
Cellular, Antibody and Cytokine Pathways in Children with Acute SARS-CoV-2 Infection and MIS-C—Can We Match the Puzzle?

 ,
, 
 ,
, 
Abstract
:1. Introduction
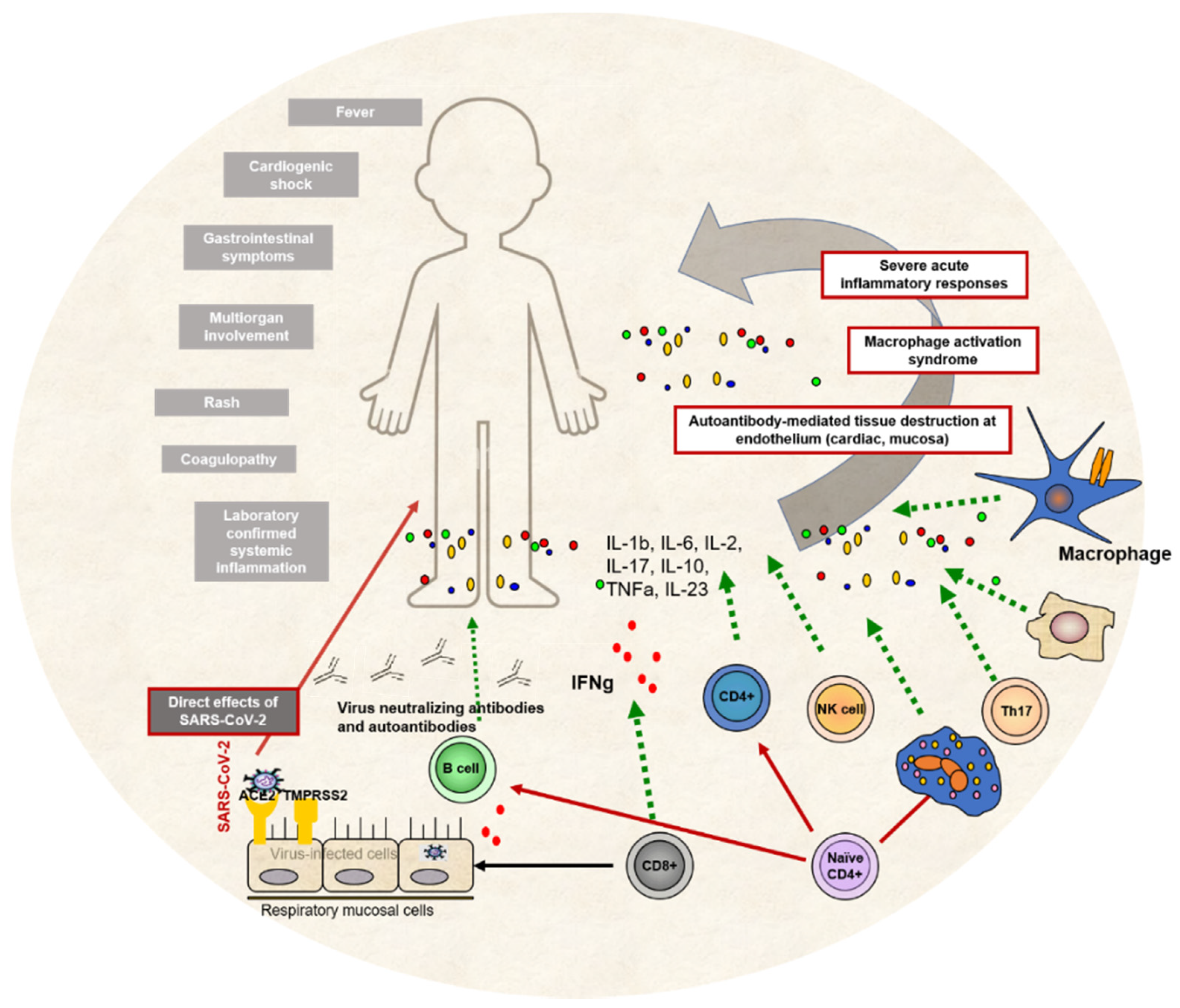
2. Immunology of MIS-C
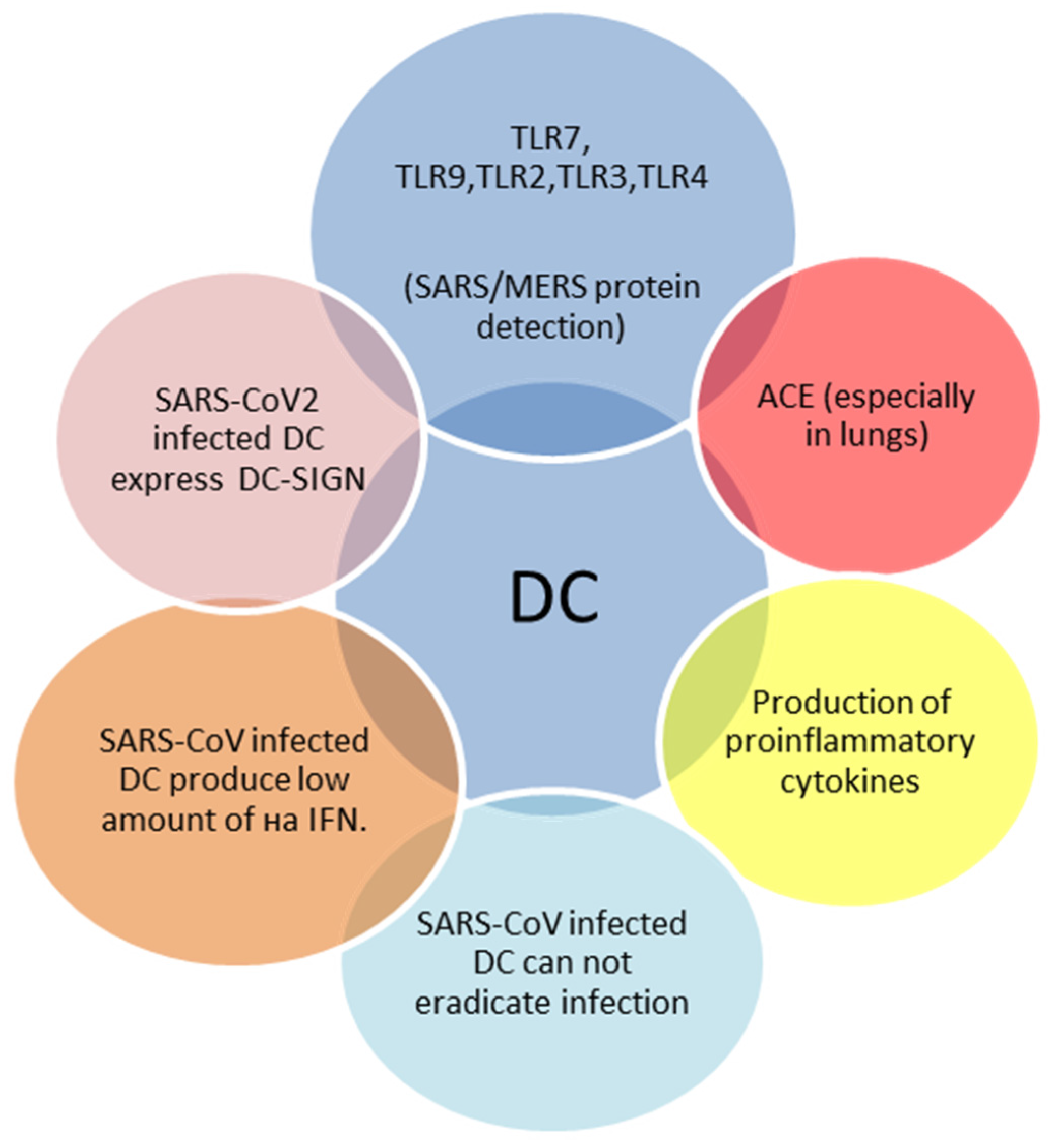
2.1. Innate Immune Response to SARS-CoV-2
2.2. Adaptive Immune Responses to SARS-CoV-2
3. Immune Response Profile and Long COVID-19 in Children
4. Future Directions of Immunological Studies of MIS-C
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Novel Coronavirus Situation Dashboard. Available online: https://covid19.who.int/ (accessed on 15 January 2022).
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. China Novel Coronavirus Investigating and Research Team. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Rothe, C.; Schunk, M.; Sothmann, P.; Bretzel, G.; Froeschl, G.; Wallrauch, C.; Zimmer, T.; Thiel, V.; Janke, C.; Guggemos, W.; et al. Transmission of 2019-nCoV Infection from an Asymptomatic Contact in Germany. N. Engl. J. Med. 2020, 382, 970–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graciano-Machuca, O.; Villegas-Rivera, G.; López-Pérez, I.; Macías-Barragán, J.; Sifuentes-Franco, S. Multisystem Inflammatory Syndrome in Children (MIS-C) Following SARS-CoV-2 Infection: Role of Oxidative Stress. Front. Immunol. 2021, 12, 723654. [Google Scholar] [CrossRef] [PubMed]
- WHO Brief Report. Multisystem Inflammatory Syndrome in Children and Adolescents with COVID-19. Available online: https://www.who.int/publications/i/item/multisystem-inflammatory-syndrome-in-children-and-adolescents-with-covid-19 (accessed on 15 January 2022).
- Centers for Disease Control and Prevention. MIS-C and COVID-19. Available online: https://www.cdc.gov/mis-c/ (accessed on 15 January 2022).
- Royal College of Paediatrics and Child Health Guidance: Paediatric Multisystem Inflammatory Syndrome Temporally Associated with COVID-19. Available online: https://www.rcpch.ac.uk/resources/paediatric-multisystem-inflammatory-syndrome-temporally-associated-covid-19-pims-guidance (accessed on 15 January 2022).
- Whittaker, E.; Bamford, A.; Kenny, J.; Kaforou, M.; Jones, C.E.; Shah, P.; Ramnarayan, P.; Fraisse, A.; Miller, O.; Davies, P.; et al. Clinical Characteristics of 58 Children with a Pediatric Inflammatory Multisystem Syndrome Temporally Associated with SARS-CoV-2. JAMA 2020, 324, 259–269. [Google Scholar] [CrossRef]
- Jones, V.G.; Mills, M.; Suarez, D.; Hogan, C.A.; Yeh, D.; Segal, J.B.; Nguyen, E.L.; Barsh, G.R.; Maskatia, S.; Mathew, R. COVID-19 and Kawasaki Disease: Novel Virus and Novel Case. Hosp Pediatr. 2020, 10, 537–540. [Google Scholar] [CrossRef]
- Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seibert, F.S.; Hoelzer, B.; Skrzypczyk, S.; Kohut, E.; Kurek, J.; Zapka, J.; et al. COVID-19 progression is potentially driven by T cell immunopathogenesis. medRxiv, 2020; preprint. [Google Scholar] [CrossRef]
- Hoste, L.; Van Paemel, R.; Haerynck, F. Multisystem Inflammatory Syndrome in Children Related to COVID-19: A Systematic Review. Eur. J. Pediatr. 2021, 180, 2019–2034. [Google Scholar] [CrossRef]
- Rowley, A.H. Understanding SARS-CoV-2-related multisystem inflammatory syndrome in children. Nat. Rev. Immunol. 2020, 20, 453–454. [Google Scholar] [CrossRef]
- Carter, M.J.; Shankar-Hari, M.; Tibby, S.M. Paediatric Inflammatory Multisystem Syndrome Temporally-Associated with SARS-CoV-2 Infection: An Overview. Intensiv. Care Med. 2021, 47, 90–93. [Google Scholar] [CrossRef]
- Masters, P.S. The Molecular Biology of Coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar] [CrossRef]
- Carter, M.J.; Fish, M.; Jennings, A.; Doores, K.J.; Wellman, P.; Seow, J.; Acors, S.; Graham, C.; Timms, E.; Kenny, J.; et al. Peripheral Immunophenotypes in Children with Multisystem Inflammatory Syndrome Associated With SARS-CoV-2 Infection. Nat. Med. 2020, 26, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Velikova, T.V.; Kotsev, S.V.; Georgiev, D.S.; Batselova, H.M. Immunological aspects of COVID-19: What do we know? World J. Biol. Chem. 2020, 11, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Tomov, L.P.; Velikova, T.V.; Batselova, H.M. Health Risk Management in Kindergartens, Schools and Universities, during COVID-19 Pandemic: A Heuristic Framework. Am. Int. J. Biol. Life Sci. 2021, 3, 40–43. [Google Scholar] [CrossRef]
- Weisberg, S.P.; Connors, T.J.; Zhu, Y.; Baldwin, M.R.; Lin, W.H.; Wontakal, S.; Szabo, P.A.; Wells, S.B.; Dogra, P.; Gray, J.; et al. Distinct antibody responses to SARS-CoV-2 in children and adults across the COVID-19 clinical spectrum. Nat. Immunol. 2021, 22, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.A.; Preston-Hurlburt, P.; Dai, Y.; Aschner, C.B.; Cheshenko, N.; Galen, B.; Garforth, S.J.; Herrera, N.G.; Jangra, R.K.; Morano, N.C.; et al. Immune responses to SARS-CoV-2 infection in hospitalized pediatric and adult patients. Sci. Transl. Med. 2020, 12, eabd5487. [Google Scholar] [CrossRef]
- Tosif, S.; Neeland, M.R.; Sutton, P.; Licciardi, P.V.; Sarkar, S.; Selva, K.J.; Do, L.; Donato, C.; Quan Toh, Z.; Higgins, R.; et al. Immune responses to SARS-CoV-2 in three children of parents with symptomatic COVID-19. Nat. Commun. 2020, 11, 5703. [Google Scholar] [CrossRef]
- Cohen, C.A.; Li, A.P.; Hachim, A.; Hui, D.S.; Kwan, M.Y.; Tsang, O.T.; Chiu, S.S.; Chan, W.H.; Yau, Y.S.; Kavian, N.; et al. SARS-CoV-2 specific T cell responses are lower in children and increase with age and time after infection. Nat. Commun. 2021, 12, 4678. [Google Scholar] [CrossRef]
- Filippatos, F.; Tatsi, E.; Michos, A. Immune response to SARS-CoV-2 in children: A review of the current knowledge. Pediatr. Investig. 2021, 5, 217–228. [Google Scholar] [CrossRef]
- Anaeigoudari, A.; Mollaei, H.R.; Arababadi, M.K.; Nosratabadi, R. Severe Acute Respiratory Syndrome Coronavirus 2: The Role of the Main Components of the Innate Immune System. Inflammation 2021, 44, 2151–2169. [Google Scholar] [CrossRef]
- Kasuga, Y.; Zhu, B.; Jang, K.-J.; Yoo, J.-S. Innate immune sensing of coronavirus and viral evasion strategies. Exp. Mol. Med. 2021, 53, 723–736. [Google Scholar] [CrossRef]
- Loske, J.; Röhmel, J.; Lukassen, S.; Stricker, S.; Magalhães, V.G.; Liebig, J.; Chua, R.L.; Thürmann, L.; Messingschlager, M.; Seegebarth, A.; et al. Pre-activated antiviral innate immunity in the upper airways controls early SARS-CoV-2 infection in children. Nat. Biotechnol. 2021, 40, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Geurtsen, J.; Driessen, N.N.; Appelmelk, B.J. Mannose–fucose recognition by DC-SIGN. In Microbial Glycobiology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 673–695. [Google Scholar]
- Han, D.P.; Lohani, M.; Cho, M.W. Specific asparagine-linked glycosylation sites are critical for DC-SIGN- and L-SIGN-mediated severe acute respiratory syndrome coronavirus entry. J. Virol. 2007, 81, 12029–12039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, L. The Membrane Protein of Severe Acute Respiratory Syndrome Coronavirus Functions as a Novel Cytosolic Pathogen-Associated Molecular Pattern to Promote Beta Interferon Induction via a Toll-Like-Receptor-Related TRAF3-Independent Mechanism. mBio 2016, 7, e01872-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.-Z.; Wang, S.-Y.; Zheng, Z.-Q.; Huang, Y.; Li, W.-W.; Xu, Z.-S.; Wang, Y.-Y. SARS-CoV-2 membrane glycoprotein M antagonizes the MAVS-mediated innate antiviral response. Cell. Mol. Immunol. 2021, 18, 613–620. [Google Scholar] [CrossRef]
- Zhou, J.H.; Wang, Y.N.; Chang, Q.Y.; Ma, P.; Hu, Y.; Cao, X. Type III Interferons in Viral Infection and Antiviral Immunity. Cell Physiol. Biochem. 2018, 51, 173–185. [Google Scholar] [CrossRef]
- Vono, M.; Huttner, A.; Lemeille, S.; Martinez-Murillo, P.; Meyer, B.; Baggio, S.; Sharma, S.; Thiriard, A.; Marchant, A.; Godeke, G.J.; et al. Robust innate responses to SARS-CoV-2 in children resolve faster than in adults without compromising adaptive immunity. Cell Rep. 2021, 37, 109773. [Google Scholar] [CrossRef]
- Worbs, T.; Hammerschmidt, S.I.; Förster, R. Dendritic cell migration in health and disease. Nat. Rev. Immunol. 2017, 17, 30–48. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. R. Soc. B 2015, 282, 20143085. [Google Scholar] [CrossRef]
- Asselin-Paturel, C.; Trinchieri, G. Production of type I interferons: Plasmacytoid dendritic cells and beyond. J. Exp. Med. 2005, 202, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Strauss-Ayali, D.; Conrad, S.M.; Mosser, D.M. Monocyte subpopulations and their differentiation patterns during infection. J. Leukoc. Biol. 2007, 82, 244–252. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Miller, L.C.; Blecha, F. Macrophage Polarization in Virus-Host Interactions. J. Clin. Cell Immunol. 2015, 6, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, C.; Benincasa, G.; Criscuolo, C.; Faenza, M.; Liberato, C.; Rusciano, M. Immune reactivity during COVID-19: Implications for treatment. Immunol. Lett. 2021, 231, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chu, H.; Hou, Y.; Chai, Y.; Shuai, H.; Lee, A.C.; Zhang, X.; Wang, Y.; Hu, B.; Huang, X.; et al. Attenuated Interferon and Proinflammatory Response in SARS-CoV-2-Infected Human Dendritic Cells Is Associated with Viral Antagonism of STAT1 Phosphorylation. J. Infect. Dis. 2020, 222, 734–745. [Google Scholar] [CrossRef]
- Dutt, T.S.; LaVergne, S.M.; Webb, T.L.; Baxter, B.A.; Stromberg, S.; McFann, K.; Berry, K.; Tipton, M.; Alnachoukati, O.; Zier, L.; et al. Comprehensive Immune Profiling Reveals CD56+ Monocytes and CD31+ Endothelial Cells Are Increased in Severe COVID-19 Disease. J. Immunol. 2022, 208, 685–696. [Google Scholar] [CrossRef]
- Moreews, M.; Le Gouge, K.; Khaldi-Plassart, S.; Pescarmona, R.; Mathieu, A.L.; Malcus, C.; Djebali, S.; Bellomo, A.; Dauwalder, O.; Perret, M.; et al. Polyclonal expansion of TCR Vbeta 21.3+ CD4+ and CD8+ T cells is a hallmark of Multisystem Inflammatory Syndrome in Children. Sci. Immunol. 2021, 6, eabh1516. [Google Scholar] [CrossRef]
- Yonker, L.M.; Neilan, A.M.; Bartsch, Y.; Patel, A.B.; Regan, J.; Arya, P.; Gootkind, E.; Park, G.; Hardcastle, M.; St John, A.; et al. Pediatric Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Clinical Presentation, Infectivity, and Immune Responses. J. Pediatr. 2020, 227, 45–52.e45. [Google Scholar] [CrossRef]
- Pierce, C.A.; Sy, S.; Galen, B.; Goldstein, D.Y.; Orner, E.; Keller, M.J.; Herold, K.C.; Herold, B.C. Natural mucosal barriers and COVID-19 in children. JCI Insight 2021, 6, e148694. [Google Scholar] [CrossRef]
- Park, A.; Iwasaki, A. Type I and type III interferons—Induction, signaling, evasion, and application to combat COVID-19. Cell Host Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V. Autoantibodies against type I IFNsin patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Vella, L.A.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Diorio, C.; Kuri-Cervantes, L.; Alanio, C.; Pampena, M.B.; Wu, J.E.; Chen, Z.; et al. Deep immune profiling of MIS-C demonstrates marked but transient immune activation compared to adult and pediatric COVID-19. Sci. Immunol. 2021, 6, eabf7570. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, A.; Brodsky, N.N.; Sumida, T.S.; Comi, M.; Asashima, H.; Hoehn, K.B.; Li, N.; Liu, Y.; Shah, A.; Ravindra, N.G.; et al. Immune dysregulation and autoreactivity correlate with disease severity in SARS-CoV-2-associated multisystem inflammatory syndrome in children. Immunity 2021, 54, 1083–1095.e7. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M. “Superantigens” may shed light on immune puzzle. Science 1990, 248, 685–686. [Google Scholar] [CrossRef]
- Brogan, P.A.; Shah, V.; Klein, N.; Dillon, M.J. V beta–restricted T cell adherence to endothelial cells: A mechanism for superantigen-dependent vascular injury. Arthritis Rheum. 2004, 50, 589–597. [Google Scholar] [CrossRef]
- Okarska-Napierała, M.; Ludwikowska, K.M.; Szenborn, L.; Dudek, N.; Mania, A.; Buda, P.; Książyk, J.; Mazur-Malewska, K.; Figlerowicz, M.; Szczukocki, M.; et al. Pediatric Inflammatory Multisystem Syndrome (PIMS) Did Occur in Poland during Months with Low COVID-19 Prevalence, Preliminary Results of a Nationwide Register. J. Clin. Med. 2020, 9, 3386. [Google Scholar] [CrossRef]
- Hoang, A.; Chorath, K.; Moreira, A.; Evans, M.; Burmeister-Morton, F.; Burmeister, F.; Naqvi, R.; Petershack, M.; Moreira, A. COVID-19 in 7780 pediatric patients: A systematic review. eClinicalMedicine 2020, 24, 100433. [Google Scholar] [CrossRef]
- Gruber, C.N.; Patel, R.S.; Trachtman, R.; Lepow, L.; Amanat, F.; Krammer, F.; Wilson, K.M.; Onel, K.; Geanon, D.; Tuballes, K.; et al. Mapping systemic inflammation and antibody responses in multisystem inflammatory syndrome in children (MIS-C). Cell 2020, 183, 982–995.e14. [Google Scholar] [CrossRef]
- Consiglio, C.R.; Cotugno, N.; Sardh, F.; Pou, C.; Amodio, D.; Rodriguez, L.; Tan, Z.; Zicari, S.; Ruggiero, A.; Pascucci, G.R.; et al. The Immunology of Multisystem Inflammatory Syndrome in Children with COVID-19. Cell 2020, 183, 968–981.e7. [Google Scholar] [CrossRef]
- Schvartz, A.; Belot, I.; Kone-Paut, I. Pediatric inflammatory multisystem syndrome and rheumatic diseases during SARS-CoV-2 pandemic. Front. Pediatr. 2020, 8, 833. [Google Scholar] [CrossRef] [PubMed]
- Van Caeseele, P.; Bailey, D.; Forgie, S.E.; Dingle, T.C.; Krajden, M.; Canadian Public Health Laboratory Network. SARS-CoV-2 (COVID-19) serology: Implications for clinical practice, laboratory medicine and public health. CMAJ 2020, 192, E973–E979. [Google Scholar] [CrossRef] [PubMed]
- Slabakova, Y.; Gerenska, D.; Ivanov, N.; Velikova, T. Immune titers of protection against severe acute respiratory syndrome coronavirus 2: Are we there yet? Explor. Immunol. 2022, 2, 9–24. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Du, R.H.; Zhang, W.; Si, H.R.; Zhu, Y.; Shen, X.R.; Li, Q.; Li, B.; Men, D.; et al. Serological investigation of asymptomatic cases of SARS-CoV-2 infection reveals weak and declining antibody responses. Emerg. Microb. Infect. 2021, 10, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Dowell, A.C.; Butler, M.S.; Jinks, E.; Tut, G.; Lancaster, T.; Sylla, P.; Begum, J.; Bruton, R.; Pearce, H.; Verma, K.; et al. Children develop robust and sustained cross-reactive spike-specific immune responses to SARS-CoV-2 infection. Nat. Immunol. 2022, 23, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Dijkman, R.; Jebbink, M.F.; El Idrissi, N.B.; Pyrc, K.; Müller, M.A.; Kuijpers, T.W.; Zaaijer, H.L.; van der Hoek, L. Human coronavirus NL63 and 229E seroconversion in children. J. Clin. Microbiol. 2008, 46, 2368–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkman, R.; Jebbink, M.F.; Gaunt, E.; Rossen, J.W.; Templeton, K.E.; Kuijpers, T.W.; van der Hoek, L. The dominance of human coronavirus OC43 and NL63 infections in infants. J. Clin. Virol. 2012, 53, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.T.; Garcia-Carreras, B.; Hitchings, M.; Yang, B.; Katzelnick, L.C.; Rattigan, S.M.; Borgert, B.A.; Moreno, C.A.; Solomon, B.D.; Trimmer-Smith, L.; et al. A systematic review of antibody-mediated immunity to coronaviruses: Kinetics, correlates of protection, and association with severity. Nat. Commun. 2020, 11, 4704. [Google Scholar] [CrossRef]
- Ng, K.W.; Faulkner, N.; Cornish, G.H.; Rosa, A.; Harvey, R.; Hussain, S.; Ulferts, R.; Earl, C.; Wrobel, A.G.; Benton, D.J.; et al. Preexisting and de novo humoral immunity to SARS-CoV-2 in humans. Science 2020, 370, 1339–1343. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lübke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Madera, S.; Crawford, E.; Langelier, C.; Tran, N.K.; Thornborrow, E.; Miller, S.; DeRisi, J.L. Nasopharyngeal SARS-CoV-2 viral loads in young children, do not differ significantly from those in older children and adults. Sci. Rep. 2021, 11, 3044. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Diorio, C.; Goodwin, E.C.; McNerney, K.O.; Weirick, M.E.; Gouma, S. SARS-CoV-2 antibody responses in children with MIS-C and mild and severe COVID-19. medRxiv, 2020; preprint. [Google Scholar] [CrossRef]
- Calder, P.C. Nutrition and immunity: Lessons for COVID-19. Nutr. Diabetes 2021, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Buonsenso, D.; Pujol, F.E.; Munblit, D.; Mcfarland, S.; Simpson, F. Clinical Characteristics, Activity Levels and Mental Health Problems in Children with Long COVID: A Survey of 510 Children. Preprints 2021, 2021030271. [Google Scholar] [CrossRef]
- Brackel, C.L.; Lap, C.R.; Buddingh, E.P.; van Houten, M.A.; van der Sande, L.J.; Langereis, E.J.; Bannier, M.A.; Pijnenburg, M.W.; Hashimoto, S.; Terheggen-Lagro, S.W. Pediatric long-COVID: An overlooked phenomenon? Pediatr. Pulmonol. 2021, 56, 2495–2502. [Google Scholar] [CrossRef]
- Borch, L.; Holm, M.; Knudsen, M.; Ellermann-Eriksen, S.; Hagstroem, S. Long COVID symptoms and duration in SARS-CoV-2 positive children- a nationwide cohort study. Eur. J. Pediatr. 2022, 181, 1597–1607. [Google Scholar] [CrossRef]
- Behnood, S.A.; Shafran, R.; Bennett, S.D.; Zhang, A.X.; O’Mahoney, L.L.; Stephenson, T.J.; Ladhani, S.N.; DeStavola, B.L.; Viner, R.M.; Swann, O.V. Persistent symptoms following SARS-CoV-2 infection amongst children and young people: A meta-analysis of controlled and uncontrolled studies. J. Infect. 2021, 84, 158–170. [Google Scholar] [CrossRef]
- McCrindle, B.W.; Manlhiot, C. SARS-CoV-2–related inflammatory, multisystem syndrome in children: Different or shared etiology and pathophysiology as Kawasaki disease? JAMA 2020, 324, 246–248. [Google Scholar] [CrossRef]
- Costagliola, G.; Spada, E.; Consolini, R. Severe COVID-19 in pediatric age: An update on the role of the anti-rheumatic agents. Pediatr. Rheumatol. 2021, 19, 68. [Google Scholar] [CrossRef] [PubMed]
- Porritt, R.A.; Paschold, L.; Rivas, M.N.; Cheng, M.H.; Yonker, L.M.; Chandnani, H.; Lopez, M.; Simnica, D.; Schultheiß, C.; Santiskulvong, C.; et al. HLA class I-associated expansion of TRBV11-2 T cells in multisystem inflammatory syndrome in children. J. Clin. Investig. 2021, 131, e146614. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| World Health Organization | Centers for Disease Control and Prevention (US) | Royal College of Pediatrics and Child Health (UK) |
|---|---|---|
| Six of 6 criteria must be met: 1. Age 0 to 19 years 2. Fever more than 3 days 3. Clinical signs of multisystem involvement (at least 2 of the following):
5. Excluding microbial cause of inflammation (bacterial sepsis and staphylococcal/streptococcal) toxic shock syndromes 6. Any of the following evidence of SARS-CoV-2 infection: RT-PCR, serology, antigen test, contact with a COVID-19 positive subject | 1. Age < 21 years 2. Presenting with Fever *, laboratory evidence of inflammation **, and evidence of clinically severe illness requiring hospitalization, with multisystem (>2) organ involvement (cardiac, renal, respiratory, hematologic, gastrointestinal, dermatologic or neurological); 3. No alternative plausible diagnoses; AND 4. Evidence for SARS-CoV-2 infection (recent or current) laboratory (RT-PCR, antigen test; serology) or epidemiology (exposure to COVID-19 four weeks before the symptoms onset | 1. Persistent fever > 38.5 °C 2. Evidence of single or multiorgan dysfunction: oxygen requirement, hypotension, abdominal pain, confusion, conjunctivitis, cough, diarrhea, headache, lymphadenopathy, mucus membrane changes, neck swelling, rash, respiratory symptoms, sore throat, swollen hands and feet, syncope, vomiting 3. Laboratory: abnormal fibrinogen, absence of potential causative organisms (other than SARS-CoV-2), hypoalbuminemia, lymphopenia, neutrophilia in most-normal neutrophils in some children; 4. Coagulopathy: high D-dimers, high ferritin, high IL-10 (if available), elevated IL-6 (if available) ***, high CRP neutrophilia, proteinuria, raised CK, raised LDH, raised triglycerides, raised troponin, thrombocytopenia, transaminitis 5./ECG and cardiac ultrasound/–evidence for cardiac involvement-myocardial, valves, pericardial, coronary arteries
|
| Expression Endosomal Surface | Name | Function | Associated Transcription Factors | Features |
| TLR3 | Recognition of dsRNA, produced during viral RNA replication | NF-κB IRF7 IRF3 |
| |
| TLR7 | Detection of ssRNA | |||
| TLR8 | Detection of ssRNA | |||
| TLR9 | incites by DNA viruses that contain unmethylated CpG DNA | |||
| Cell surface | TLR1 TLR2 TLR4 TLR5 TLR6 | Recognition of viral nucleic acids |
In a mice model of infected respiratory lungs (SARS and H1N1), the TLR4 mobilizations depend on oxidized phospholipids′ pulmonary expression | |
| Cytosol | RIG I | Detection of RNA containing 5′-triphosphate residue |
| |
| MDA-5 | long dsRNA |
| ||
| DC-SIGN; L-SIGN | C-type lectins; bind to carbohydrate residues; recognize the S protein RBD |
| ||
| ACE-2 | Binding to RBD domain of S protein |
|
| Type | Function |
|---|---|
| M1 macrophages | M1 macrophages present antigens to T cells, inhibit tumor growth, produce proinflammatory cytokines such as IL-6, IL-12, and TNF-α [35] M1 decreases during SARS-CoV-2 infection through apoptosis and necrosis [38]. On the other hand, the overactivation of M1 macrophages contribute to the development of severe disease course [23] |
| M2 macrophages | M2 macrophages are the key player in tissue repair and wound healing by producing anti-inflammatory cytokines such as IL-10 and TGF-β [36,37] |
| Classical monocytes CD14++CD16− | Phagocytic cells, which absorb pathogens; production of proinflammatory cytokines; activation of other immune cells [38] |
| Non-classical monocytes CD14+CD16++ SLAN (glycosylated expression form of P-selective glycoprotein-1 (PSGL-1) | SLAN + non-classical monocytes (type 2) are undetectable in both moderate and severe patients with COVID-19, while non-classical type 1 monocytes are elevated [23] |
| Intermediate monocytes CD14++CD16+ | Release of inflammatory cytokines—IL-1β, IL-6, IL-8 and TNF-α [38,39] Severe disease manifestation [23] |
| CD56+CD14+Ki67+IFN-γ+ monocyte | Produce IFN-g and Granzyme B in patients with moderate and severe COVID-19 [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazova, S.; Dimitrova, Y.; Hristova, D.; Tzotcheva, I.; Velikova, T. Cellular, Antibody and Cytokine Pathways in Children with Acute SARS-CoV-2 Infection and MIS-C—Can We Match the Puzzle? Antibodies 2022, 11, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11020025
Lazova S, Dimitrova Y, Hristova D, Tzotcheva I, Velikova T. Cellular, Antibody and Cytokine Pathways in Children with Acute SARS-CoV-2 Infection and MIS-C—Can We Match the Puzzle? Antibodies. 2022; 11(2):25. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11020025
Chicago/Turabian StyleLazova, Snezhina, Yulia Dimitrova, Diana Hristova, Iren Tzotcheva, and Tsvetelina Velikova. 2022. "Cellular, Antibody and Cytokine Pathways in Children with Acute SARS-CoV-2 Infection and MIS-C—Can We Match the Puzzle?" Antibodies 11, no. 2: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/antib11020025