
Characterizing Spatial Neighborhoods of Refugia Following Large Fires in Northern New Mexico USA


Abstract
:1. Introduction
2. Materials and Methods
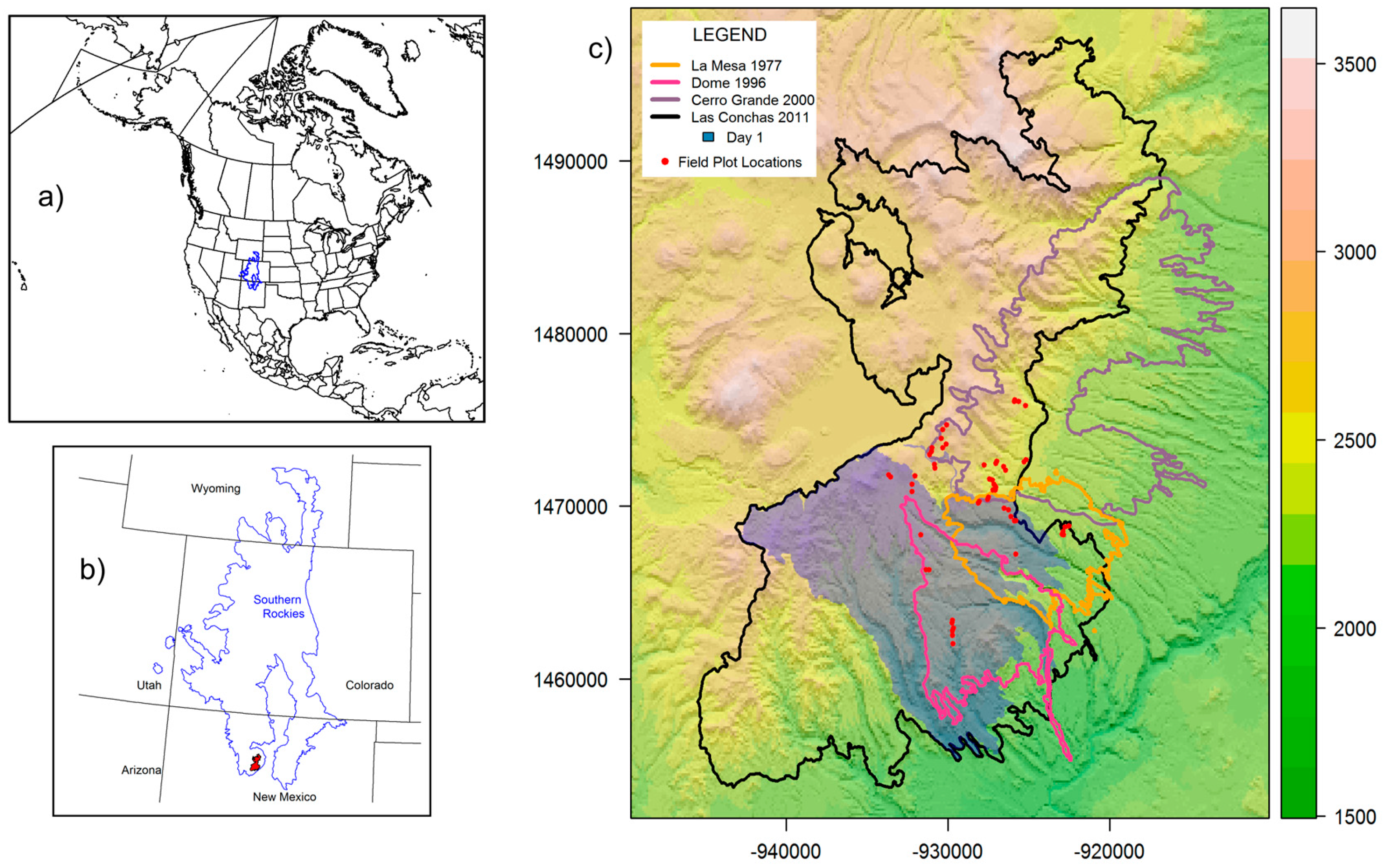
2.1. Study Area
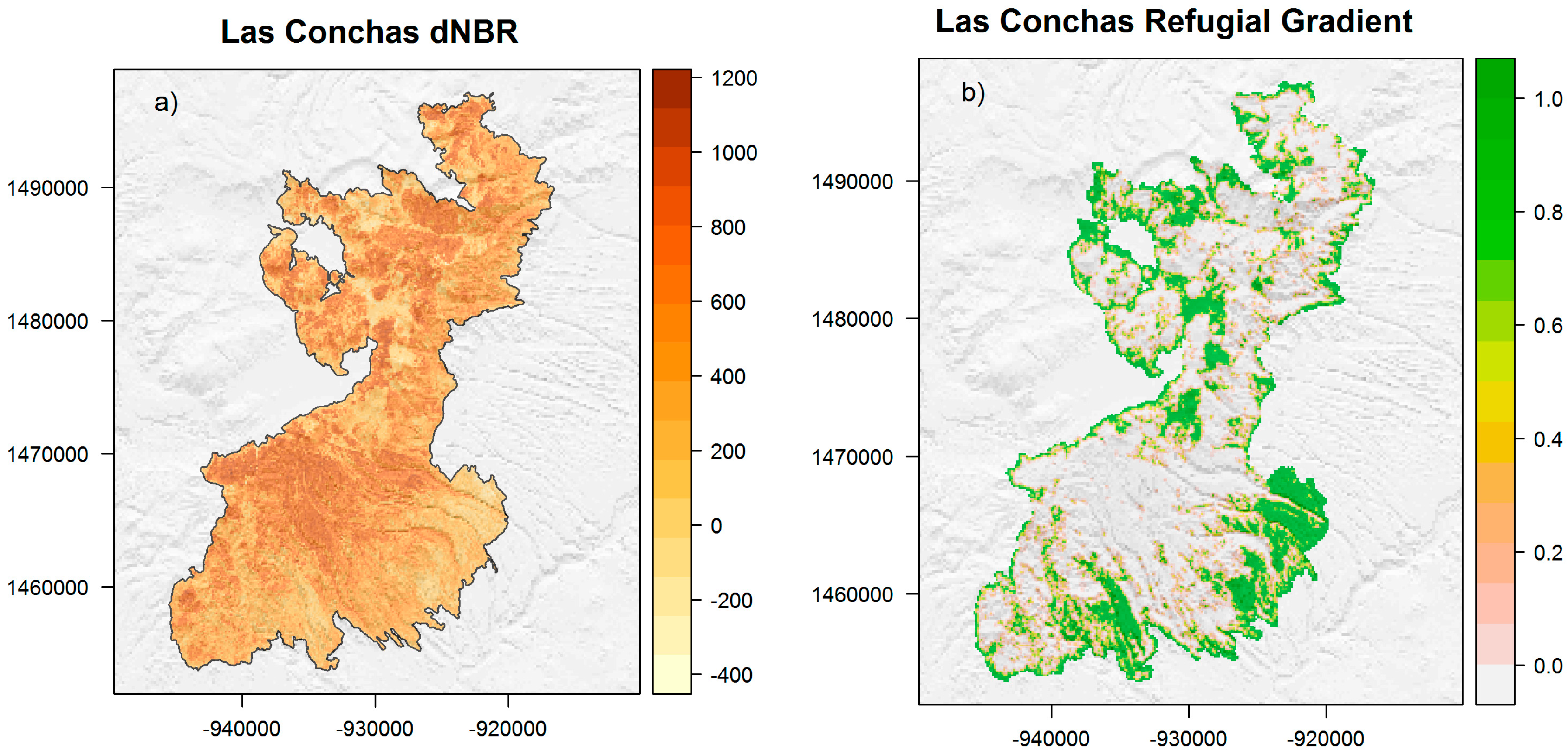
2.2. Las Conchas RG
2.3. Refugial Environments
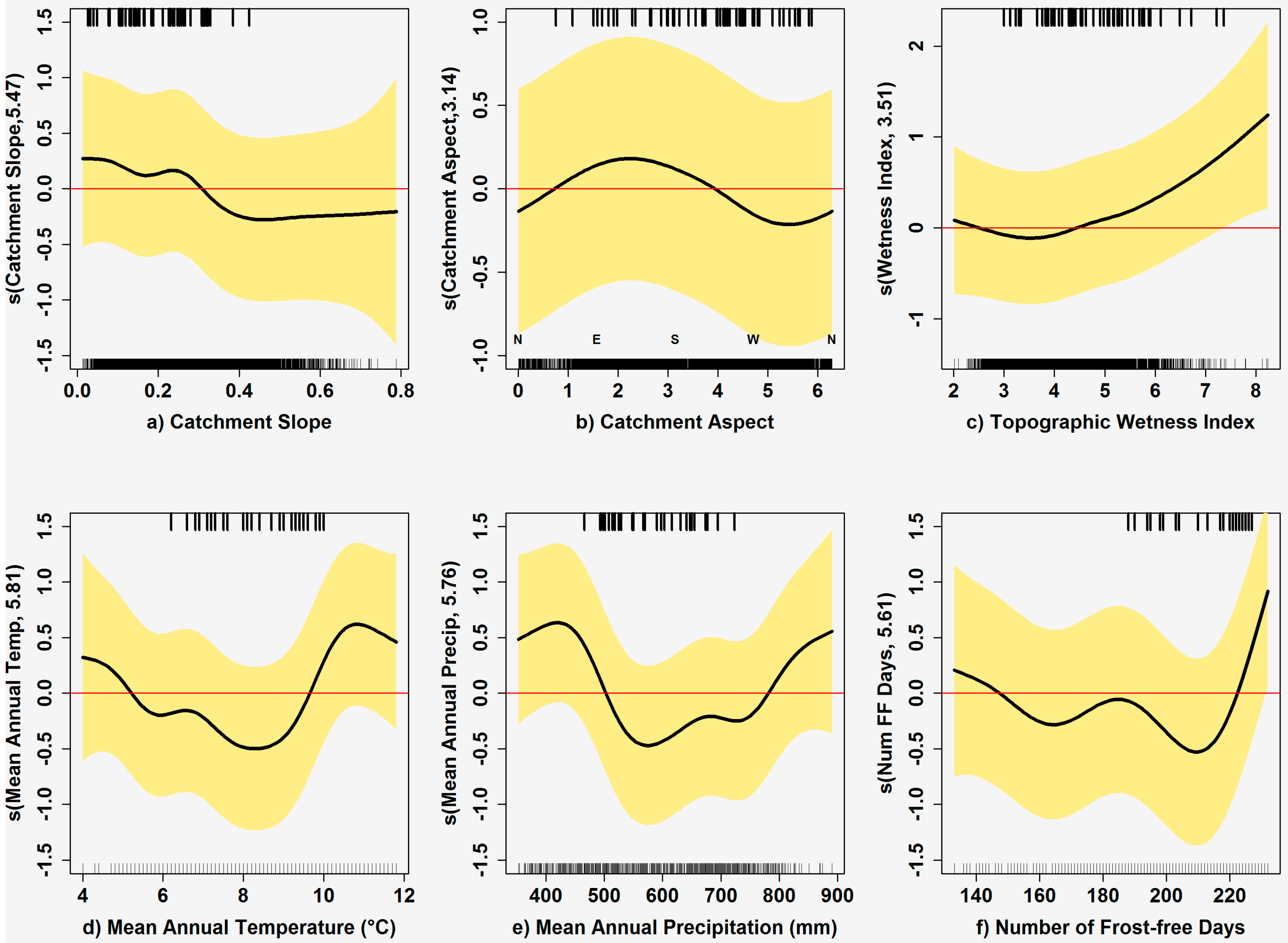
2.4. Modeling Plant Communities
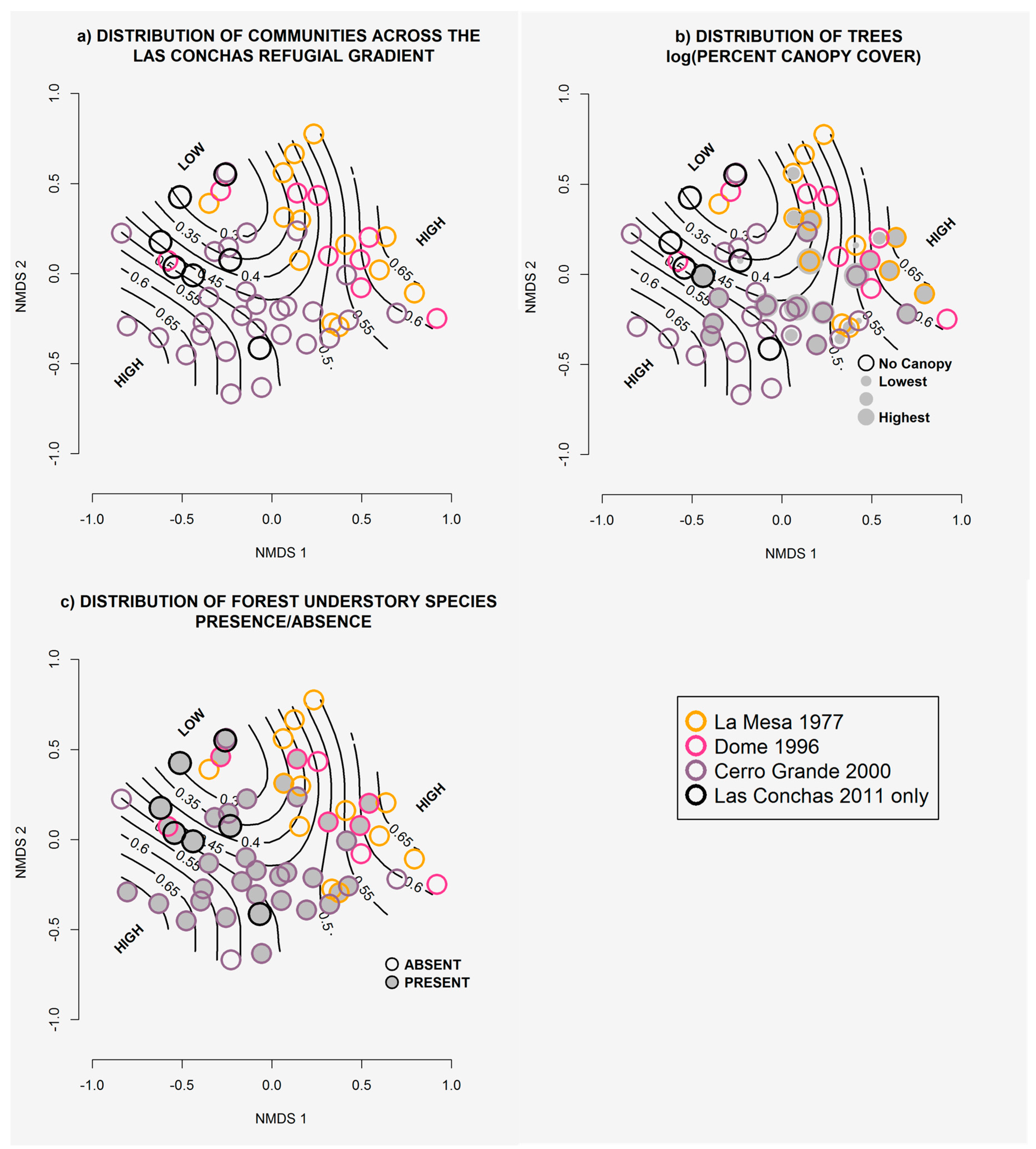
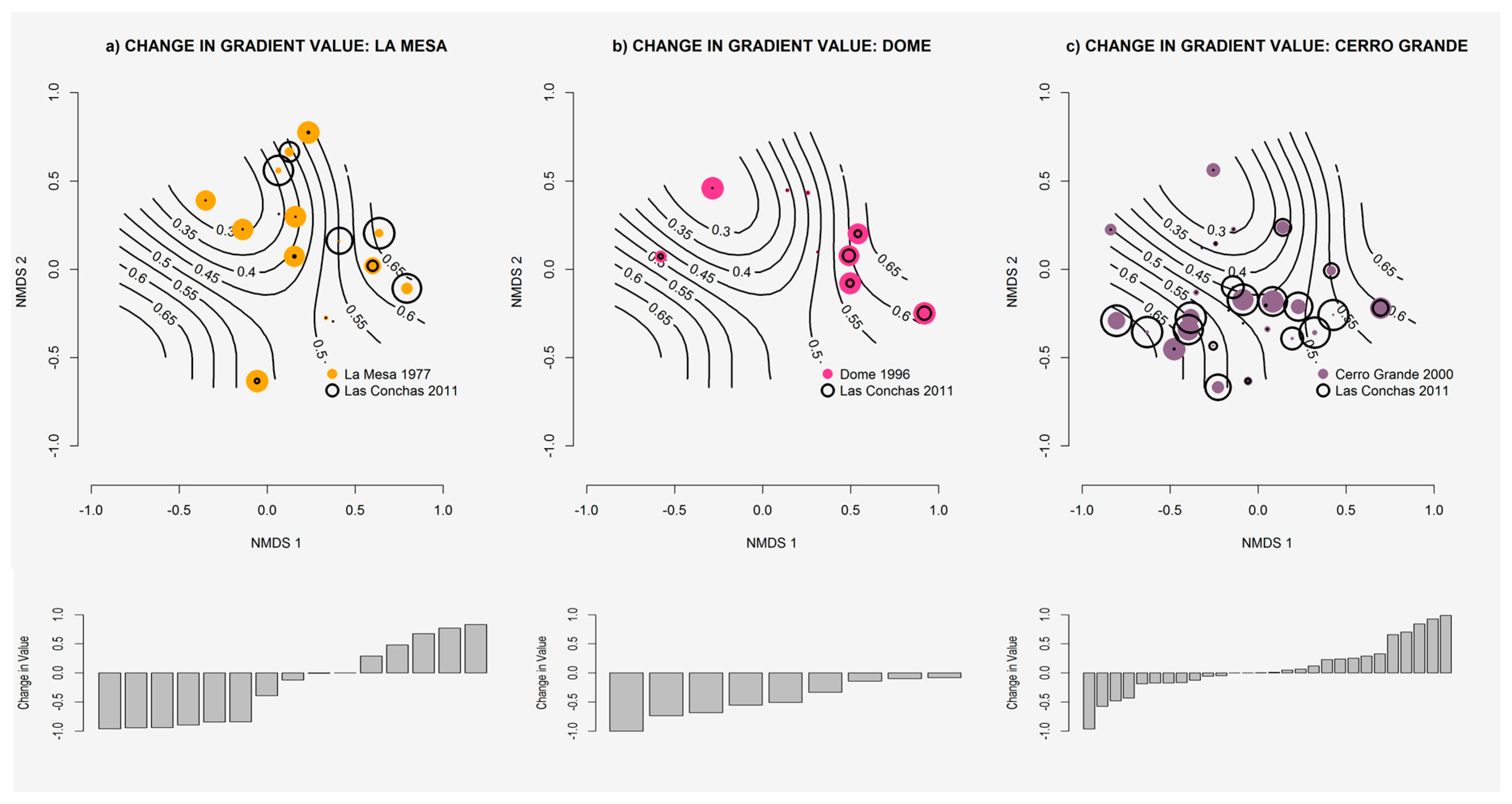
2.5. Gradient Dynamics
3. Results
3.1. Las Conchas RG
3.2. Refugial Environments
3.3. Community Models
3.4. Gradient Dynamics
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Foxx and Hoard 1995 Class | Frequency (%) | Gradient Value Mean (min, max) | Gradient Mean Class | |
|---|---|---|---|---|---|---|
| Allium geyeri | Geyer’s onion | herbaceous | showy monocots | 0.02 | 0.83 (0.83, 0.83) | high |
| Artemisia carruthii | Carruth’s sagewort | herbaceous | composites | 0.07 | 0.8 (0.35, 1) | high (+) |
| Artemisia dracunculus | tarragon | herbaceous | composites | 0.09 | 0.75 (0.08, 1) | high (+) |
| Grindelia squarrosa | curly-cup gumweed | herbaceous | composites | 0.02 | 0.72 (0.72, 0.72) | high (+) |
| Hymenoxys acaulis * | perky sue | herbaceous | composites | 0.02 | 1 (1, 1) | high (+) |
| Solidago nemoralis var. decemflora | goldenrod | herbaceous | composites | 0.02 | 0.72 (0.72, 0.72) | high (+) |
| Andropogon gerardii | big bluestem | herbaceous | grasses | 0.07 | 0.72 (0.09, 1) | high (+/−) |
| Elymus spp. | wildrye | herbaceous | grasses | 0.02 | 0.93 (0.93, 0.93) | high |
| Festuca thurberi | Thurber’s fescue | herbaceous | grasses | 0.09 | 0.79 (0.04, 1) | high (+/−) |
| Lycurus setosus | bristly wolfstail | herbaceous | grasses | 0.02 | 1 (1, 1) | high |
| Carex praegracilis | clustered field sedge | herbaceous | rushes and sedges | 0.05 | 1 (1, 1) | high (+) |
| Arenaria lanuginosa ssp. saxosa | spreading sandwort | herbaceous | showy dicots | 0.04 | 0.97 (0.94, 1) | high |
| Campanula rotundifolia | bluebell bellflower | herbaceous | showy dicots | 0.12 | 0.8 (0.05, 1) | high |
| Erigeron formosissimus | beautiful fleabane | herbaceous | composites | 0.02 | 1 (1, 1) | high |
| Frasera speciosa | showy frasera | herbaceous | showy dicots | 0.02 | 1 (1, 1) | high (+) |
| Ipomopsis aggregata | skyrocket gilia | herbaceous | showy dicots | 0.02 | 1 (1, 1) | high (+) |
| Lupinus argenteus | silvery lupine | herbaceous | showy dicots | 0.04 | 0.89 (0.83, 0.96) | high |
| Sisymbrium altissimum | tall tumblemustard | herbaceous | showy dicots | 0.02 | 0.81 (0.81, 0.81) | high |
| Trifolium wormskioldii | cows clover | herbaceous | showy dicots | 0.07 | 0.72 (0, 1) | high |
| Abies concolor | white fir | woody | trees | 0.07 | 0.81 (0.49, 1) | high |
| Quercus x pauciloba | wavyleaf oak | woody | shrub | 0.09 | 0.7 (0.02, 1) | high (+) |
| Echinocereus triglochidiatus | kingcup cactus | herbaceous | cacti | 0.02 | 0.45 (0.45, 0.45) | medium (−) |
| Achillea millefolium | common yarrow | herbaceous | composites | 0.46 | 0.4 (0, 1) | medium (+/−) |
| Antennaria spp. | pussytoes | herbaceous | composites | 0.14 | 0.5 (0.01, 1) | medium (+) |
| Antennaria parvifolia | smallleaf pussytoes | herbaceous | composites | 0.04 | 0.4 (0.09, 0.72) | medium (+/−) |
| Brickellia eupatorioides var. chlorolepis | false boneset | herbaceous | composites | 0.07 | 0.61 (0, 1) | medium (+) |
| Brickellia grandiflora | tasselflower brickellbush | herbaceous | composites | 0.04 | 0.58 (0.16, 1) | medium (+) |
| Heterotheca villosa | false goldenaster | herbaceous | composites | 0.23 | 0.58 (0, 1) | medium (+/−) |
| Laennecia schiedeana | pineland marshtail | herbaceous | composites | 0.07 | 0.48 (0.06, 0.97) | medium (+/−) |
| 1 Oreochrysum parryi | Parry’s goldenrod | herbaceous | composites | 0.19 | 0.49 (0, 1) | medium (+) |
| Solidago missouriensis | Missouri goldenrod | herbaceous | composites | 0.07 | 0.4 (0, 0.72) | medium (+) |
| Bouteloua gracilis | blue grama | herbaceous | grasses | 0.09 | 0.46 (0, 1) | medium (+/−) |
| Danthonia parryi | Parry’s oatgrass | herbaceous | grasses | 0.18 | 0.5 (0, 1) | medium (+/−) |
| Elymus elymoides | bottlebrush squirreltail | herbaceous | grasses | 0.30 | 0.49 (0, 1) | medium (+/−) |
| Elymus trachycaulus | slender wheatgrass | herbaceous | grasses | 0.18 | 0.55 (0, 1) | medium (+) |
| Koeleria macrantha | prairie junegrass | herbaceous | grasses | 0.32 | 0.46 (0, 1) | medium (+/−) |
| Muhlenbergia montana | mountain muhly | herbaceous | grasses | 0.42 | 0.5 (0, 1) | medium (+/−) |
| Muhlenbergia ramulosa | green muhly | herbaceous | grasses | 0.02 | 0.45 (0.45, 0.45) | medium (−) |
| Poa fendleriana | muttongrass | herbaceous | grasses | 0.30 | 0.56 (0, 1) | medium (+/−) |
| Carex spp.—upland small | sm. upland sedge <25 cm | herbaceous | rushes and sedges | 0.12 | 0.65 (0.01, 1) | medium (+/−) |
| 1 Carex occidentalis | western sedge | herbaceous | rushes and sedges | 0.09 | 0.63 (0, 1) | medium (+/−) |
| 1 Carex siccata | dryspike sedge | herbaceous | rushes and sedges | 0.25 | 0.48 (0, 1) | medium (+/−) |
| Arenaria fendleri | Fendler’s sandwort | herbaceous | showy dicots | 0.04 | 0.59 (0.35, 0.83) | medium |
| Castilleja miniata | scarlet Indian paintbrush | herbaceous | showy dicots | 0.04 | 0.5 (0.04, 0.96) | medium (−) |
| Erigeron divergens | spreading fleabane | herbaceous | composites | 0.02 | 0.45 (0.45, 0.45) | medium (−) |
| Erigeron flagellaris | trailing fleabane | herbaceous | composites | 0.12 | 0.43 (0, 1) | medium (−) |
| Erigeron subtrinervis | threenerve fleabane | herbaceous | composites | 0.05 | 0.56 (0.04, 0.83) | medium (−) |
| Erysimum capitatum | sanddune wallflower | herbaceous | showy dicots | 0.09 | 0.43 (0, 0.97) | medium (+/−) |
| 1 Fragaria vesca | woodland strawberry | herbaceous | showy dicots | 0.09 | 0.44 (0, 1) | medium (−) |
| Galium aparine | stickywilly | herbaceous | showy dicots | 0.04 | 0.42 (0.04, 0.81) | medium (−) |
| Galium boreale | Northern bedstraw | herbaceous | showy dicots | 0.04 | 0.5 (0.01, 1) | medium |
| Lathyrus lanszwertii | Nevada peavine | herbaceous | showy dicots | 0.16 | 0.44 (0, 1) | medium (+/−) |
| Oxalis violacea | violet woodsorrel | herbaceous | showy dicots | 0.02 | 0.45 (0.45, 0.45) | medium (−) |
| Potentilla hippiana | woolly cinquefoil | herbaceous | showy dicots | 0.21 | 0.6 (0, 1) | medium (+/−) |
| Pseudocymopterus montanus | alpine false springparsley | herbaceous | showy dicots | 0.04 | 0.52 (0.09, 0.96) | medium (−) |
| Thermopsis montana var. montana | mountain goldenbanner | herbaceous | showy dicots | 0.18 | 0.41 (0, 1) | medium (+/−) |
| ** Chenopodium graveolens | fetid goosefoot | herbaceous | herbaceous dicots | 0.11 | 0.45 (0.06, 0.93) | medium (+/−) |
| Chenopodium leptophyllum | narrowleaf goosefoot | herbaceous | herbaceous dicots | 0.04 | 0.49 (0.18, 0.81) | medium |
| Iris missouriensis | Rocky Mountain iris | herbaceous | showy monocots | 0.18 | 0.66 (0, 1) | medium (+/−) |
| 1 Arctostaphylos uva-ursi | kinnikinnick | woody | shrubs | 0.18 | 0.45 (0.01, 1) | medium (+/−) |
| Berberis fendleri | Colorado barberry | woody | shrubs | 0.04 | 0.49 (0, 0.97) | medium (+) |
| Brickellia californica | California brickellbush | woody | shrubs | 0.04 | 0.62 (0.26, 0.99) | medium (+) |
| Cercocarpus montanus | mountain mahogany | woody | shrubs | 0.04 | 0.43 (0, 0.85) | medium (+) |
| Jamesia americana | cliffbush | woody | shrubs | 0.04 | 0.51 (0.05, 0.97) | medium (+) |
| Sambucus spp. | elderberry | woody | shrubs | 0.07 | 0.48 (0.05, 1) | medium (+) |
| Pinus ponderosa | ponderosa pine | woody | trees | 0.40 | 0.57 (0, 1) | medium (+/−) |
| Populus tremuloides | quaking aspen | woody | trees | 0.28 | 0.46 (0, 1) | medium (+/−) |
| Pseudotsuga menziesii | Douglas-fir | woody | trees | 0.12 | 0.56 (0, 0.97) | medium (+) |
| Agoseris aurantica | orange agoseris | herbaceous | composites | 0.02 | 0 (0, 0) | low (−) |
| Allium cernuum | nodding onion | herbaceous | showy monocots | 0.14 | 0.3 (0, 1) | low (−) |
| Antennaria marginata | whitemargin pussytoes | herbaceous | composites | 0.02 | 0.18 (0.18, 0.18) | low |
| Artemisia franserioides | ragweed sagebrush | herbaceous | composites | 0.02 | 0.04 (0.04, 0.04) | low (+) |
| Artemisia ludoviciana | white sagebrush | herbaceous | composites | 0.26 | 0.36 (0, 0.93) | low (+/−) |
| Aster spp. | aster | herbaceous | composites | 0.02 | 0.18 (0.18, 0.18) | low |
| Bahia dissecta | ragleaf bahia | herbaceous | composites | 0.25 | 0.39 (0, 1) | low (+/−) |
| Brickelliastrum fendleri | Fendler’s brickellbush | herbaceous | composites | 0.02 | 0 (0, 0) | low |
| Cirsium pallidium | pale thistle | herbaceous | composites | 0.16 | 0.22 (0, 0.81) | low (+) |
| Cirsium spp. | thistle | herbaceous | composites | 0.04 | 0.37 (0.02, 0.72) | low (+) |
| Cirsium undulatum | wavyleaf thistle | herbaceous | composites | 0.02 | 0.04 (0.04, 0.04) | low |
| Conyza canadensis | Canadian horseweed | herbaceous | composites | 0.14 | 0.3 (0.01, 0.99) | low (+/−) |
| Hymenoxys richardsonii | pingue hymenoxys | herbaceous | composites | 0.02 | 0.35 (0.35, 0.35) | low |
| Lactuca serriola | prickly lettuce | herbaceous | composites | 0.04 | 0.1 (0.04, 0.16) | low (−) |
| Packera fendleri | Fendler’s ragwort | herbaceous | composites | 0.05 | 0.32 (0.25, 0.45) | low (−) |
| Pericome caudata | mountain leaftail | herbaceous | composites | 0.02 | 0.05 (0.05, 0.05) | low |
| Pseudognaphalium macounii | Macoun’s cudweed | herbaceous | composites | 0.02 | 0.35 (0.35, 0.35) | low |
| 1 Senecio bigloviii | nodding ragwort | herbaceous | composites | 0.05 | 0.16 (0.08, 0.23) | low (−) |
| 1 Senecio eremophilus | desert groundsel | herbaceous | composites | 0.35 | 0.37 (0, 1) | low (+/−) |
| 1 Senecio wootonii | Wooton’s ragwort | herbaceous | composites | 0.02 | 0.18 (0.18, 0.18) | low |
| Solidago spp. | goldenrod | herbaceous | composites | 0.05 | 0.16 (0, 0.41) | low (−) |
| Solidago mollis | velvety goldenrod | herbaceous | composites | 0.07 | 0.13 (0.01, 0.26) | low |
| Taraxacum officinale | common dandelion | herbaceous | composites | 0.30 | 0.26 (0, 1) | low (−) |
| Tragopogon dubius | yellow salsify | herbaceous | composites | 0.02 | 0 (0, 0) | low |
| Pteridium aquilinum | western brackenfern | herbaceous | ferns | 0.05 | 0.38 (0.04, 0.96) | low (−) |
| Blepharoneuron tricholepis | pine dropseed | herbaceous | grasses | 0.14 | 0.36 (0, 1) | low (−) |
| Bromus anomalus | nodding brome | herbaceous | grasses | 0.02 | 0 (0, 0) | low |
| Bromus carinatus | California brome | herbaceous | grasses | 0.02 | 0 (0, 0) | low (−) |
| Bromus catharticus | rescuegrass | herbaceous | grasses | 0.02 | 0.04 (0.04, 0.04) | low (−) |
| 1 Bromus ciliatus | fringed brome | herbaceous | grasses | 0.16 | 0.31 (0, 0.93) | low (+) |
| Bromus inermis | smooth brome | herbaceous | grasses | 0.19 | 0.13 (0, 0.72) | low (+/−) |
| Bromus tectorum | cheatgrass | herbaceous | grasses | 0.25 | 0.37 (0, 1) | low (+/−) |
| Dactylis glomerata | orchardgrass | herbaceous | grasses | 0.07 | 0.34 (0, 0.81) | low (−) |
| Elymus x pseudorepens | false quackgrass | herbaceous | grasses | 0.07 | 0.26 (0, 0.72) | low (+) |
| Festuca arizonica | Arizona fescue | herbaceous | grasses | 0.11 | 0.26 (0.01, 0.83) | low (−) |
| Festuca brevipila | hard fescue | herbaceous | grasses | 0.09 | 0.27 (0, 0.81) | low (−) |
| Festuca spp. | fescue | herbaceous | grasses | 0.02 | 0 (0, 0) | low |
| Muhlenbergia wrightii | spike muhly | herbaceous | grasses | 0.04 | 0 (0, 0) | low |
| Pascopyrum smithii | western wheatgrass | herbaceous | grasses | 0.18 | 0.14 (0, 0.97) | low (+/−) |
| 1 Piptatherum micranthum | littleseed ricegrass | herbaceous | grasses | 0.02 | 0.01 (0.01, 0.01) | low (−) |
| Poa pratensis | Kentucky bluegrass | herbaceous | grasses | 0.35 | 0.31 (0, 1) | low (+/−) |
| Schizachyrium scoparium | little bluestem | herbaceous | grasses | 0.05 | 0.39 (0, 0.95) | low (−) |
| Carex geophila | White Mountain sedge | herbaceous | rushes and sedges | 0.09 | 0.22 (0, 0.94) | low |
| Carex inops ssp. heliophila | sun sedge | herbaceous | rushes and sedges | 0.23 | 0.33 (0, 0.96) | low (+/−) |
| Carex pellita | woolly sedge | herbaceous | rushes and sedges | 0.02 | 0 (0, 0) | low |
| Carex rossii | Ross’ sedge | herbaceous | rushes and sedges | 0.30 | 0.38 (0, 1) | low (−) |
| Juncus arcticus var. balticus | Baltic rush | herbaceous | rushes and sedges | 0.02 | 0 (0, 0) | low |
| Androsace septentrionalis | pygmyflower rockjasmine | herbaceous | showy dicots | 0.11 | 0.2 (0.01, 0.81) | low (−) |
| Apocynum androsaemifolium | spreading dogbane | herbaceous | showy dicots | 0.04 | 0.21 (0, 0.41) | low (−) |
| Capsella bursa-pastoris | Shepard’s purse | herbaceous | showy dicots | 0.02 | 0.01 (0.01, 0.01) | low |
| Cerastium arvense | mouseear chickweed | herbaceous | showy dicots | 0.19 | 0.37 (0, 1) | low (−) |
| Corydalis aurea | golden smoke | herbaceous | showy dicots | 0.14 | 0.05 (0, 0.18) | low (−) |
| Cryptantha cinerea | James’ catseye | herbaceous | showy dicots | 0.05 | 0.18 (0.02, 0.35) | low (−) |
| Dasiphora fruticosa ssp. floribunda | shrubby cinquefoil | herbaceous | shrubs | 0.02 | 0 (0, 0) | low |
| Descurainia spp. | tansymustard | herbaceous | showy dicots | 0.04 | 0 (0, 0.01) | low (−) |
| Erigeron speciosus | aspen fleabane | herbaceous | composites | 0.05 | 0.03 (0, 0.08) | low (−) |
| 1 Fragaria spp. | strawberry | herbaceous | showy dicots | 0.04 | 0.13 (0.08, 0.18) | low |
| 1 Fragaria virginiana | Virginia strawberry | herbaceous | showy dicots | 0.05 | 0.28 (0, 0.81) | low |
| Geranium caespitosum | pineywoods geranium | herbaceous | showy dicots | 0.07 | 0.03 (0.01, 0.05) | low (−) |
| Geranium richardsonii | Richardson’s geranium | herbaceous | showy dicots | 0.09 | 0.34 (0, 0.96) | low (+) |
| Glandularia bipinnatifida | Dakota mock vervain | herbaceous | showy dicots | 0.02 | 0.05 (0.05, 0.05) | low |
| Hackelia floribunda | manyflowered stickseed | herbaceous | showy dicots | 0.04 | 0.04 (0.02, 0.05) | low |
| Heuchera parvifolia | littleleaf alumroot | herbaceous | showy dicots | 0.02 | 0.05 (0.05, 0.05) | low |
| Lathyrus spp. | peavine | herbaceous | showy dicots | 0.02 | 0.08 (0.08, 0.08) | low |
| Lithospermum multiflorum | manyflowered gromwell | herbaceous | showy dicots | 0.21 | 0.33 (0, 1) | low (+/−) |
| Mentzelia multiflora | manyflowered mentzelia | herbaceous | showy dicots | 0.02 | 0 (0, 0) | low |
| Oenothera elata ssp. hirsutissima | Hooker’s eveningprimrose | herbaceous | showy dicots | 0.04 | 0 (0, 0.01) | low (−) |
| 1 Penstemon barbatus | beardlip penstemon | herbaceous | showy dicots | 0.19 | 0.38 (0, 1) | low (+/−) |
| Thalictrum fendleri | Fendler’s meadowrue | herbaceous | showy dicots | 0.14 | 0.3 (0, 0.96) | low (−) |
| Trifolium repens | white clover | herbaceous | showy dicots | 0.07 | 0.27 (0, 1) | low (+) |
| Verbascum thapsus | common mullein | herbaceous | showy dicots | 0.35 | 0.21 (0, 0.99) | low (+/−) |
| Vicia americana | American vetch | herbaceous | showy dicots | 0.18 | 0.39 (0, 1) | low (+/−) |
| 1 Maianthemum stellatum | starry false Solomon’s seal | herbaceous | showy monocots | 0.05 | 0.29 (0, 0.81) | low |
| Chenopodium fremontii | Fremont’s goosefoot | herbaceous | herbaceous dicots | 0.28 | 0.2 (0, 0.97) | low (+/−) |
| Chenopodium spp. | goosefoot | herbaceous | herbaceous dicots | 0.07 | 0.23 (0, 0.81) | low |
| Ceanothus fendleri | Fendler’s ceanothus | woody | shrubs | 0.16 | 0.39 (0.01, 1) | low (+/−) |
| 1 Juniperus communis | common juniper | woody | shrubs | 0.02 | 0 (0, 0) | low |
| 1 Mahonia repens | creeping barberry | woody | shrubs | 0.02 | 0.01 (0.01, 0.01) | low |
| Physocarpus monogynus | mountain ninebark | woody | shrubs | 0.02 | 0.01 (0.01, 0.01) | low (−) |
| Prunus virginiana | common chokecherry | woody | shrubs | 0.05 | 0.02 (0, 0.05) | low |
| Quercus gambelii | Gambel’s oak | woody | shrubs | 0.49 | 0.36 (0, 1) | low (+/−) |
| Ribes spp. | currant; gooseberry | woody | shrubs | 0.02 | 0.16 (0.16, 0.16) | low (−) |
| Ribes leptanthum | trumpet gooseberry | woody | shrubs | 0.05 | 0.35 (0, 1) | low (+) |
| Robinia neomexicana | New Mexico locust | woody | shrubs | 0.63 | 0.3 (0, 1) | low (+/−) |
| Rosa woodsii | Woods’ rose | woody | shrubs | 0.25 | 0.36 (0, 1) | low (+/−) |
| Rubus idaeus | Red Raspberry | woody | shrubs | 0.11 | 0.33 (0, 1) | low (+/−) |
| 1 Rubus parviflorus | thimbleberry | woody | shrubs | 0.04 | 0.05 (0.04, 0.05) | low |
| Acer glabrum | Rocky Mountain maple | woody | trees | 0.02 | 0.05 (0.05, 0.05) | low |
| Cornus sericea | redosier dogwood | woody | trees | 0.02 | 0.05 (0.05, 0.05) | low |
| 1 Clematis columbiana | rock clematis | woody | vines and trailing plants | 0.12 | 0.15 (0, 0.55) | low (−) |
| Clematis spp. | clematis | woody | vines and trailing plants | 0.07 | 0.23 (0.01, 0.81) | low (−) |
References
- Turner, M.G.; Hargrove, W.W.; Gardner, R.H.; Romme, W.H. Effects of fire on landscape heterogeneity in Yellowstone National Park, Wyoming. J. Veg. Sci. 1994, 5, 731–742. [Google Scholar] [CrossRef]
- Burton, P.J.; Parisien, M.-A.; Hicke, J.A.; Hall, R.J.; Freeburn, J.T. Large fires as agents of ecological diversity in the North American boreal forest. Int. J. Wildl. Fire 2008, 17, 754–767. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.W.J.; Bennett, A.F.; Clarke, M.F. Determinants of the occurrence of unburnt forest patches: Potential biotic refuges within a large, intense wildfire in south-eastern Australia. For. Ecol. Manag. 2014, 314, 85–93. [Google Scholar] [CrossRef]
- Robinson, N.M.; Leonard, S.W.J.; Bennett, A.F.; Clarke, M.F. Refuges for birds in fire-prone landscapes: The influence of fire severity and fire history on the distribution of forest birds. For. Ecol. Manag. 2014, 318, 110–121. [Google Scholar] [CrossRef]
- Reside, A.E.; Welbergen, J.A.; Phillips, B.L.; Wardell-Johnson, G.W.; Keppel, G.; Ferrier, S.; Williams, S.E.; van der Wal, J. Characteristics of climate change refugia for Australian biodiversity. Austral Ecol. 2014, 39, 887–897. [Google Scholar] [CrossRef]
- Keppel, G.; Mokany, K.; Wardell-Johnson, G.W.; Phillips, B.L.; Welbergen, J.A.; Reside, A.E. The capacity of refugia for conservation planning under climate change. Front. Ecol. Environ. 2015, 13, 106–112. [Google Scholar] [CrossRef]
- Keppel, G.; van Niel, K.P.; Wardell-Johnson, G.W.; Yates, C.J.; Byrne, M.; Mucina, L.; Schut, A.G.T.; Hopper, S.D.; Franklin, S.E. Refugia: Identifying and understanding safe havens for biodiversity under climate change. Glob. Ecol. Biogeogr. 2012, 21, 393–404. [Google Scholar] [CrossRef]
- Camp, A.; Oliver, C.; Hessburg, P.; Everett, R. Predicting late-successional fire refugia pre-dating European settlement in the Wenatchee mountains. For. Ecol. Manag. 1997, 95, 63–77. [Google Scholar] [CrossRef]
- Mackey, B.; Lindenmayer, D.B.; Gill, A.M.; McCarthy, M.A.; Lindesay, J. Wildlife, Fire and Future Climate: A Forest Ecosystem Analysis; CSIRO Publishing: Collingwood, Australia, 2002. [Google Scholar]
- Krawchuk, M.A.; Haire, S.L.; Coop, J.D.; Parisien, M.-A.; Whitman, E.; Chong, G.W.; Miller, C. Topographic and fire weather controls of fire refugia in forested ecosystems of northwestern North America. Ecosphere 2016, 7, e01632. [Google Scholar] [CrossRef]
- Wilkin, K.; Ackerly, D.; Stephens, S. Climate change refugia, fire ecology and management. Forests 2016. [Google Scholar] [CrossRef]
- Ouarmim, S.; Asselin, H.; Hély, C.; Bergeron, Y.; Ali, A.A. Long-term dynamics of fire refuges in boreal mixedwood forests. J. Quat. Sci. 2014, 29, 123–129. [Google Scholar] [CrossRef]
- Madoui, A.; Leduc, A.; Gauthier, S.; Bergeron, Y. Spatial pattern analyses of post-fire residual stands in the black spruce boreal forest of western Quebec. Int. J. Wildl. Fire 2010, 19, 1110. [Google Scholar] [CrossRef]
- Collins, L.; Bradstock, R.A.; Tasker, E.M.; Whelan, R.J. Can gullies preserve complex forest structure in frequently burnt landscapes? Biol. Conserv. 2012, 153, 177–186. [Google Scholar] [CrossRef]
- Román-Cuesta, R.M.; Gracia, M.; Retana, J. Factors influencing the formation of unburned forest islands within the perimeter of a large forest fire. For. Ecol. Manag. 2009, 258, 71–80. [Google Scholar] [CrossRef]
- Buma, B. Disturbance interactions: Characterization, prediction, and the potential for cascading effects. Ecosphere 2015, 6, art70. [Google Scholar] [CrossRef]
- Coop, J.D.; Parks, S.A.; McClernan, S.R.; Holsinger, L.M. Influences of prior wildfires on vegetation response to subsequent fire in a reburned southwestern landscape. Ecol. Appl. 2016, 26, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-wildfire logging hinders regeneration and increases fire risk. Science 2006, 311, 352. [Google Scholar] [CrossRef] [PubMed]
- Haire, S.L.; McGarigal, K. Inhabitants of landscape scars: Succession of woody plants after large, severe forest fires in Arizona and New Mexico. Southwest. Nat. 2008, 53, 146–161. [Google Scholar] [CrossRef]
- Serra-Diaz, J.M.; Scheller, R.M.; Syphard, A.D.; Franklin, J. Disturbance and climate microrefugia mediate tree range shifts during climate change. Landsc. Ecol. 2015, 30, 1039–1053. [Google Scholar] [CrossRef]
- Ouarmim, S.; Asselin, H.; Bergeron, Y.; Ali, A.A.; Hély, C. Stand structure in fire refuges of the eastern Canadian boreal mixedwood forest. For. Ecol. Manag. 2014, 324, 1–7. [Google Scholar] [CrossRef]
- Anand, M.; Leithead, M.; Silva, L.C.R.; Wagner, C.; Ashiq, M.W.; Cecile, J.; Drobyshev, I.; Bergeron, Y.; Das, A.; Bulger, C. The scientific value of the largest remaining old-growth red pine forests in North America. Biodivers. Conserv. 2013, 22, 1847–1861. [Google Scholar] [CrossRef]
- Bond, W.J.; Dickinson, K.J.M.; Mark, A.F. What limits the spread of fire-dependent vegetation? Evidence from geographic variation of serotiny in a New Zealand shrub. Glob. Ecol. Biogeogr. 2004, 13, 115–127. [Google Scholar] [CrossRef]
- Mackey, B.; Berry, S.; Hugh, S.; Ferrier, S.; Harwood, T.D.; Williams, K.J. Ecosystem greenspots: Identifying potential drought, fire, and climate-change micro-refuges. Ecol. Appl. 2012, 22, 1852–1864. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J. Habitat islands in fire-prone vegetation: Do landscape features influence community composition? J. Biogeogr. 2002, 29, 677–684. [Google Scholar] [CrossRef]
- Gould, S.F.; Hugh, S.; Porfirio, L.L.; Mackey, B. Ecosystem greenspots pass the first test. Landsc. Ecol. 2015, 30, 141–151. [Google Scholar] [CrossRef]
- Wood, S.W.; Murphy, B.P.; Bowman, D.M.J.S. Firescape ecology: How topography determines the contrasting distribution of fire and rain forest in the south-west of the Tasmanian Wilderness World Heritage Area. J. Biogeogr. 2011, 38, 1807–1820. [Google Scholar] [CrossRef]
- Kolden, C.A.; Abatzoglou, J.T.; Lutz, J.A.; Cansler, C.A.; Kane, J.T.; van Wagtendonk, J.W.; Key, C.H. Climate contributors to forest mosaics: Ecological persistence following wildfire. Northwest Sci. 2015, 89, 219–238. [Google Scholar] [CrossRef]
- Kolden, C.A.; Lutz, J.A.; Key, C.H. Mapped versus actual burned area within wildfire perimeters: Characterizing the unburned. For. Ecol. 2012, 286, 38–47. [Google Scholar] [CrossRef]
- Nelson, T.A.; Boots, B. Detecting spatial hot spots in landscape ecology. Ecography (Cop.) 2008, 31, 556–566. [Google Scholar] [CrossRef]
- Silverman, B.W. Density Estimation for Statistics and Data Analysis; Chapman and Hall: New York, NY, USA, 1986. [Google Scholar]
- Azzalini, A.; Torelli, N. Clustering via nonparametric density estimation. Stat. Comput. 2007, 17, 71–80. [Google Scholar] [CrossRef]
- Longland, W.S.; Bateman, S.L. Viewpoint: The ecological value of shrub islands on disturbed sagebrush rangelands. J. Range Manag. 2014, 55, 571–575. [Google Scholar] [CrossRef]
- Liebmann, M.J.; Farella, J.; Roos, C.I.; Stack, A.; Martini, S.; Swetnam, T.W. Native American depopulation, reforestation, and fire regimes in the Southwest United States, 1492–1900 CE. Proc. Natl. Acad. Sci. USA 2016, 113, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D. Lots of Lightning and Plenty of People: An Ecological History of Fire in the Upland Southwest. In Fire, Native Peoples, and the Natural Landscape; Vale, T.R., Ed.; Island Press: Washington, DC, USA, 2002; pp. 143–193. [Google Scholar]
- Dumire, W.W.; Tierney, G.D. Wild plants of the Pueblo Province: Exploring Ancient and Enduring Uses; Museum of New Mexico Press: Santa Fe, NM, USA, 1995. [Google Scholar]
- Swetnam, T.W.; Farella, J.; Roos, C.I.; Liebmann, M.J.; Falk, D.A.; Allen, C.D. Multiscale perspectives of fire, climate and humans in western North America and the Jemez Mountains, USA. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Muldavin, E.; Kennedy, A.; Jackson, C.; Neville, P.; Neville, T.; Schultz, K.; Reid, M. Vegetation Classification and Map: Bandelier National Monument Natural Resource 361 Technical Report NPS/SCPN/NRTR--2011/438; National Park Service: Fort Collins, CO, USA, 2011. [Google Scholar]
- Commission for Environmental Cooperation. North American Terrestrial Ecosystems—Level III; Commission for Environmental Cooperation: Montréal, QC, Canada, 2011. [Google Scholar]
- Swetnam, T.W.; Betancourt, J.L.; Swetnam, T.W.; Betancourt, J.L. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. J. Clim. 1998, 11, 3128–3147. [Google Scholar] [CrossRef]
- Wolter, K. Multivariate ENSO Index. Available online: http://www.esrl.noaa.gov/psd/enso/mei/ (accessed on 14 November 2016).
- Schoeffler, F.J.; Wachter, J.B. Dynamic Fire Weather Associated with the June 2011 New Mexico (USA) Las Conchas Fire (95th American Meteorological Society Annual Meeting). In Proceedings of the 95th American Meterorological Society, Phoenix, AZ, USA, 4–8 January 2015.
- Reneau, S.L.; Mcdonald, E.V. Landscape History and Processes on the Pajarito Plateau, Northern New Mexico; Los Alamos National Laboratory: Los Alamos, NM, USA, 1996. [Google Scholar]
- Schoennagel, T.; Veblen, T.T.; Romme, W.H. The interaction of fire, fuels, and climate across Rocky Mountain forests. Bioscience 2004, 54, 661–676. [Google Scholar] [CrossRef]
- Romme, W.H.; Allen, C.D.; Bailey, J.D.; Baker, W.L.; Bestelmeyer, B.T.; Brown, P.M.; Eisenhart, K.S.; Floyd, M.L.; Huffman, D.W.; Jacobs, B.F.; et al. Historical and modern disturbance regimes, stand structures, and landscape dynamics in piñon–juniper vegetation of the Western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef]
- Haire, S.L.; McGarigal, K. Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landsc. Ecol. 2010, 25, 1055–1069. [Google Scholar] [CrossRef]
- Eidenshink, J.; Schwind, B.; Brewer, K.; Zhu, Z.; Quayle, B.; Howard, S.; Falls, S.; Falls, S. A project for monitoring trends in burn severity. Fire Ecol. 2007, 3, 3–21. [Google Scholar] [CrossRef]
- GEOMAC. Geosciences and Environmental Change Science Center (GECSC) Outgoing Datasets. Available online: http://rmgsc.cr.usgs.gov/outgoing/GeoMAC/2011_fire_data/New_Mexico/Las_Conchas/ (accessed on 14 November 2016).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 2016, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 2.5–8. Available online: https://cran.r-project.org/package=raster (accessed on 14 November 2016).
- Brenning, A. Statistical Geocomputing Combining R and SAGA: The example of landslide susceptibility analysis with Generalized Additive Models. In SAGA--Seconds Out (Hamburger Beiträge zur Physischen Geographie und Landschaftsökologie; Böhner, J., Blaschke, T., Montanarella, L., Eds.; Saga Press: Portland, OR, USA, 2008; Volume 19, pp. 23–32. [Google Scholar]
- Brenning, A.; Bangs, D. RSAGA: SAGA Geoprocessing and Terrain Analysis in R. Available online: https://CRAN.R-project.org/package=RSAGA (accessed on 14 November 2016).
- Wood, S.N. mgcv: Mixed GAM Computation Vehicle with GCV/AIC/REML Smoothness Estimation. R Package Version 1.8–15. Available online: https://cran.r-project.org/package=mgcv (accessed on 14 November 2016).
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Hurvich, C.M.; Tsai, C.-L. Model selection for least absolute deviations regression in small samples. Stat. Probab. Lett. 1990, 9, 259–265. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/package=vegan (accessed on 14 November 2016).
- Foxx, T.S.; Hoard, D. Flowering Plants of the Southwestern Woodlands; Otowi Crossing Press: Los Alamos, NM, USA, 1995. [Google Scholar]
- Turner, M.G.; Romme, W.H.; Gardner, R.H.; Hargrove, W.W. Effects of fire size and pattern on early succcession in Yellowstone National Park. Ecol. Monogr. 1997, 67, 411–433. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Tinker, D.B. Surprises and lessons from the 1988 Yellowstone fires. Front. Ecol. Environ. 2003, 1, 351–358. [Google Scholar] [CrossRef]
- Coop, J.D.; Givnish, T.J. Gradient analysis of reversed treelines and grasslands of the Valles Caldera, New Mexico. J. Veg. Sci. 2007, 18, 43–54. [Google Scholar] [CrossRef]
- Holden, Z.A.; Morgan, P.; Evans, J.S. A predictive model of burn severity based on 20-year satellite-inferred burn severity data in a large southwestern US wilderness area. For. Ecol. Manag. 2009, 258, 2399–2406. [Google Scholar] [CrossRef]
- Kane, V.R.; Lutz, J.A.; Cansler, C.A.; Povak, N.A.; Churchill, D.J.; Smith, D.F.; Kane, J.T.; North, M.P. Water balance and topography predict fire and forest structure patterns. For. Ecol. Manag. 2015, 338, 1–13. [Google Scholar] [CrossRef]
- Dobrowski, S.Z. A climatic basis for microrefugia: The influence of terrain on climate. Glob. Chang. Biol. 2010, 17, 1022–1035. [Google Scholar] [CrossRef]
- Hume, A.; Chen, H.Y.H.; Taylor, A.R.; Kayahara, G.J.; Man, R. Soil C:N:P dynamics during secondary succession following fire in the boreal forest of central Canada. For. Ecol. Manag. 2016, 369, 1–9. [Google Scholar] [CrossRef]
- Herrmann, J.D.; Carlo, T.A.; Brudvig, L.A.; Damschen, E.I.; Haddad, N.M.; Levey, D.J.; Orrock, J.L.; Tewksbury, J.J. Connectivity from a different perspective: Comparing seed dispersal kernels in connected vs. unfragmented landscapes. Ecology 2016, 97, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Neary, D.G.; Ryan, K.C.; de Bano, L.F. Wildland Fire in Ecosystems: Effects of Fire on Soils and Water; Gen. Tech. Rep. RMRS-GTR-42-vol.4; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2005.
- Weber, C.F.; Lockhart, J.S.; Charaska, E.; Aho, K.; Lohse, K.A. Bacterial composition of soils in ponderosa pine and mixed conifer forests exposed to different wildfire burn severity. Soil Biol. Biochem. 2014, 69, 242–250. [Google Scholar] [CrossRef]
- Kotliar, N.B.; Kennedy, P.L.; Ferree, K. Avifaunal responses to fire in southwestern montane forests along a burn severity gradient. Ecol. Appl. 2007, 17, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, T.J.; Vierling, K.T.; Kozma, J.M.; Millard, J.E.; Raphael, M.G. Space use by white-headed woodpeckers and selection for recent forest disturbances. J. Wildl. Manag. 2015, 79, 1286–1297. [Google Scholar] [CrossRef]
- Chia, E.K.; Bassett, M.; Nimmo, D.G.; Leonard, S.W.J.; Ritchie, E.G.; Clarke, M.F.; Bennett, A.F. Fire severity and fire-induced landscape heterogeneity affect arboreal mammals in fire-prone forests. Ecosphere 2015, 6, art190. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N.; Feddema, J.J. Double whammy: High-severity fire and drought in ponderosa pine forests of the Southwest. Can. J. For. Res. 2013, 43, 570–583. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N. How resilient are southwestern ponderosa pine forests after crown fires? Can. J. For. Res. 2005, 35, 967–977. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; Della Sala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar]
- Roberts, S.L.; Kelt, D.A.; van Wagtendonk, J.W.; Miles, A.K.; Meyer, M.D. Effects of fire on small mammal communities in frequent-fire forests in California. J. Mammal. 2015, 96, 107–119. [Google Scholar] [CrossRef]
- Soulard, C.E.; Albano, C.M.; Villarreal, M.L.; Walker, J.J. Continuous 1985–2012 Landsat monitoring to assess fire effects on meadows in Yosemite National Park, California. Remote Sens. 2016, 8, 371. [Google Scholar] [CrossRef]
- Debinski, D.M.; Jakubauskas, M.E.; Kindscher, K. Montane meadows as indicators of environmental change. Environ. Monit. Assess. 2000, 64, 213–225. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Keeley, J.E. A plant distribution shift: Temperature, drought or past disturbance? PLoS ONE 2012, 7, e31173. [Google Scholar] [CrossRef] [PubMed]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.P.; Freeborn, P.; Rieck, J.D.; Calkin, D.E.; Gilbertson-Day, J.W.; Cochrane, M.A.; Hand, M.S. Quantifying the influence of previously burned areas on suppression effectiveness and avoided exposure: A case study of the Las Conchas Fire. Int. J. Wildl. Fire 2016, 25, 167. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. Biol. Sci. 2010, 277, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Breshears, D.D. Drought-induced shift of a forest woodland ecotone: Rapid landscape response to climate variation. Proc. Natl. Acad. Sci. USA 1998, 95, 14839–14842. [Google Scholar] [CrossRef] [PubMed]
- Hampe, A.; Jump, A.S. Climate relicts: Past, present, and future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.B.; Macgregor, C.; Wood, J.T.; Cunningham, R.B.; Crane, M.; Michael, D.; Montague-Drake, R.; Brown, D.; Fortescue, M.; Dexter, N.; et al. What factors influence rapid post-fire site re-occupancy? A case study of the endangered Eastern Bristlebird in eastern Australia. Int. J. Wildl. Fire 2009, 18, 84–95. [Google Scholar] [CrossRef]
- Berry, L.E.; Driscoll, D.A.; Stein, J.A.; Blanchard, W.; Banks, S.C.; Bradstock, R.A.; Lindenmayer, D.B. Identifying the location of fire refuges in wet forest ecosystems. Ecol. Appl. 2015, 25, 2337–2348. [Google Scholar] [CrossRef] [PubMed]
- Stephens, S.L.; Agee, J.K.; Fule, P.Z.; North, M.P.; Romme, W.H.; Swetnam, T.W.; Turner, M.G. Managing forests and fire in changing climates. Science 2013, 342, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Montaño, C.; Fulé, P.Z.; Falk, D.A.; Villanueva-Díaz, J.; Yocom, L.L. Linking old-growth forest composition, structure, fire history, climate and land-use in the mountains of northern México. Ecosphere 2012, 3, art106. [Google Scholar] [CrossRef]
- Margolis, E.Q.; Malevich, S.B. Historical dominance of low-severity fire in dry and wet mixed-conifer forest habitats of the endangered terrestrial Jemez Mountains salamander (Plethodon neomexicanus). For. Ecol. Manag. 2016, 375, 12–26. [Google Scholar] [CrossRef]
- Shinneman, D.J.; Means, R.E.; Potter, K.M.; Hipkins, V.D. Exploring climate niches of ponderosa pine (Pinus ponderosa Douglas ex Lawson) haplotypes in the western United States: Implications for evolutionary history and conservation. PLoS ONE 2016, 11, e0151811. [Google Scholar] [CrossRef] [PubMed]





| Terrain Metric | Description |
|---|---|
| Local morphometry | Method: 2nd degree polynomial |
| Slope | In radians |
| Aspect | In radians |
| Overall curvature | Positive values: convex |
| Profile curvature | Positive values: convex |
| Plan curvature | Positive values: divergent |
| Characteristics of the Catchment Basin | Method: multiple flow direction |
| Slope | Average slope of the contributing area in radians |
| Size | Size of the contributing area in m2 |
| Height | Altitudinal difference (relief) |
| Length | Length of flow path (m) |
| Aspect | Direction of slope (radians) |
| Other Indices | |
| Topographic Convergence Index | Cold air pooling: log(size of the contributing area/local slope) |
| Topographic Wetness Index | Potential soil wetness based on area and slope of the contributing area |
| Relative position | Position of a raster cell compared to its surrounding cells (500-m radius window) |
| Topographic Wind Shelter | Maximum gradient in topographic relief from eight upwind directions (500-m radius window) |
| Climate Variable (Bioclim) | |
| MAT | Mean Annual Temperature (°C) |
| MAP | Mean Annual Precipitation (mm) |
| NFFD | The Number Of Frost-Free Days |
| Fire Name (Start Date) | Fire Size (ha) | Data Source (Resolution) | Pre- and Post-Fire Image Dates | Processing Algorithm | Field Plots (n) |
|---|---|---|---|---|---|
| La Mesa (16/06/1977) | 5777 | Multispectral Scanner Path/Row 35/36 (60-m) | 29/05/1977; 18/05/1978 | 15 | |
| Dome (25/04/1996) | 6387 | Thematic Mapper Path/Row 33/35 (30-m) | 23/07/1995; 14/09/1997 | 9 | |
| Cerro Grande (05/05/2000) | 17919 | Thematic Mapper Path/Row 33/35 (30-m) | 14/04/2000; 25/04/2001 | 28 | |
| Las Conchas (26/06/2011) | 61057 | Thematic Mapper Path/Row 33/35 (30-m) | 02/09/2010; 05/09/2011 | 7 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haire, S.L.; Coop, J.D.; Miller, C. Characterizing Spatial Neighborhoods of Refugia Following Large Fires in Northern New Mexico USA. Land 2017, 6, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/land6010019
Haire SL, Coop JD, Miller C. Characterizing Spatial Neighborhoods of Refugia Following Large Fires in Northern New Mexico USA. Land. 2017; 6(1):19. https://0-doi-org.brum.beds.ac.uk/10.3390/land6010019
Chicago/Turabian StyleHaire, Sandra L., Jonathan D. Coop, and Carol Miller. 2017. "Characterizing Spatial Neighborhoods of Refugia Following Large Fires in Northern New Mexico USA" Land 6, no. 1: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/land6010019