Estimation of the Spatiotemporal Patterns of Vegetation and Associated Ecosystem Services in a Bornean Montane Zone Using Three Shifting-Cultivation Scenarios

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Field Survey
2.2. Field Data Analysis
3. Remote-Sensing Analysis
3.1. Extrapolation Procedure of Above-Ground Biomass (AGB) and Community Composition
3.2. Simulations Using Land-Use Scenarios
4. Results
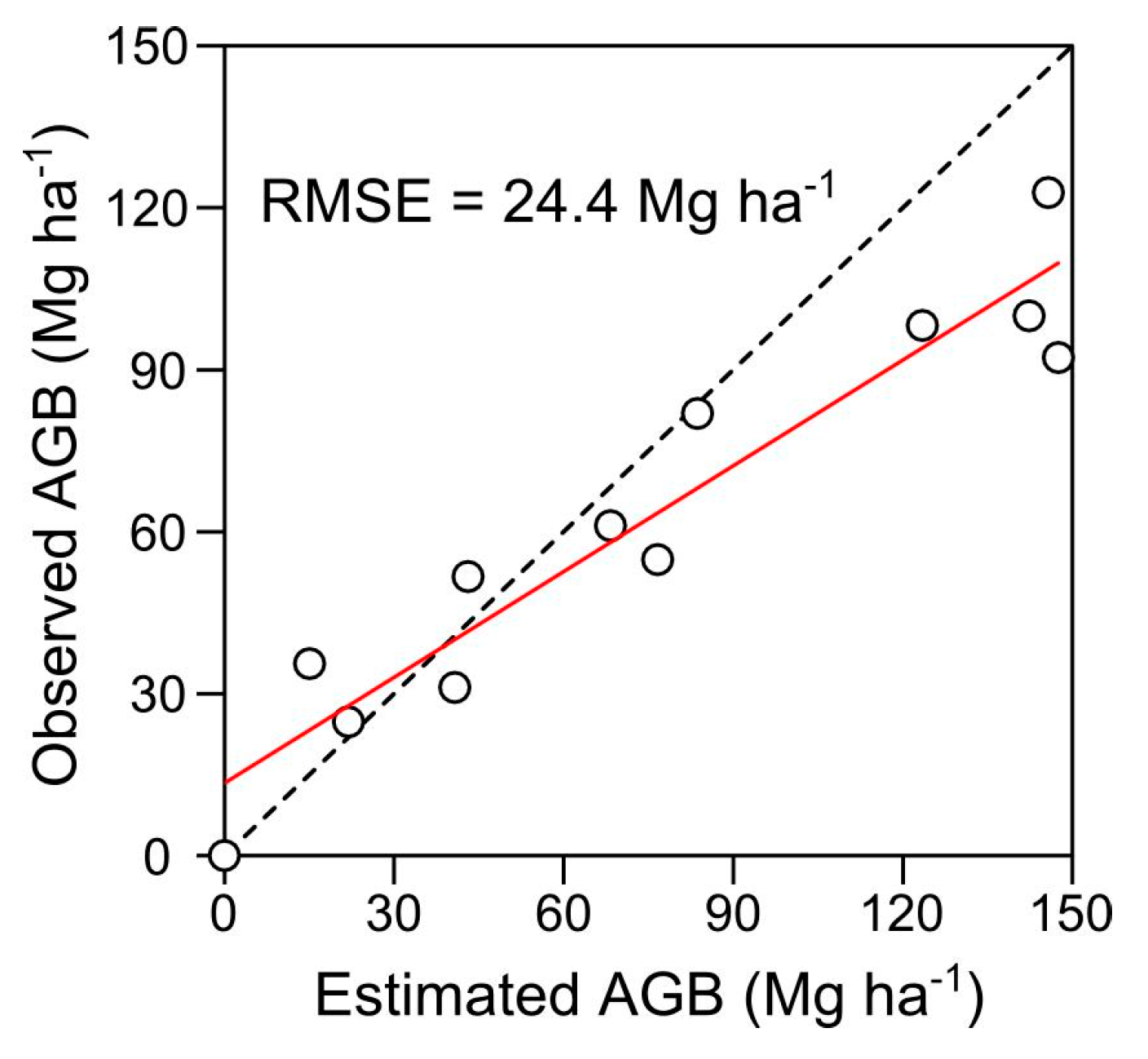
4.1. AGB Map and Plant-Community Composition Map
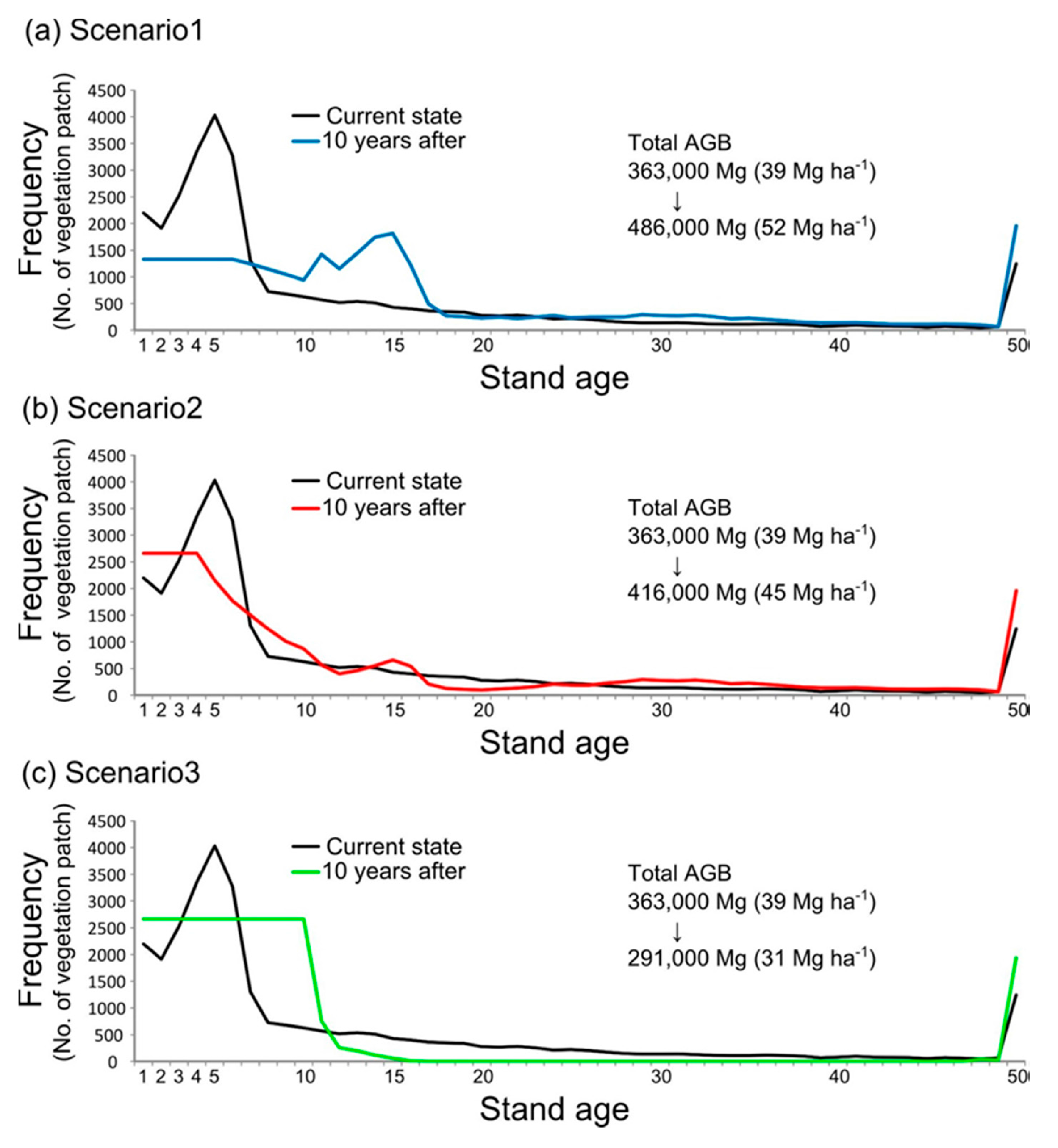
4.2. Numerical Simulation of Vegetation Patterns Based on Three Scenarios
5. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Metzger, J.P. Effects of slash-and-burn fallow periods on landscape structure. Environ. Conserv. 2003, 30, 325–333. [Google Scholar] [CrossRef]
- Mertz, O.; Padoch, C.; Fox, J.; Cramb, R.A.; Leisz, S.J.; Lam, N.T.; Vien, T.D. Swidden change in Southeast Asia: Understanding causes and consequences. Hum. Ecol. 2009, 37, 259–264. [Google Scholar] [CrossRef]
- Van Vliet, N.; Mertz, O.; Heinimann, A.; Langanke, T.; Pascual, U.; Schmook, B.; Adams, C.; Schmidt-Vogt, D.; Messerli, P.; Leisz, S.; et al. Trends, drivers and impacts of changes in Swidden cultivation in tropical forest-agriculture frontiers: A global assessment. Glob. Environ. Chang. 2012, 22, 418–429. [Google Scholar] [CrossRef]
- Dressler, W.; Wilson, D.; Clendenning, J.; Cramb, R.; Mahanty, S.; Lasco, R.; Keenan, R.; To, P.; Gevana, D. Examining how long fallow Swidden systems impact upon livelihood and ecosystem services outcomes compared with alternative land-uses in the uplands of Southeast Asia. J. Dev. Eff. 2015, 210–229. [Google Scholar] [CrossRef]
- Dalle, S.P.; Pulido, M.T.; Blois, S. De Balancing shifting cultivation and forest conservation: Lessons from a “sustainable landscape” in southeastern Mexico. Ecol. Appl. 2011, 21, 1557–1572. [Google Scholar] [CrossRef] [PubMed]
- Geist, H.J.; Lambin, E.F. Proximate Causes and Underlying Driving Forces of Tropical Deforestation Tropical forests are disappearing as the result of many pressures, both local and regional, acting in various combinations in different geographical locations. Bioscience 2002, 52, 143–150. [Google Scholar] [CrossRef]
- Davidson, E.A.; de Abreu Sá, T.D.; Reis Carvalho, C.J.; de Oliveira Figueiredo, R.; Kato, M.S.A.; Kato, O.R.; Ishida, F.Y. An integrated greenhouse gas assessment of an alternative to slash-and-burn agriculture in eastern Amazonia. Glob. Chang. Biol. 2008, 14, 998–1007. [Google Scholar] [CrossRef]
- Novacek, M.J.; Cleland, E.E. The current biodiversity extinction event: Scenarios for mitigation and recovery. Proc. Natl. Acad. Sci. USA 2001, 98, 5466–5470. [Google Scholar] [CrossRef] [PubMed]
- Brook, B.W.; Sodhi, N.S.; Ng, P.K.L. Catastrophic extinctions follow deforestation in Singapore. Nature 2003, 424, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Pandit, M.K.; Sodhi, N.S.; Koh, L.P.; Bhaskar, A.; Brook, B.W. Unreported yet massive deforestation driving loss of endemic biodiversity in Indian Himalaya. Biodivers. Conserv. 2007, 16, 153–163. [Google Scholar] [CrossRef]
- Allnutt, T.F.; Ferrier, S.; Manion, G.; Powell, G.V.N.; Ricketts, T.H.; Fisher, B.L.; Harper, G.J.; Irwin, M.E.; Kremen, C.; Labat, J.-N.; et al. A method for quantifying biodiversity loss and its application to a 50-year record of deforestation across Madagascar. Conserv. Lett. 2008, 1, 173–181. [Google Scholar] [CrossRef]
- Fujiki, S.; Okada, K.; Nishio, S.; Kitayama, K. Estimation of the stand ages of tropical secondary forests after shifting cultivation based on the combination of WorldView-2 and time-series Landsat images. ISPRS J. Photogramm. Remote Sens. 2016, 119, 280–293. [Google Scholar] [CrossRef]
- Riswan, S.; Hartanti, L. Human impacts on tropical forest dynamics. Vegetatio 1995, 121, 41–52. [Google Scholar] [CrossRef]
- Langner, A.; Miettinen, J.; Siegert, F. Land cover change 2002–2005 in Borneo and the role of fire derived from MODIS imagery. Glob. Chang. Biol. 2007, 13, 2329–2340. [Google Scholar] [CrossRef]
- Padoch, C.; Pinedo-Vasquez, M. Saving slash-and-burn to save biodiversity. Biotropica 2010, 42, 550–552. [Google Scholar] [CrossRef]
- Bonner, M.T.L.; Schmidt, S.; Shoo, L.P. A meta-analytical global comparison of aboveground biomass accumulation between tropical secondary forests and monoculture plantations. For. Ecol. Manag. 2013, 291, 73–86. [Google Scholar] [CrossRef]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P.; Gilroy, J.J.; Woodcock, P.; Edwards, F.A.; Larsen, T.H.; Andrews, D.J.R.; Derhé, M.A.; Docherty, T.D.S.; Hsu, W.W.; Mitchell, S.L.; et al. Land-sharing versus land-sparing logging: Reconciling timber extraction with biodiversity conservation. Glob. Chang. Biol. 2014, 20, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Vogt, D.; Leisz, S.J.; Mertz, O.; Heinimann, A.; Thiha, T.; Messerli, P.; Epprecht, M.; Van Cu, P.; Chi, V.K.; Hardiono, M.; et al. An assessment of trends in the extent of Swidden in Southeast Asia. Hum. Ecol. 2009, 37, 269. [Google Scholar] [CrossRef]
- DeFries, R.; Rosenzweig, C. Toward a whole-landscape approach for sustainable land use in the tropics. Proc. Natl. Acad. Sci. USA 2010, 107, 19627–19632. [Google Scholar] [CrossRef] [PubMed]
- Pettorelli, N.; Wegmann, M.; Skidmore, A.; Mücher, S.; Dawson, T.P.; Fernandez, M.; Lucas, R.; Schaepman, M.E.; Wang, T.; O’Connor, B.; et al. Framing the concept of satellite remote sensing essential biodiversity variables: Challenges and future directions. Remote Sens. Ecol. Conserv. 2016, 2, 122–131. [Google Scholar] [CrossRef]
- Fujiki, S.; Nishio, S.; Okada, K.; Nais, J.; Kitayama, K. Plant communities and ecosystem processes in a succession-altitude matrix after shifting cultivation in the tropical montane forest zone of northern Borneo. J. Trop. Ecol. 2017, 33, 33–49. [Google Scholar] [CrossRef]
- Beaman, J.H. Mount Kinabalu: Hotspot of plant diversity in Borneo. Biol. Skr. 2005, 55, 103–127. [Google Scholar]
- ERE Consulting Group Sdn Bhd. Study on the Establishment of Ecological Linkages Connecting the Kinabalu Park and Crocker Range Park Final Report; ERE Consulting Group Sdh Bhd: Subang Jaya, Malaysia, 2011. [Google Scholar]
- Kitayama, K. An altitudinal transect study of the vegetation on Mount Kinabalu, Borneo. Vegetatio 1992, 102, 149–171. [Google Scholar] [CrossRef]
- Aiba, S.; Kitayama, K. Structure, composition and species diversity in an altitude-substrate matrix of rain forest tree communities on Mount Kinabalu, Borneo. Plant Ecol. 1999, 140, 139–157. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Hattori, D.; Itioka, T.; Handa, C.; Ohkubo, T.; Kendawang, J.J.; Nakamura, M.; Sakaguchi, M.; Takahashi, N.; et al. Development of allometric relationships for accurate estimation of above- and below-ground biomass in tropical secondary forests in Sarawak, Malaysia. J. Trop. Ecol. 2009, 25, 371–386. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Vermote, E.F.; Tanre, D.; Deuze, J.L.; Herman, M.; Morcette, J.-J. Second simulation of the satellite signal in the solar spectrum, 6S: An overview. IEEE Trans. Geosci. Remote Sens. 1997, 35, 675–686. [Google Scholar] [CrossRef]
- Ekstrand, S. No TLandsat TM-based forest damage assessment: Correction for topographic effects. Photogramm. Eng. Remote Sens. 1996, 62, 151–162. [Google Scholar]
- Uhl, C.; Clark, K.; Clark, H.; Murphy, P. Early plant succession after cutting and burning in the upper Rio Negro region of the Amazon Basin. J. Ecol. 1981, 69, 631–649. [Google Scholar] [CrossRef]
- Lawrence, D. Erosion of tree diversity during 200 years of shifting cultivation in Bornean rain forest. Ecol. Appl. 2004, 14, 1855–1869. [Google Scholar] [CrossRef]
- Lawrence, D.; Radel, C.; Tully, K.; Schmook, B.; Schneider, L. Untangling a decline in tropical forest resilience: Constraints on the sustainability of shifting cultivation across the globe. Biotropica 2010, 42, 21–30. [Google Scholar] [CrossRef]
- Jakovac, C.C.; Peña-Claros, M.; Kuyper, T.W.; Bongers, F. Loss of secondary-forest resilience by land-use intensification in the Amazon. J. Ecol. 2015, 103, 67–77. [Google Scholar] [CrossRef]
- Lawrence, D.; D’Odorico, P.; Diekmann, L.; DeLonge, M.; Das, R.; Eaton, J. Ecological feedbacks following deforestation create the potential for a catastrophic ecosystem shift in tropical dry forest. Proc. Natl. Acad. Sci. USA 2007, 104, 20696–20701. [Google Scholar] [CrossRef] [PubMed]
- Runyan, C.W.; D’Odorico, P.; Lawrence, D. Effect of repeated deforestation on vegetation dynamics for phosphorus-limited tropical forests. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiki, S.; Nishio, S.; Okada, K.-i.; Nais, J.; Repin, R.; Kitayama, K. Estimation of the Spatiotemporal Patterns of Vegetation and Associated Ecosystem Services in a Bornean Montane Zone Using Three Shifting-Cultivation Scenarios. Land 2018, 7, 29. https://0-doi-org.brum.beds.ac.uk/10.3390/land7010029
Fujiki S, Nishio S, Okada K-i, Nais J, Repin R, Kitayama K. Estimation of the Spatiotemporal Patterns of Vegetation and Associated Ecosystem Services in a Bornean Montane Zone Using Three Shifting-Cultivation Scenarios. Land. 2018; 7(1):29. https://0-doi-org.brum.beds.ac.uk/10.3390/land7010029
Chicago/Turabian StyleFujiki, Shogoro, Shogo Nishio, Kei-ichi Okada, Jamili Nais, Rimi Repin, and Kanehiro Kitayama. 2018. "Estimation of the Spatiotemporal Patterns of Vegetation and Associated Ecosystem Services in a Bornean Montane Zone Using Three Shifting-Cultivation Scenarios" Land 7, no. 1: 29. https://0-doi-org.brum.beds.ac.uk/10.3390/land7010029