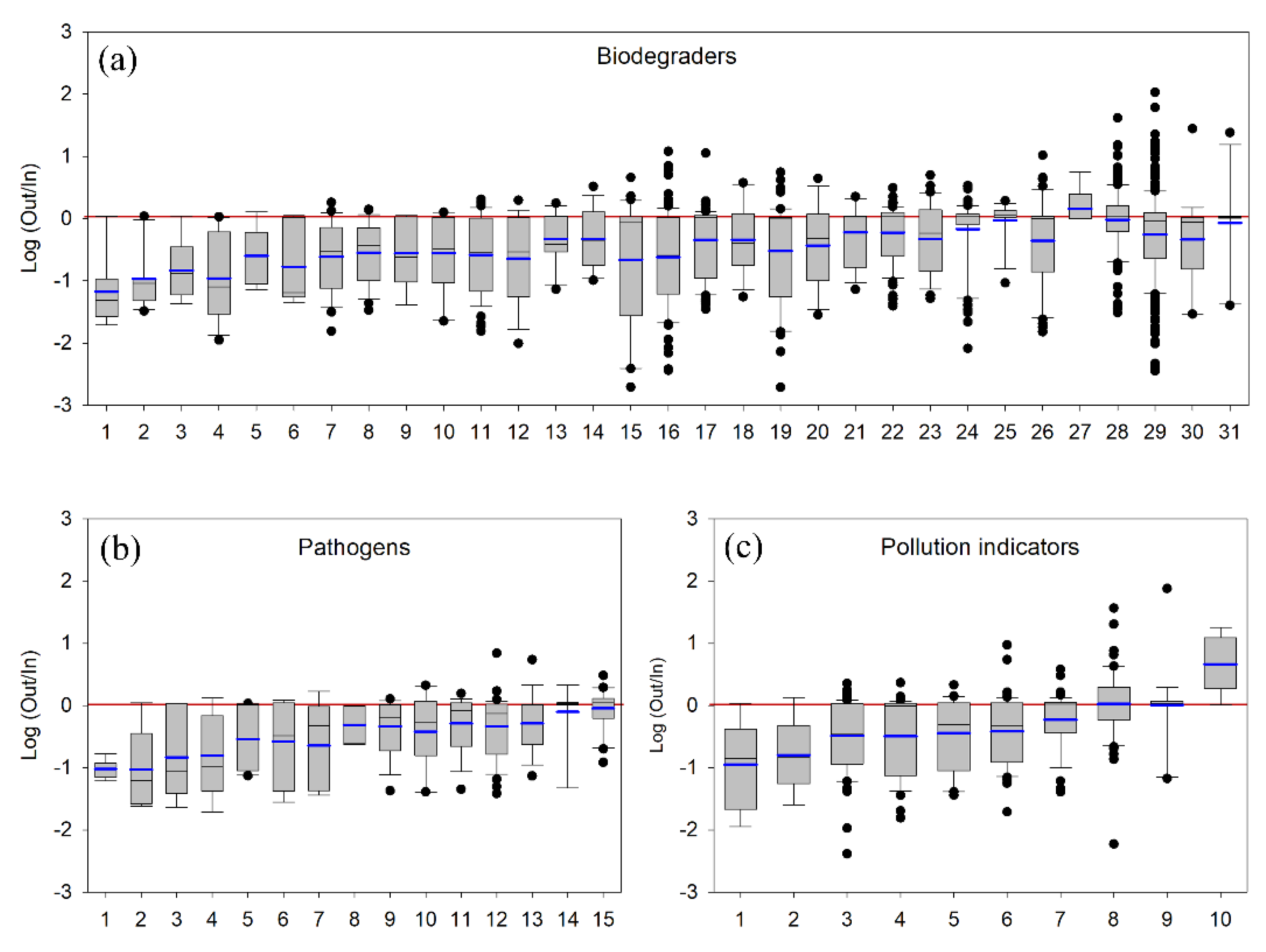
4.2.1. Pollutant-Degrading Microorganisms
4.2.2. Pollution Indicators
4.2.3. Pathogenic Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.; Liu, H.; Xiong, S.; Zeng, F.; Bu, J.; Zhang, B.; Liu, W.; Zhou, H.; Qi, S.; Xu, L.; et al. Rapid transport of organochlorine pesticides (OCPs) in multimedia environment from karst area. Sci. Total Environ. 2021, 775, 145698. [Google Scholar] [CrossRef] [PubMed]
- Brad, T.; Fekete, A.; Sandor, M.S.; Purcarea, C. Natural attenuation potential of selected hydrokarst systems in the Carpathian Mountains (Romania). Water Sci. Technol. Water Supply 2015, 15, 196–206. [Google Scholar] [CrossRef]
- Orăşeanu, I.; Iurkiewicz, A. Karst Hydrogeology of Romania; Belvedere: Oradea, Romania, 2010. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Van Breukelen, B.M.; Röling, W.F.M.; Groen, J.; Griffioen, J.; Van Verseveld, H.W. Biogeochemistry and isotope geochemistry of a landfill leachate plume. J. Contam. Hydrol. 2003, 65, 245–268. [Google Scholar] [CrossRef]
- Huebsch, M.; Fenton, O.; Horan, B.; Hennessy, D.; Richards, K.G.; Jordan, P.; Goldscheider, N.; Butscher, C.; Blum, P. Mobilisation or dilution? Nitrate response of karst springs to high rainfall events. Hydrol. Earth Syst. Sci. 2014, 18, 4423–4435. [Google Scholar] [CrossRef] [Green Version]
- Fridrich, B.; Krčmar, D.; Dalmacija, B.; Molnar, J.; Pešić, V.; Kragulj, M.; Varga, N. Impact of wastewater from pig farm lagoons on the quality of local groundwater. Agric. Water Manag. 2014, 135, 40–53. [Google Scholar] [CrossRef]
- Spoelstra, J.; Leal, K.A.; Senger, N.D.; Schiff, S.L.; Post, R. Isotopic Characterization of Sulfate in a Shallow Aquifer Impacted by Agricultural Fertilizer. Ground Water 2021, 59, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Kumar, M. Sulphate contamination in groundwater and its remediation: An overview. Environ. Monit. Assess. 2020, 192, 74. [Google Scholar] [CrossRef] [PubMed]
- Szklarek, S.; Górecka, A.; Wojtal-Frankiewicz, A. The effects of road salt on freshwater ecosystems and solutions for mitigating chloride pollution—A review. Sci. Total Environ. 2022, 805, 150289. [Google Scholar] [CrossRef] [PubMed]
- Röling, W.F.M.; Van Verseveld, H.W. Natural attenuation: What does the subsurface have in store? Biodegradation 2002, 13, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Negoro, S. Biodegradation of nylon oligomers. Appl. Microbiol. Biotechnol. 2000, 54, 461–466. [Google Scholar] [CrossRef]
- Nagata, Y.; Kato, H.; Ohtsubo, Y.; Tsuda, M. Lessons from the genomes of lindane-degrading sphingomonads. Environ. Microbiol. Rep. 2019, 11, 630–644. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Xiang, F.; Zhao, L.; Qiao, Z. Activated Sludge Microbial Community and Treatment Performance of Wastewater Treatment Plants in Industrial and Municipal Zones. Int. J. Environ. Res. Public Health 2020, 17, 436. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fang, W.; Li, X.; Lu, W.; Li, J. Strong linkages between dissolved organic matter and the aquatic bacterial community in an urban river. Water Res. 2020, 184, 116089. [Google Scholar] [CrossRef]
- Ke, X.; Liu, C.; Tang, S.-Q.; Guo, T.-T.; Pan, L.; Xue, Y.-P.; Zheng, Y.-G. Characterization of Acinetobacter indicus ZJB20129 for heterotrophic nitrification and aerobic denitrification isolated from an urban sewage treatment plant. Bioresour. Technol. 2022, 347, 126423. [Google Scholar] [CrossRef]
- Bogan, B.W.; Sullivan, W.R.; Kayser, K.J.; Derr, K.D.; Aldrich, H.C.; Paterek, J.R. Alkanindiges illinoisensis gen. nov., sp. nov., an obligately hydrocarbonoclastic, aerobic squalane-degrading bacterium isolated from oilfield soils. Int. J. Syst. Evol. Microbiol. 2003, 53, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- De Gusseme, B.; Vanhaecke, L.; Verstraete, W.; Boon, N. Degradation of acetaminophen by Delftia tsuruhatensis and Pseudomonas aeruginosa in a membrane bioreactor. Water Res. 2011, 45, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.R.; de Gannes, V.; Obi, C.C.; Lucas, S.; Lapidus, A.; Cheng, J.-F.; Goodwin, L.A.; Pitluck, S.; Peters, L.; Mikhailova, N.; et al. Complete genome sequence of the phenanthrene-degrading soil bacterium Delftia acidovorans Cs1-4. Stand. Genom. Sci. 2015, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Huang, H.; Ling, Z.; Yu, Z.; Jiang, Y.; Liu, P.; Li, X. Genome sequencing reveals mechanisms for heavy metal resistance and polycyclic aromatic hydrocarbon degradation in Delftia lacustris strain LZ-C. Ecotoxicology 2016, 25, 234–247. [Google Scholar] [CrossRef]
- Leibeling, S.; Schmidt, F.; Jehmlich, N.; von Bergen, M.; Müller, R.H.; Harms, H. Declining Capacity of Starving Delftia acidovorans MC1 to Degrade Phenoxypropionate Herbicides Correlates with Oxidative Modification of the Initial Enzyme. Environ. Sci. Technol. 2010, 44, 3793–3799. [Google Scholar] [CrossRef]
- Liu, Y.; Tie, B.; Li, Y.; Lei, M.; Wei, X.; Liu, X.; Du, H. Inoculation of soil with cadmium-resistant bacterium Delftia sp. B9 reduces cadmium accumulation in rice (Oryza sativa L.) grains. Ecotoxicol. Environ. Saf. 2018, 163, 223–229. [Google Scholar] [CrossRef]
- Johnston, C.W.; Wyatt, M.A.; Li, X.; Ibrahim, A.; Shuster, J.; Southam, G.; Magarvey, N.A. Gold biomineralization by a metallophore from a gold-associated microbe. Nat. Chem. Biol. 2013, 9, 241–243. [Google Scholar] [CrossRef]
- Hou, Y.; Li, B.; Feng, G.; Zhang, C.; He, J.; Li, H.; Zhu, J. Responses of bacterial communities and organic matter degradation in surface sediment to Macrobrachium nipponense bioturbation. Sci. Total Environ. 2021, 759, 143534. [Google Scholar] [CrossRef]
- Danczak, R.E.; Johnston, M.D.; Kenah, C.; Slattery, M.; Wrighton, K.C.; Wilkins, M.J. Members of the Candidate Phyla Radiation are functionally differentiated by carbon- and nitrogen-cycling capabilities. Microbiome 2017, 5, 112. [Google Scholar] [CrossRef]
- Hubalek, V.; Wu, X.; Eiler, A.; Buck, M.; Heim, C.; Dopson, M.; Bertilsson, S.; Ionescu, D. Connectivity to the surface determines diversity patterns in subsurface aquifers of the Fennoscandian shield. ISME J. 2016, 10, 2447–2458. [Google Scholar] [CrossRef]
- Fernando, P.-S.; Alejandro, A.-R.; Monike, O.; Miriam, G.-V.; Mercedes, M.-P.; Yolanda, B.; Nuria, R.; Laurence, B.; Lincoln, S.A.; Fernando, T.; et al. Viable cyanobacteria in the deep continental subsurface. Proc. Natl. Acad. Sci. USA 2018, 115, 10702–10707. [Google Scholar] [CrossRef] [Green Version]
- Kuritz, T.; Wolk, C.P. Use of filamentous cyanobacteria for biodegradation of organic pollutants. Appl. Environ. Microbiol. 1995, 61, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Bestawy, E.A.; El-Salam, A.Z.A.; Mansy, A.E.R.H. Potential use of environmental cyanobacterial species in bioremediation of lindane-contaminated effluents. Int. Biodeterior. Biodegrad. 2007, 59, 180–192. [Google Scholar] [CrossRef]
- Cohen, R.R.H. Use of microbes for cost reduction of metal removal from metals and mining industry waste streams. J. Clean. Prod. 2006, 14, 1146–1157. [Google Scholar] [CrossRef]
- Vijayakumar, S. Potential Applications of Cyanobacteria in Industrial Effluents—A Review. J. Bioremediat. Biodegrad. 2012, 3, 154. [Google Scholar] [CrossRef]
- Forlani, G.; Pavan, M.; Gramek, M.; Kafarski, P.; Lipok, J. Biochemical bases for a widespread tolerance of cyanobacteria to the phosphonate herbicide glyphosate. Plant Cell Physiol. 2008, 49, 443–456. [Google Scholar] [CrossRef]
- Lipok, J.; Wieczorek, D.; Jewgiński, M.; Kafarski, P. Prospects of in vivo31P NMR method in glyphosate degradation studies in whole cell system. Enzyme Microb. Technol. 2009, 44, 11–16. [Google Scholar] [CrossRef]
- Sarmah, P.; Rout, J. Chapter 22—Role of Algae and Cyanobacteria in Bioremediation: Prospects in Polyethylene Biodegradation; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K.B.T.-A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 333–349. ISBN 978-0-12-819311-2. [Google Scholar]
- Chaillan, F.; Gugger, M.; Saliot, A.; Couté, A.; Oudot, J. Role of cyanobacteria in the biodegradation of crude oil by a tropical cyanobacterial mat. Chemosphere 2006, 62, 1574–1582. [Google Scholar] [CrossRef]
- Hiraishi, A.; Hoshino, Y.; Satoh, T. Rhodoferax fermentans gen. nov., sp. nov., a phototrophic purple nonsulfur bacterium previously referred to as the “Rhodocyclus gelatinosus-like” group. Arch. Microbiol. 1991, 155, 330–336. [Google Scholar] [CrossRef]
- Madigan, M.T.; Jung, D.O.; Woese, C.R.; Achenbach, L.A. Rhodoferax antarcticus sp. nov., a moderately psychrophilic purple nonsulfur bacterium isolated from an Antarctic microbial mat. Arch. Microbiol. 2000, 173, 269–277. [Google Scholar] [CrossRef]
- Finneran, K.T.; Johnsen, C.V.; Lovley, D.R. Rhodoferax ferrireducens sp. nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the reduction of Fe (III). Int. J. Syst. Evol. Microbiol. 2003, 53, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrity, G.M. Genus II, Duganella. In Bergey’s Manual of Systematic Bacteriology; Springer Science & Business Media: Hoboken, NJ, USA, 2001. [Google Scholar]
- Chung, B.S.; Ryu, S.H.; Park, M.; Jeon, Y.; Chung, Y.R.; Jeon, C.O. Hydrogenophaga caeni sp. nov., isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2007, 57, 1126–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, J.H.; Kim, J.S.; Lee, D.H.; Jeon, J.H.; Heo, S.Y.; Seo, J.W.; Kim, C.H.; Park, D.S.; Oh, B.R. Thermomonas aquatica sp. nov., isolated from an industrial wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 2019, 69, 3399–3404. [Google Scholar] [CrossRef] [PubMed]
- Spring, S.; Wagner, M.; Schumann, P.; Kämpfer, P. Malikia granosa gen. nov., sp. nov., a novel polyhydroxyalkanoate- and polyphosphate-accumulating bacterium isolated from activated sludge, and reclassification of Pseudomonas spinosa as Malikia spinosa comb. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Gnida, A.; Żabczyński, S.; Surmacz-Górska, J. Filamentous bacteria in the nitrifying activated sludge. Water Sci. Technol. 2018, 77, 2709–2713. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.N.; Frigon, D.; Raskin, L. Populations related to Alkanindiges, a novel genus containing obligate alkane degraders, are implicated in biological foaming in activated sludge systems. Environ. Microbiol. 2007, 9, 1898–1912. [Google Scholar] [CrossRef] [PubMed]
- Fahrbach, M.; Kuever, J.; Remesch, M.; Huber, B.E.; Kämpfer, P.; Dott, W.; Hollender, J. Steroidobacter denitrificans gen. nov., sp. nov., a steroidal hormone-degrading gammaproteobacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2215–2223. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P.H.; de Muro, M.A.; Nielsen, J.L. Studies on the in situ physiology of Thiothrix spp. present in activated sludge. Environ. Microbiol. 2000, 2, 389–398. [Google Scholar] [CrossRef]
- Henriet, O.; Meunier, C.; Henry, P.; Mahillon, J. Filamentous bulking caused by Thiothrix species is efficiently controlled in full-scale wastewater treatment plants by implementing a sludge densification strategy. Sci. Rep. 2017, 7, 1430. [Google Scholar] [CrossRef]
- Gerritsen, J.; Umanets, A.; Staneva, I.; Hornung, B.; Ritari, J.; Paulin, L.; Rijkers, G.T.; de Vos, W.M.; Smidt, H. Romboutsia hominis sp. nov., the first human gut-derived representative of the genus romboutsia, isolated from ileostoma effluent. Int. J. Syst. Evol. Microbiol. 2018, 68, 3479–3486. [Google Scholar] [CrossRef]
- Bojović, K.; Ignjatović, Ð.D.I.; Soković Bajić, S.; Vojnović Milutinović, D.; Tomić, M.; Golić, N.; Tolinački, M. Gut Microbiota Dysbiosis Associated With Altered Production of Short Chain Fatty Acids in Children With Neurodevelopmental Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Thierry, S.; Macarie, H.; Iizuka, T.; Geissdoerfer, W.; Assih, E.; Verhe, F.; Thomas, P.; Fudou, R.; Monroy, O.; Labat, M.; et al. Pseudoxanthomonas mexicana sp. nov. and Pseudoxanthomonas japonensis sp. nov., isolated from diverse environments, and emended descriptions of the genus Pseudoxanthomonas Finkmann et al. 2000 and of its type species. Int. J. Syst. Evol. Microbiol. 2004, 54, 2245–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finegold, S.M.; Vaisanen, M.-L.; Molitoris, D.R.; Tomzynski, T.J.; Song, Y.; Liu, C.; Collins, M.D.; Lawson, P.A. Cetobacterium somerae sp. nov. from Human Feces and Emended Description of the Genus Cetobacterium. Syst. Appl. Microbiol. 2003, 26, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.L.; Nagaraja, T.G.; Chengappa, M.M. Fusobacterium necrophorum infections: Virulence factors, pathogenic mechanism and control measures. Vet. Res. Commun. 1996, 20, 113–140. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, H.; Chen, S.; Zhou, L.; Li, Y.; Zhao, K.; Huang, F.; Fan, Z.; Xuan, L.; Zhang, X.; et al. Intestinal Microbiota Can Predict Acute Graft-versus-Host Disease Following Allogeneic Hematopoietic Stem Cell Transplantation. Biol. Blood Marrow Transplant. 2019, 25, 1944–1955. [Google Scholar] [CrossRef]
- Yu, X.; Du, Y.; Wang, G. Knoellia flava sp. nov., isolated from pig manure. Int. J. Syst. Evol. Microbiol. 2012, 62, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Cotta, M.A.; Whitehead, T.R.; Collins, M.D.; Lawson, P.A. Atopostipes suicloacale gen. nov., sp. nov., isolated from an underground swine manure storage pit. Anaerobe 2004, 10, 191–195. [Google Scholar] [CrossRef]
- Graf, J. The Family Rikenellaceae BT—The Prokaryotes: Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 857–859. ISBN 978-3-642-38954-2. [Google Scholar]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes; Springer Science & Business Media: Hoboken, NJ, USA, 2011; Volume 3, ISBN 0387684891. [Google Scholar]
- Theelen, M.J.P.; Luiken, R.E.C.; Wagenaar, J.A.; Sloet van Oldruitenborgh-Oosterbaan, M.M.; Rossen, J.W.A.; Zomer, A.L. The Equine Faecal Microbiota of Healthy Horses and Ponies in The Netherlands: Impact of Host and Environmental Factors. Animals 2021, 11, 1762. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [Green Version]
- Ormerod, K.L.; Wood, D.L.A.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.O.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Rajilić-Stojanović, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Mukerji, R.; Kakarala, R.; Smith, S.J.; Kusz, H.G. Chryseobacterium indologenes: An emerging infection in the USA. BMJ Case Rep. 2016, 2016, bcr2016214486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, R.; Pathak, A. Emerging Chryseobacterium indologenes Infection in Indian Neonatal Intensive Care Units: A Case Report. Antibiotics 2018, 7, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina, J.; Gil, J.; Salva, F.; Gomez, J.; Alomar, P. Microbiological characteristics of Weeksella virosa (formerly CDC group IIf) isolated from the human genitourinary tract. J. Clin. Microbiol. 1990, 28, 2357–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Liao, K.; Ai, L.; Guo, P.; Huang, H.; Wu, Z.; Liu, M. Bacteremia caused by Bergeyella zoohelcum in an infective endocarditis patient: Case report and review of literature. BMC Infect. Dis. 2017, 17, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, R.C. Bacillus anthracis. J. Clin. Pathol. 2003, 56, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Kuisiene, N.; Raugalas, J.; Spröer, C.; Kroppenstedt, R.M.; Chitavichius, D. Bacillus butanolivorans sp. nov., a species with industrial application for the remediation of n-butanol. Int. J. Syst. Evol. Microbiol. 2008, 58, 505–509. [Google Scholar] [CrossRef] [Green Version]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Inglett, K.S.; Bae, H.S.; Aldrich, H.C.; Hatfield, K.; Ogram, A.V. Clostridium chromiireducens sp. nov., isolated from Cr(VI)-contaminated soil. Int. J. Syst. Evol. Microbiol. 2011, 61, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Eugenio, G.; Fardeau, M.-L.; Cayol, J.-L.; Patel, B.K.C.; Thomas, P.; Macarie, H.; Garcia, J.-L.; Ollivier, B. Clostridium thiosulfatireducens sp. nov., a proteolytic, thiosulfate- and sulfur-reducing bacterium isolated from an upflow anaerobic sludge blanket (UASB) reactor. Int. J. Syst. Evol. Microbiol. 2002, 52, 1461–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druggan, P.; Iversen, C. Chapter 21: Culture Media for Food Spoilage Bacteria of the Order Pseudomonadales: Pseudomonas, Acinetobacter and Psychrobacter spp. In Handbook of Culture Media for Food and Water Microbiology, 3rd ed.; Royal Society of Chemistry: London, UK, 2011; pp. 482–507. [Google Scholar] [CrossRef]
- Bonwitt, J.; Tran, M.; Droz, A.; Gonzalez, A.; Glover, W.A. Psychrobacter sanguinis wound infection associated with marine environment exposure, Washington, DC, USA. Emerg. Infect. Dis. 2018, 24, 1942–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
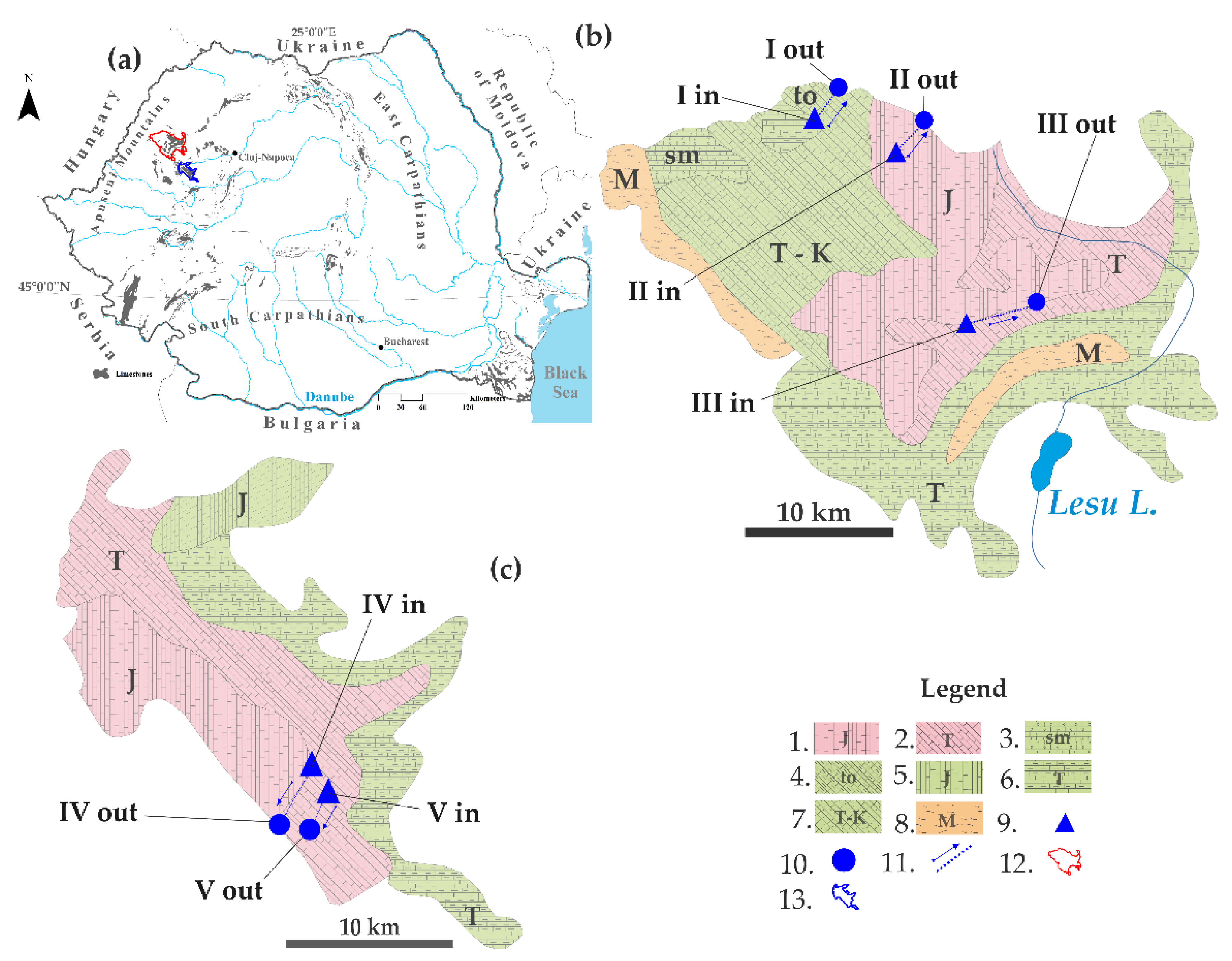
| Mountain | HKS Ponor–Spring (Inflow–Outflow) | Length (m) | Time (h) | H (m) | UT | |
|---|---|---|---|---|---|---|
| Pădurea Craiului | I | Potriva Cave–Aștileu Cave | 2620 | 10 | 107 | 2 |
| II | Gălășeni Cave–Josani Spring | 1750 | 13 | 95 | 0 | |
| III | Toaie Ponor–Dămișenilor Spring | 3550 | 90 | 255 | 2 | |
| Bihor | IV | Ocoale Valley–Cotețul Dobreștilor Spring | 2800 | 388 | 770 | 1 |
| V | Vuiagă well–Poliței Spring | 1360 | 10 | 225 | 1 | |
| Functional Group | Taxon | Average Ratio | Standard Deviation | Highest Abundance | |
|---|---|---|---|---|---|
| 1 | Biodegraders | Bryobacter sp. | 0.18 | 0.36 | Upstream |
| 2 | Biodegraders | Massilia sp. | 0.21 | 0.33 | Upstream |
| 3 | Biodegraders | Clostridium chromiireducens | 0.28 | 0.37 | Upstream |
| 4 | Biodegraders | Pedobacter boryungensis | 0.30 | 0.39 | Upstream |
| 5 | Biodegraders | Methylobacterium sp. | 0.41 | 0.45 | Upstream |
| 6 | Biodegraders | Flavobacterium hauense | 0.41 | 0.50 | Upstream |
| 7 | Biodegraders | Acinetobacter lwoffii | 0.47 | 0.49 | Upstream |
| 8 | Biodegraders | Flavobacterium limicola | 0.49 | 0.42 | Upstream |
| 9 | Biodegraders | Bacteriovorax sp. | 0.50 | 0.48 | Upstream |
| 10 | Biodegraders | Alkanindiges sp. | 0.53 | 0.48 | Upstream |
| 12 | Biodegraders | Parasediminibacterium sp. | 0.56 | 0.59 | Upstream |
| 11 | Biodegraders | Flavobacterium psychrolimnae | 0.56 | 0.57 | Upstream |
| 13 | Biodegraders | Clostridium butyricum | 0.66 | 0.52 | Upstream |
| 14 | Biodegraders | Bradyrhizobium elkanii | 0.80 | 0.88 | Upstream |
| 16 | Biodegraders | Pedobacter sp. | 0.84 | 1.62 | Upstream |
| 21 | Biodegraders | Terrimonas sp. | 0.85 | 0.62 | Upstream |
| 17 | Biodegraders | Sediminibacterium sp. | 0.86 | 1.35 | Upstream |
| 19 | Biodegraders | Flavobacterium saccharophilum | 0.88 | 0.99 | Upstream |
| 18 | Biodegraders | Sphingomonas faeni | 0.89 | 1.10 | Upstream |
| 20 | Biodegraders | Sphingomonas glacialis | 0.89 | 1.24 | Upstream |
| 22 | Biodegraders | Novosphingobium sp. | 0.90 | 0.62 | Upstream |
| 23 | Biodegraders | Flavobacterium segetis | 0.93 | 1.09 | Upstream |
| 26 | Biodegraders | Flavobacterium aquatile | 1.00 | 1.51 | Downstream |
| 24 | Biodegraders | Pseudomonas sp. | 1.02 | 0.68 | Downstream |
| 15 | Biodegraders | Flavobacterium chungangense | 1.09 | 1.64 | Downstream |
| 25 | Biodegraders | Nitrospira sp. | 1.12 | 0.47 | Downstream |
| 27 | Biodegraders | Flavobacterium buctense | 1.93 | 2.03 | Downstream |
| 28 | Biodegraders | Limnohabitans sp. | 1.94 | 3.98 | Downstream |
| 29 | Biodegraders | Flavobacterium succinicans | 2.03 | 7.86 | Downstream |
| 30 | Biodegraders | Flavobacterium pectinovorum | 2.19 | 6.40 | Downstream |
| 31 | Biodegraders | Flavobacterium aquidurense | 3.10 | 6.98 | Downstream |
| 1 | Pathogens | Staphylococcus equorum | 0.10 | 0.04 | Upstream |
| 2 | Pathogens | Chryseobacterium chaponense | 0.25 | 0.42 | Upstream |
| 3 | Pathogens | Carnobacterium maltaromaticum | 0.37 | 0.48 | Upstream |
| 4 | Pathogens | Ralstonia pickettii | 0.38 | 0.48 | Upstream |
| 5 | Pathogens | Facklamia tabacinasalis | 0.49 | 0.45 | Upstream |
| 6 | Pathogens | Carnobacterium inhibens | 0.52 | 0.49 | Upstream |
| 7 | Pathogens | Brevundimonas intermedia | 0.57 | 0.66 | Upstream |
| 8 | Pathogens | Polynucleobacter sp. | 0.60 | 0.40 | Upstream |
| 9 | Pathogens | Paeniclostridium sp. | 0.65 | 0.42 | Upstream |
| 10 | Pathogens | Brevundimonas staleyi | 0.71 | 0.67 | Upstream |
| 11 | Pathogens | Romboutsia sp. | 0.73 | 0.46 | Upstream |
| 12 | Pathogens | Clostridium sp. | 0.82 | 1.13 | Upstream |
| 13 | Pathogens | Clostridium estertheticum | 0.89 | 1.28 | Upstream |
| 14 | Pathogens | Aeromonas media | 1.07 | 0.56 | Downstream |
| 15 | Pathogens | Polynucleobacter asymbioticus | 1.13 | 0.68 | Downstream |
| 1 | Pollution indicators | Ruminiclostridium sp. | 0.29 | 0.41 | Upstream |
| 2 | Pollution indicators | Rhizobacter gummiphilus | 0.36 | 0.55 | Upstream |
| 3 | Pollution indicators | Luteolibacter sp. | 0.64 | 0.56 | Upstream |
| 4 | Pollution indicators | Undibacterium sp. | 0.66 | 0.56 | Upstream |
| 5 | Pollution indicators | Sphaerotilus sp. | 0.66 | 0.58 | Upstream |
| 6 | Pollution indicators | Devosia sp. | 0.76 | 1.35 | Upstream |
| 7 | Pollution indicators | Arenimonas sp. | 0.88 | 0.69 | Upstream |
| 8 | Pollution indicators | Rhodoferax sp. | 1.24 | 2.24 | Downstream |
| 9 | Pollution indicators | Perlucidibaca sp. | 4.97 | 17.19 | Downstream |
| 10 | Pollution indicators | Pseudoxanthomonas mexicana | 6.87 | 6.40 | Downstream |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brad, T.; Bizic, M.; Ionescu, D.; Chiriac, C.M.; Kenesz, M.; Roba, C.; Ionescu, A.; Fekete, A.; Mirea, I.C.; Moldovan, O.T. Potential for Natural Attenuation of Domestic and Agricultural Pollution in Karst Groundwater Environments. Water 2022, 14, 1597. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101597
Brad T, Bizic M, Ionescu D, Chiriac CM, Kenesz M, Roba C, Ionescu A, Fekete A, Mirea IC, Moldovan OT. Potential for Natural Attenuation of Domestic and Agricultural Pollution in Karst Groundwater Environments. Water. 2022; 14(10):1597. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101597
Chicago/Turabian StyleBrad, Traian, Mina Bizic, Danny Ionescu, Cecilia Maria Chiriac, Marius Kenesz, Carmen Roba, Artur Ionescu, Alexandru Fekete, Ionut Cornel Mirea, and Oana Teodora Moldovan. 2022. "Potential for Natural Attenuation of Domestic and Agricultural Pollution in Karst Groundwater Environments" Water 14, no. 10: 1597. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101597