
Sex-Related Left-Lateralized Development of the Crus II Region of the Ansiform Lobule in Cynomolgus Monkeys


1
Department of Nutrition, Faculty of Medical and Health Sciences, Tsukuba International University, Tsuchiura 300-0051, Japan
2
Department of Medical Physics and Engineering, Division of Health Sciences, Graduate School of Medicine, Osaka University, Osaka 565-0871, Japan
3
Department of Advanced Medical Technologies, National Cerebral and Cardiovascular Center Research Institute, Osaka 564-8565, Japan
*
Author to whom correspondence should be addressed.
Symmetry 2022, 14(5), 1015; https://0-doi-org.brum.beds.ac.uk/10.3390/sym14051015
Submission received: 25 March 2022
/
Revised: 30 April 2022
/
Accepted: 11 May 2022
/
Published: 16 May 2022
(This article belongs to the Special Issue Brain Asymmetry in Evolution II)
Abstract
:The asymmetric development of the cerebellum has been reported in several mammalian species. The current study quantitatively characterized cerebellar asymmetry and sexual dimorphism in cynomolgus macaques using magnetic resonance (MR) imaging-based volumetry. Three-dimensional T1W MR images at 7-tesla were acquired ex vivo from fixed adult male (n = 5) and female (n = 5) monkey brains. Five transverse domains of the cerebellar cortex, known as cerebellar compartmentation defined by the zebrin II/aldolase expression pattern, were segmented on MR images, and the left and right sides of their volumes were calculated. Asymmetry quotient (AQ) analysis revealed significant left-lateralization at the population level in the central zone posterior to the cerebellar transverse domains, which included lobule VII of the vermis with the crura I and II of ansiform lobules, in males but not females. Next, the volume of the cerebellar hemispherical lobules was calculated. Population-level leftward asymmetry was revealed in the crus II regions in males using AQ analysis. The AQ values of the other hemispherical lobules showed no left/right side differences at the population level in either sex. The present findings suggest a sexually dimorphic asymmetric aspect of the cerebellum in cynomolgus macaques, characterized by a leftward lateralization of the crus II region in males, but no left/right bias in females.
1. Introduction
Structural and functional lateralization of the brain is a distinctive aspect acquired during the evolutionary trajectory in mammals. Asymmetries in regional volumes and sulcal development of the cerebral cortex have been well documented in humans [1,2,3] and nonhuman primates [4,5,6]. Volume asymmetry in the cerebellum has also been reported in several mammalian species [7,8,9,10,11,12,13]. Some of these studies mentioned cerebellar asymmetry in relation to handedness/paw-use dominance [7,8,9,10]. On the other hand, asymmetrical cerebellar development is altered in human patients with neurodevelopmental and/or psychotic diseases showing cognitive impairments, such as autism [14,15] and schizophrenia [16]. The posterior cerebellum, including the ansiform lobules, is functionally lateralized to the left in association with cognition and spatial attention in humans [17,18,19] and to the right in association with working memory and language [20,21]. Thus, cerebellar lateralization plays a role in non-motor functions such as cognition and language, as well as handedness/paw-use dominance.
In our previous study using magnetic resonance imaging (MRI)-based volumetry, sexual dimorphism was revealed in the ferret cerebellum through observed leftward volume asymmetry in the posterior half in males but not in females [11,12]. In our definition, the lateralized cerebellar region, the central zone posterior (CZp), includes the cura I and II regions of the ansiform lobules [11]. Although the volume of the posterior cerebellum is entirely right-lateralized in humans [9], there have been no reports on sex-related asymmetric development of the cerebellar lobules in nonhuman primates. The current study quantitatively characterized cerebellar asymmetry and its sexual dimorphism in cynomolgus macaques (Macaca fascicularis) using MRI-based volumetry. Cerebellar asymmetry involves contralateral fiber connections to the cerebral cortex [21], which exhibits species-related presence or absence of sexual dimorphism of the morphological lateralization [1,6,22]. The current finding has evolutionary significance in sex-related structural asymmetry in the cerebellum in primates.
2. Materials and Methods
2.1. Samples
The present study used fixed brain samples from five sexually mature male and female cynomolgus monkeys (Macaca fascicularis) (3.5 to 6.6 years of age). These samples were used in our previous study [22], which was carried out in accordance with the Guide for the Care and Use of Laboratory Animals by the National Institutes of Health (NIH, Bethesda, MD, USA) and the ethics criteria stated in the bylaws of the Experimental Animal Ethics Committee of Shin Nippon Biomedical Laboratories. The study was approved by the Institutional Animal Care and Use Committee of Shin Nippon Biomedical Laboratories (Approval code: B999-178). The tissue was fixed using intracardiac perfusion with 0.9% NaCl followed by 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) under deep anesthesia with an intravenous injection of sodium pentobarbital (26 mg/kg; Tokyo Chemical Industry, Tokyo, Japan) as described in a previous report [22], and immersed in the same fixative. Anatomical magnetic resonance imaging (MRI) was performed using a horizontal 7.0 T scanner (BioSpec 70/30 USR; Bruker Biospin, Ettlingen, Germany) with an 86 mm volume coil. This was in order to acquire three-dimensional (3D) MR images covering the entire fixed brain using rapid acquisition with a relaxation enhancement (RARE) sequence with the following parameters: repetition time (TR) = 400 ms; echo time (TE) = 6 ms (effective TE = 19.2 ms); RARE factor = 4; field of view (FOV) = 72 × 64 × 47.5 mm3; acquisition matrix = 288 × 256 × 192; voxel size = 250 × 250 × 250 µm3; number of acquisitions (NEX) = 4; and total scan time = 5 h, 27 min, and 36 s.
2.2. Volumetry
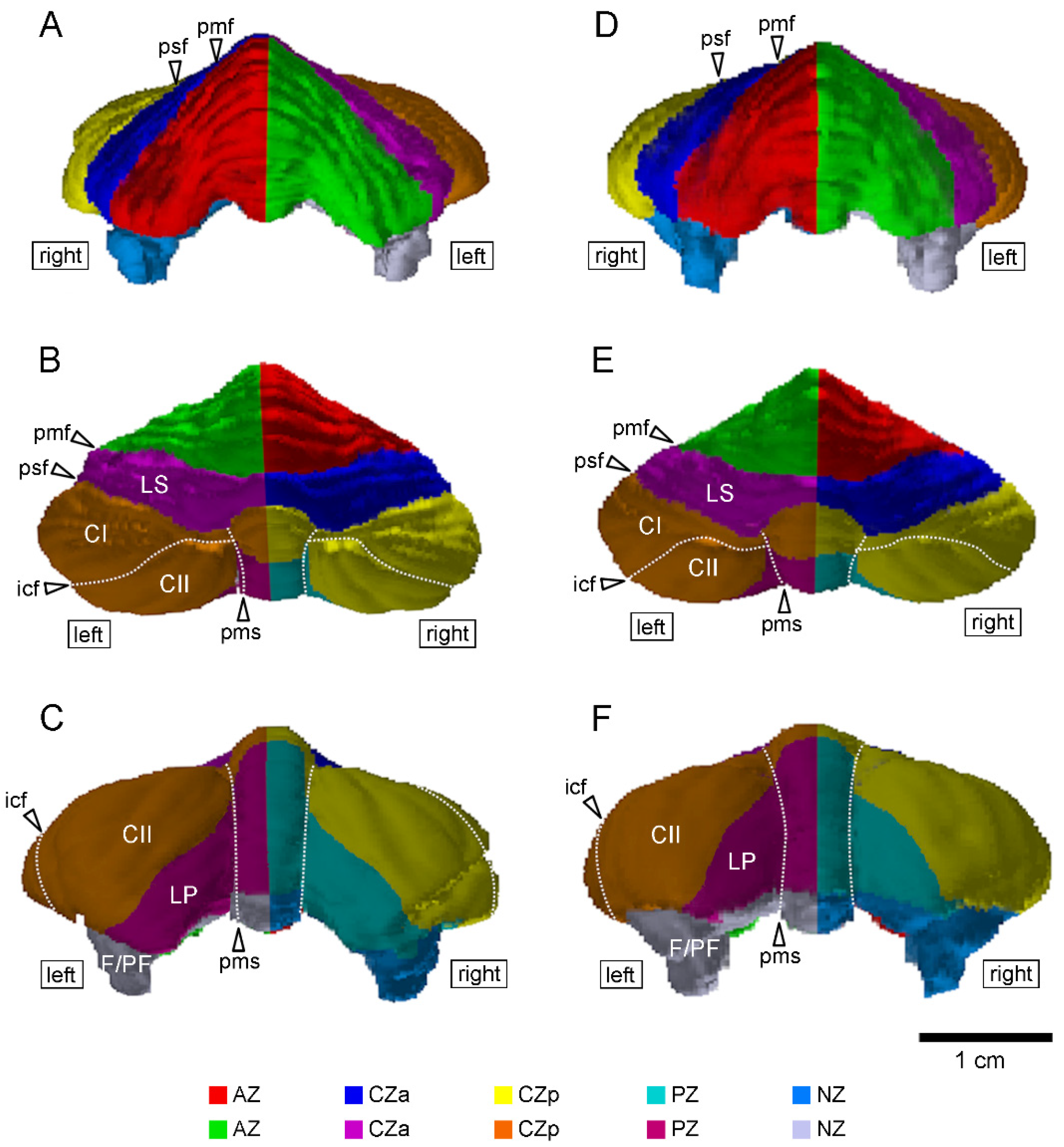
Volumetric analysis was conducted on 3D MR images covering the entire cerebellum in accordance with our previous procedure [11]. Cerebellar transverse domains were defined primarily based on the zebrin II/aldolase C expression pattern [23], which is highly conserved among mammalian species [24,25,26]. The transverse domains included the anterior zone (AZ; lobules I–V), central zone anterior (CZa; lobule VI and the lobules simplex), central zone posterior (CZp; lobule VII and ansiform lobules), posterior zone (PZ; lobules VIII–IXa and the paramedian lobule), and nodular zone (NZ; flocculus and paraflocculus) [23], and these were further divided into the left and right sides at the midline, as defined by the position of the cerebral longitudinal fissure [11]. The left and right sides of these cerebellar cortex domains were semi-automatically segmented, and their areas were measured using SliceOmatic software version 4.3 (TomoVision, Montreal, QC, Canada). The volume (mm3) was then calculated by multiplying the combined areas by the slice thickness (250 μm). We further estimated the volumes of the cerebellar hemispherical lobules, which could be delineated boundaries by cerebellar fissures/sulci, as shown in Figure 1 and Figure 2. The asymmetry quotient (AQ) values of the cerebellar transverse domains and hemispherical lobules were calculated using the formula ((R − L)/{(R + L) × 0.5}). The asymmetry direction indicated a rightward bias when AQ values were positive and a leftward bias when AQ values were negative [27]. The segmented images were further used to construct 3D volume-rendered images of the cerebellar transverse domains using the 3D-rendering module of the SliceOmatic software version 4.3 (TomoVision, Montreal, QC, Canada).
2.3. Statistical Analysis
Sex differences in the volumes of the cerebellar transverse domains and hemispherical lobules were statistically evaluated using one-way ANOVA, followed by a two-tailed Student’s t-test. The left/right side difference in the whole cerebellar volume was calculated using a two-tailed, paired-sample Student’s t-test. We further assessed the sex-related changes in the volume laterality of the cerebellar transverse domains and hemispherical lobules using repeated measures three-way ANOVA using sex as an intergroup factor and the cerebellar regions and left/right sides as intragroup factors. Sexual differences were assessed using Scheffe’s test, and left/right side differences were compared using a paired sample t-test for post-hoc testing, following simple main effects at p < 0.05. The AQ values of the cerebellar transverse domains and hemispherical lobules were analyzed using a one-sample t-test to determine any significant population-level asymmetry.
3. Results
3.1. Cerebellar Volumes of Males and Females
The volumes of the whole cerebellum and the five cerebellar transverse domains in male and female cynomolgus monkeys are summarized in Table 1. There was no statistically significant difference in the whole cerebellar volume between males (3441 ± 147 mm3) and females (3338 ± 204 mm3) according to Student’s t-test. The sexual differences in volume were not statistically significant when the cerebellum was divided into five transverse domains. We further estimated the volumes of the cerebellar hemispherical lobules, which could be delineated boundaries by cerebellar fissures/sulci, as shown in Figure 1 and Figure 2. The volumes did not differ between the sexes in the crura I and II regions of the ansiform lobules, paramedian lobule, and flocculus/paraflocculus by the Student’s t-test (Table 2).
3.2. Left/Right-Side Differences in Cerebellar Volumes
There was no left/right side difference in the volume of the whole cerebellum in either male or female cynomolgus monkeys by paired sample Student’s t-test (Table 3). The left bias of the whole cerebellar volume was revealed by the AQ analysis, but a significant population level asymmetry was not detected by the one-sample t-test (Table 3).
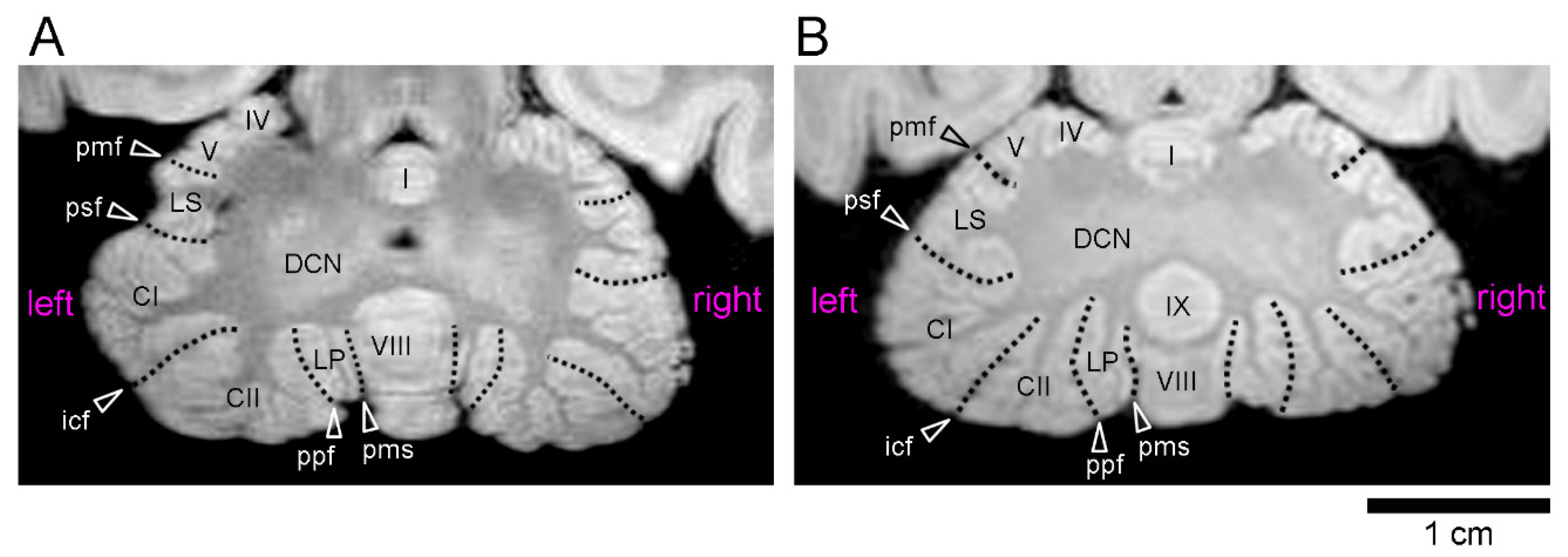
The volumes of the five cerebellar transverse domains were compared between the left and right sides. Repeated-measures three-way ANOVA revealed significant effects in the left/right sides [F(1,8) = 9.282; p < 0.05], cerebellar transverse domains [F(4,32) = 195.805; p < 0.001], and an interaction between the left/right sides and cerebellar transverse domains [F(4,32) = 9.282; p < 0.01]. Following significant simple main effects, the paired sample t-test indicated significantly larger volumes on the left side than on the right side in the CZp of the males (p < 0.01) and in the AZ of the females (p < 0.01) (Table 3). Notably, a significant left bias of the male CZp volume (p < 0.05), but not the female AZ volume, was detected at the population level by the one-sample t-test (Table 3). We further calculated the volumes of the left/right of the cerebellar hemispherical lobules in the CZp (crura I and II), PZ (paramedian lobule), and NZ (flocculus and paraflocculus) regions. The repeated measures three-way ANOVA revealed significant effects on the left/right sides [F(1,8) = 5.614; p < 0.05] and in the cerebellar hemispherical lobules [F(3,24) = 27.861; p < 0.001]. A significantly larger volume was detected in male crus II regions on the left side compared to on the right side, as determined by the paired sample t-test (p < 0.05) (Table 4), following a significant simple main effect (p < 0.01). A significant left bias indicated by AQ values in the male crus II region volumes was revealed at the population level by the one-sample t-test (p < 0.01) (Table 4). This left-lateralization of the male crus II region could be confirmed optically on MR images (Figure 2).
4. Discussion
The cerebellar volume is right-lateralized, particularly in the posterior cerebellum, in humans, forming a characteristic clockwise torque asymmetry [9]. Although cerebellar volume asymmetry has been reported to be associated with handedness in humans [7,9] and nonhuman primates [8], the function of the posterior cerebellum, including the ansiform lobules, is right-biased in relation to language in humans [17,18,19]. Such cerebellar functional asymmetry involves contralateral fiber connections to the left association cortex [21,28]. A notable finding of the present investigation was the leftward volume asymmetry of the CZp, particularly in the crus II region, in male cynomolgus monkeys but not in females. In our previous study, the arcuate sulcus, which encompasses the posterior borders of the dorsolateral and ventrolateral prefrontal cortex (dlPFC and vlPFC) [29] was right-lateralized in its length in male cynomolgus monkeys; however, this region was symmetrical in females [22]. The crus II region is involved in cognitive control via the frontoparietal network, including the dlPFC [30]. The functional implications of sex differences in crus II volume asymmetry are unclear. One possibility is that an increased linkage of the left crus II to the right dlPFC is associated with stronger desires and reward-seeking behaviors in males by reducing the relative activity of the contralateral (left) side of the dlPFC [31,32]. Further investigation is required in this context.
The present study applied cerebellar transverse domains, which were defined primarily on the basis of the zebrin II/aldolase C expression pattern [23], to evaluate cerebellar volume asymmetry. Since the expression pattern of zebrin II/aldolase C in the cerebellum is highly conserved among mammalian species [24,25,26], use of the cerebellar transverse domains allows for comparison of cerebellar volume asymmetry among mammalian species. The current investigation revealed a male-preferred left-lateralized CZp volume in cynomolgus monkeys, reminiscent of the findings obtained from ferrets [11,12]. Although cerebellar volume asymmetry has not been evaluated using cerebellar transverse domains in humans, the posterior cerebellum, except for the crus II region, exhibited higher leftward asymmetry in males than in females [33,34]. These findings suggest that male-preferred leftward development of the posterior cerebellum (CZp in our definition) is phylogenetically conserved among ferrets, cynomolgus macaques, and humans. In contrast, the leftward asymmetry of the posterior cerebellum in humans may have been lost due to the expansion of language-related regions in the right crus II region [33,34]. Notably, the volume of the posterior cerebellum is right-lateralized in humans [9].
Another sex difference reported in humans [35,36,37] and ferrets [11,12] is the larger size and volume of the whole cerebellum in males compared to in females, as well as those in the whole brain. The sexual differences in cerebellar volume in humans have been attributed to X-chromosome escape genes, independent of perinatal and/or post-pubertal androgens [33,38]. The whole volume of the cerebellum did not differ between sexes in cynomolgus monkeys in the present study, although significantly greater brain weight with cerebral size was observed in males than in females [22]. No sexual difference in cerebellar volume was noted in both dogs [10] and mice [39]. However, brain weight was significantly greater in male dogs [10], but not sexually dimorphic in mice [40]. These suggests that a genetically altered, male-over-female larger cerebellar size/volume is a species-specific trait but not a phylogenetically conserved trait among mammalian species.
5. Conclusions
A striking aspect of cerebellar asymmetry has been reported in humans [9], nonhuman primates [8] and carnivores [10,11,12,13]. The direction of cerebellar asymmetry varies depending on the species and/or sexes [7,8,9,10,11,12,13]. Notably, the population-level volume asymmetry in the posterior cerebellum, including the CZp in our definition, changed from leftward to rightward after the split between cynomolgus macaques (the present results) and humans [9]. On the other hand, the cerebellar asymmetry is disturbed in male-preferred or male-earlier onset neurodevelopmental and/or psychotic diseases in humans, such as autism [14,15] and schizophrenia [14]. The current findings provide keys to understanding the evolution of the mammalian brain and the pathogenesis of human neurodevelopmental and/or psychotic diseases on the basis of cerebellar asymmetry.
Author Contributions
K.S. conceived and designed the experiments; K.S. and S.S. performed the experiments and analyzed the data; K.S. and S.S. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was the result of using research equipment shared by the MEXT Project for promoting public utilization of advanced research infrastructure (program for supporting the construction of core facilities) (grant number JPMXS0450400021).
Institutional Review Board Statement
The study was approved by the Institutional Animal Care and Use Committee of Shin Nippon Biomedical Laboratories (Approval code: B999-178).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Good, C.D.; Johnsrude, I.; Ashburner, J.; Henson, R.N.; Friston, K.J.; Frackowiak, R.S. Cerebral asymmetry and the effects of sex and handedness on brain structure: A voxel-based morphometric analysis of 465 normal adult human brains. Neuroimage 2001, 14, 685–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, M.A.; Lombardino, L.J.; Walczak, A.R.; Bonihla, L.; Leonard, C.M.; Binder, J.R. Manual and automated measures of superior temporal gyrus asymmetry: Concordant structural predictors of verbal ability in children. Neuroimage 2008, 41, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, M.A.; Iuricich, F.; Vaden , K.I., Jr.; Glaze, B.T. Dyslexia Data Consortium. The topology of pediatric structural asymmetries in language-related cortex. Symmetry 2020, 12, 1809. [Google Scholar] [CrossRef] [PubMed]
- Falk, D.; Hildebolt, C.; Cheverud, J.; Vannier, M.; Helmkamp, R.C.; Konigsberg, L. Cortical asymmetries in frontal lobes of rhesus monkeys (Macaca mulatta). Brain Res. 1990, 512, 40–45. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Nir, T. Planum temporale surface area and grey matter asymmetries in chimpanzees (Pan troglodytes): The effect of handedness and comparison within findings in humans. Behav. Brain Res. 2010, 208, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Sawada, K. Cerebral sulcal asymmetry in macaque monkeys. Symmetry 2020, 12, 1509. [Google Scholar] [CrossRef]
- Synder, P.J.; Bilder, R.M.; Wu, H.; Bogerts, B.; Lieberman, J.A. Cerebellar volume asymmetries are related to handedness: A quantitative MRI study. Neuropsychologia 1995, 33, 407–419. [Google Scholar] [CrossRef]
- Phillips, K.; Hopkins, W.D. Exploring the relationship between cerebellar asymmetry and handedness in chimpanzees (Pan troglodytes) and capuchins (Cebus apella). Neuropsychologia 2007, 45, 2333–2339. [Google Scholar] [CrossRef] [Green Version]
- Rosch, R.E.; Ronan, L.; Cherkas, L.; Gurd, J.M. Cerebellar asymmetry in a pair of monozygotic handedness-discordant twins. J. Anat. 2010, 217, 38–47. [Google Scholar] [CrossRef]
- Koyun, N.; Aydinlioğlu, A.; Aslan, K. A morphometric study on dog cerebellum. Neurol. Res. 2011, 33, 220–224. [Google Scholar] [CrossRef]
- Sawada, K.; Horiuchi-Hirose, M.; Saito, S.; Aoki, I. Male prevalent enhancement of leftward asymmetric development of the cerebellar cortex in ferrets (Mustela putorius). Laterality 2015, 20, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Aoki, I. Age-dependent sexually-dimorphic asymmetric development of the ferret cerebellar cortex. Symmetry 2017, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Sawada, K.; Kamiya, S.; Aoki, I. Asymmetry of cerebellar lobular development in ferrets. Symmetry 2020, 12, 735. [Google Scholar] [CrossRef]
- Hodge, S.M.; Makris, N.; Kennedy, D.N.; Caviness, V.S., Jr.; Howard, J.; McGrath, L.; Steele, S.; Frazier, J.A.; Tager-Flusberg, H.; Harris, G.J. Cerebellum, language, and cognition in autism and specific language impairment. J. Autism Dev. Disord. 2010, 40, 300–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, Q.; Zuo, C.; Zhao, L.; Hao, L. Longitudinal changes of cerebellar thickness in autism. spectrum disorder. Neurosci. Lett. 2020, 728, 134949. [Google Scholar] [CrossRef]
- Szeszko, P.R.; Gunning-Dixon, F.; Ashtari, M.; Snyder, P.J.; Lieberman, J.A.; Bilder, R.M. Reversed cerebellar asymmetry in men with first-episode schizophrenia. Biol. Psychiatry 2003, 53, 450–459. [Google Scholar] [CrossRef]
- Hu, D.; Shen, H.; Zhou, Z. Functional asymmetry in the cerebellum: A brief review. Cerebellum 2008, 7, 304–313. [Google Scholar] [CrossRef]
- Iglói, K.; Doeller, C.F.; Paradis, A.L.; Benchenane, K.; Berthoz, A.; Burgess, N.; Rondi-Reig, L. Interaction between hippocampus and cerebellum crus I in sequence-based but not place-based navigation. Cereb. Cortex 2015, 25, 4146–4154. [Google Scholar] [CrossRef] [Green Version]
- Ciricugno, A.; Ferrari, C.; Rusconi, M.L.; Cattaneo, Z. The left posterior cerebellum is involved in orienting attention along the mental number line: An online-TMS study. Neuropsychologia 2020, 143, 107497. [Google Scholar] [CrossRef]
- Valera, E.M.; Faraone, S.V.; Biederman, J.; Poldrack, R.A.; Seidman, L.J. Functional neuroanatomy of working memory in adults with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 439–447. [Google Scholar] [CrossRef]
- Wang, D.; Buckner, R.L.; Liu, H. Cerebellar asymmetry and its relation to cerebral asymmetry estimated by intrinsic functional connectivity. J. Neurophysiol. 2013, 109, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Imai, N.; Sawada, K.; Fukunishi, K.; Sakata-Haga, H.; Fukui, Y. Sexual dimorphism of sulcal length asymmetry in cerebrum of adult cynomolgus monkeys (Macaca fascicularis). Congenit. Anom. 2011, 51, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Sillitoe, R.V.; Hawkes, R. Whole-mount immunohistochemistry: A high-throughput screen for patterning defects in the mouse cerebellum. J. Histochem. Cytochem. 2002, 50, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Sillitoe, R.V.; Hulliger, M.; Dyck, R.; Hawkes, R. Antigenic compartmentation of the cat cerebellar cortex. Brain Res. 2003, 977, 1–15. [Google Scholar] [CrossRef]
- Sillitoe, R.V.; Malz, C.R.; Rockland, K.; Hawkes, R. Antigenic compartmentation of the primate and tree shrew cerebellum: A common topography of zebrin II in Macaca mulatta and Tupaia belangeri. J. Anat. 2004, 204, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Hawkes, R. On the architecture of the posterior zone of the cerebellum. Cerebellum 2011, 10, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Marino, L. Asymmetries in cerebral width in nonhuman primate brains as revealed by magnetic resonance imaging (MRI). Neuropsychologia 2000, 38, 493–499. [Google Scholar] [CrossRef]
- Siniscalchi, M.; Franchini, D.; Pepe, A.M.; Sasso, R.; Dimatteo, S.; Vallortigara, G.; Quaranta, A. Volumetric assessment of cerebral asymmetries in dogs. Laterality 2011, 16, 528–536. [Google Scholar] [CrossRef]
- Miller, E.K.; Walls, J.D. The frefrontal cortex and executive brain functions. In Fundamental Neuroscience, 2nd ed.; Squire, L., Berg, D., Bloom, F.E., McConnell, S., Roberts, J.L., Spitzer, N.C., Zigmond, M.J., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 1353–1376. [Google Scholar]
- Sang, L.; Qin, W.; Liu, Y.; Han, W.; Zhang, Y.; Jiang, T.; Yu, C. Resting-state functional connectivity of the vermal and hemispheric subregions of the cerebellum with both the cerebral cortical networks and subcortical structures. Neuroimage 2012, 61, 1213–1225. [Google Scholar] [CrossRef]
- Goldstein, R.Z.; Volkow, N.D. Dysfunction of the prefrontal cortex in addiction: Neuroimaging findings and clinical implications. Nat. Rev. Neurosci. 2011, 12, 652–669. [Google Scholar] [CrossRef]
- Sofuoglu, M.; DeVito, E.E.; Waters, A.J.; Carroll, K.M. Cognitive enhancement as a treatment for drug addictions. Neuropharmacology 2013, 64, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentini, E.; Kasahara, M.; Arver, S.; Savic, I. Sex differences in the human brain and the impact of sex chromosomes and sex hormones. Cereb. Cortex 2013, 23, 2322–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Tang, Y.; Sun, B.; Gong, G.; Chen, Z.J.; Lin, X.; Yu, T.; Li, Z.; Evans, A.C.; Liu, S. Sexual dimorphism and asymmetry in human cerebellum: An MRI-based morphometric study. Brain Res. 2010, 1353, 60–73. [Google Scholar] [CrossRef]
- Raz, N.; Dupuis, J.H.; Briggs, S.D.; McGavran, C.; Acker, J.D. Differential effects of age and sex on the cerebellar hemispheres and the vermis: A prospective MR study. AJNR Am. J. Neuroradiol. 1998, 19, 65–71. [Google Scholar] [PubMed]
- Raz, N.; Gunning-Dixon, F.; Head, D.; Williamson, A.; Acker, J.D. Age and sex differences in the cerebellum and the ventral pons: A prospective MR study of healthy adults. AJNR Am. J. Neuroradiol. 2001, 22, 1161–1167. [Google Scholar]
- Giedd, J.N.; Raznahan, A.; Mills, K.L.; Lenroot, R.K. Review: Magnetic resonance imaging of male/female differences in human adolescent brain anatomy. Biol. Sex Differ. 2012, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, S.G.; Read, G.; Pan, H.; Fear, C.; Lindenbaum, R.; Crossley, J. Prenatal testosterone levels in XXY and XYY males. Horm. Res. 1994, 42, 106–109. [Google Scholar] [CrossRef]
- Dorr, A.E.; Lerch, J.P.; Spring, S.; Kabani, N.; Henkelman, R.M. High resolution three-dimensional brain atlas using an average magnetic resonance image of 40 adult C57Bl/6J mice. Neuroimage 2008, 42, 60–69. [Google Scholar] [CrossRef]
- Bishop, K.M.; Wahlsten, D. Sex and species differences in mouse and rat forebrain commissures depend on the method of adjusting for brain size. Brain Res. 1999, 815, 358–366. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Three-dimensional volume-rendered images of the cerebella of young adult cynomolgus monkeys. (A–C) Anterior, dorsal, and posterior views in the male cerebellum. (D–F) Anterior, dorsal, and posterior views in the female cerebellum. The cerebellar cortex was divided into five transverse domains: the left and right sides of the anterior zone (AZ) (vermal lobules I–V), central zone anterior (CZa; vermal lobule VI and lobules simplex), central zone posterior (CZp; vermal lobule VII, and the crura I and II regions of the ansiform lobules), posterior zone (PZ; vermal lobules VIII–IXa, and paramedian lobule), and nodular zone (NZ; vermal lobules IXb–X, paraflocculus and flocculus). The left and right sides are divided at midline, which was defined by the position of the cerebral longitudinal fissure. Dot lines delineate the intercrucial fissure (icf) and paramedian sulcus (pms). CI—crus I of ansiform lobule; CII—crus II of ansiform lobule; F—flocculus; LP—paramedian lobule; LS—lobulus simplex; PF—paraflocculus; pmf—primary fissure; psf—posterior superior fissure.
Figure 1.
Three-dimensional volume-rendered images of the cerebella of young adult cynomolgus monkeys. (A–C) Anterior, dorsal, and posterior views in the male cerebellum. (D–F) Anterior, dorsal, and posterior views in the female cerebellum. The cerebellar cortex was divided into five transverse domains: the left and right sides of the anterior zone (AZ) (vermal lobules I–V), central zone anterior (CZa; vermal lobule VI and lobules simplex), central zone posterior (CZp; vermal lobule VII, and the crura I and II regions of the ansiform lobules), posterior zone (PZ; vermal lobules VIII–IXa, and paramedian lobule), and nodular zone (NZ; vermal lobules IXb–X, paraflocculus and flocculus). The left and right sides are divided at midline, which was defined by the position of the cerebral longitudinal fissure. Dot lines delineate the intercrucial fissure (icf) and paramedian sulcus (pms). CI—crus I of ansiform lobule; CII—crus II of ansiform lobule; F—flocculus; LP—paramedian lobule; LS—lobulus simplex; PF—paraflocculus; pmf—primary fissure; psf—posterior superior fissure.

Figure 2.
Ex vivo MR images (using a RARE sequence with a short TR and the minimum TE settings) of cerebella through the deep cerebellar nuclei in young adult cynomolgus monkeys. (A) Axial MR image of the male cerebellum. (B) Axial MR image of the female cerebellum. Abbreviations are posted at the left side. Roman numerals identify the vermal lobules. CI—crus I of ansiform lobule; CII—crus II of ansiform lobule; DCN—deep cerebellar nuclei; icf—intercrucial fissure; LP—paramedian lobule; LS—lobulus simplex; pmf—primary fissure; pms—paramedian sulcus; ppt—prepyramidal fissure; psf—posterior superior fissure.
Figure 2.
Ex vivo MR images (using a RARE sequence with a short TR and the minimum TE settings) of cerebella through the deep cerebellar nuclei in young adult cynomolgus monkeys. (A) Axial MR image of the male cerebellum. (B) Axial MR image of the female cerebellum. Abbreviations are posted at the left side. Roman numerals identify the vermal lobules. CI—crus I of ansiform lobule; CII—crus II of ansiform lobule; DCN—deep cerebellar nuclei; icf—intercrucial fissure; LP—paramedian lobule; LS—lobulus simplex; pmf—primary fissure; pms—paramedian sulcus; ppt—prepyramidal fissure; psf—posterior superior fissure.


{kind=link}
{kind=link}
Table 1.
Volumes of the whole cerebellum and five cerebellar transverse domains in young adult cynomolgus monkeys.
Table 1.
Volumes of the whole cerebellum and five cerebellar transverse domains in young adult cynomolgus monkeys.
| Males (n = 5) | Females (n = 5) | |
|---|---|---|
| Volumes (mm3) | ||
| Whole cerebellum | 3441 ± 147 | 3338 ± 204 |
| Anterior zone | 916 ± 81 | 844 ± 68 |
| Central zone anterior | 491 ± 46 | 489 ± 48 |
| Central zone posterior | 1050 ± 86 | 1033 ± 70 |
| Posterior zone | 493 ± 34 | 442 ± 57 |
| Nodular zone | 492 ± 87 | 530 ± 36 |
Data are represented as mean ± SD.
Table 2.
Volumes of cerebellar hemispherical lobules in young adult cynomolgus monkeys.
| Males (n = 5) | Females (n = 5) | ||
|---|---|---|---|
| Volumes (mm3) | |||
| Crus I | 520 ± 66 | 541 ± 44 | |
| Crus II | 455 ± 92 | 409 ± 64 | |
| Paramedian lobule | 286 ± 29 | 247 ± 31 | |
| Flocculus/Paraflocculus | 404 ± 83 | 436 ± 22 | |
Data are represented as mean ± SD.
Table 3.
Left/right side differences in the volumes of the whole cerebellum and five cerebellar transverse domains in adult male cynomolgus monkeys.
Table 3.
Left/right side differences in the volumes of the whole cerebellum and five cerebellar transverse domains in adult male cynomolgus monkeys.
| Left Side (mm3) | Right Side(mm3) | AQ Value | |
|---|---|---|---|
| Whole cerebellum | |||
| Males (n = 5) | 1791 ± 116 | 1650 ± 67 | −0.080 ± 0.070 |
| Females (n = 5) | 1703 ± 126 | 1635 ± 96 | −0.040 ± 0.056 |
| Anterior zone | |||
| Males (n = 5) | 479 ± 64 | 437 ± 24 | −0.085 ± 0.118 |
| Females (n = 5) | 439 ± 40 * | 404 ± 33 * | −0.083 ± 0.068 |
| Central zone anterior | |||
| Males (n = 5) | 252 ± 30 | 239 ± 21 | −0.048 ± 0.111 |
| Females (n = 5) | 245 ± 27 | 243 ± 23 | 0.007 ± 0.062 |
| Central zone posterior | |||
| Males (n = 5) | 547 ± 47 ** | 502 ± 41 ** | −0.085 ± 0.030 # |
| Females (n = 5) | 536 ± 51 | 497 ± 40 | −0.075 ± 0.114 |
| Posterior zone | |||
| Males (n = 5) | 251 ± 16 | 243 ± 31 | −0.038 ± 0.141 |
| Females (n = 5) | 215 ± 34 | 227 ± 29 | 0.058 ± 0.125 |
| Nodular zone | |||
| Males (n = 5) | 262 ± 51 | 229 ± 43 | −0.131 ± 0.135 |
| Females (n = 5) | 267 ± 15 | 264 ± 23 | −0.014 ± 0.055 |
Data are represented as the mean ± SD. * p < 0.05, ** p < 0.01 (left vs. right, paired sample Student’s t-test); # p < 0.05 (One-sample t-test).
Table 4.
Left/right side differences in the volumes of the cerebellar hemispherical lobules in adult male cynomolgus monkeys.
Table 4.
Left/right side differences in the volumes of the cerebellar hemispherical lobules in adult male cynomolgus monkeys.
| Left Side | Right Side | AQ Value | |
|---|---|---|---|
| Crus I | |||
| Males (n = 5) | 263 ± 24 | 257 ± 43 | −0.030 ± 0.094 |
| Females (n = 5) | 277 ± 33 | 267 ± 17 | −0.044 ± 0.095 |
| Crus II | |||
| Males (n = 5) | 240 ± 52 * | 215 ± 41 * | −0.108 ± 0.034 # |
| Females (n = 5) | 213 ± 33 | 196 ± 38 | −0.087 ± 0.165 |
| Paramedian lobule | |||
| Males (n = 5) | 141 ± 11 | 145 ± 20 | 0.025 ± 0.112 |
| Females (n = 5) | 119 ± 21 | 127 ± 11 | 0.072 ± 0.094 |
| Flocculus/Paraflocculus | |||
| Males (n = 5) | 211 ± 48 | 193 ± 40 | −0.081 ± 0.136 |
| Females (n = 5) | 222 ± 11 | 214 ± 19 | −0.034 ± 0.100 |
Data are represented as the mean ± SD. * p < 0.05 (left vs. right, paired sample Student’s t-test); # p < 0.01 (one-sample t-test).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sawada, K.; Saito, S. Sex-Related Left-Lateralized Development of the Crus II Region of the Ansiform Lobule in Cynomolgus Monkeys. Symmetry 2022, 14, 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14051015
AMA Style
Sawada K, Saito S. Sex-Related Left-Lateralized Development of the Crus II Region of the Ansiform Lobule in Cynomolgus Monkeys. Symmetry. 2022; 14(5):1015. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14051015
Chicago/Turabian StyleSawada, Kazuhiko, and Shigeyoshi Saito. 2022. "Sex-Related Left-Lateralized Development of the Crus II Region of the Ansiform Lobule in Cynomolgus Monkeys" Symmetry 14, no. 5: 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14051015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.