
Asymmetric Prefrontal Cortex Activation Associated with Mutual Gaze of Mothers and Children during Shared Play

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
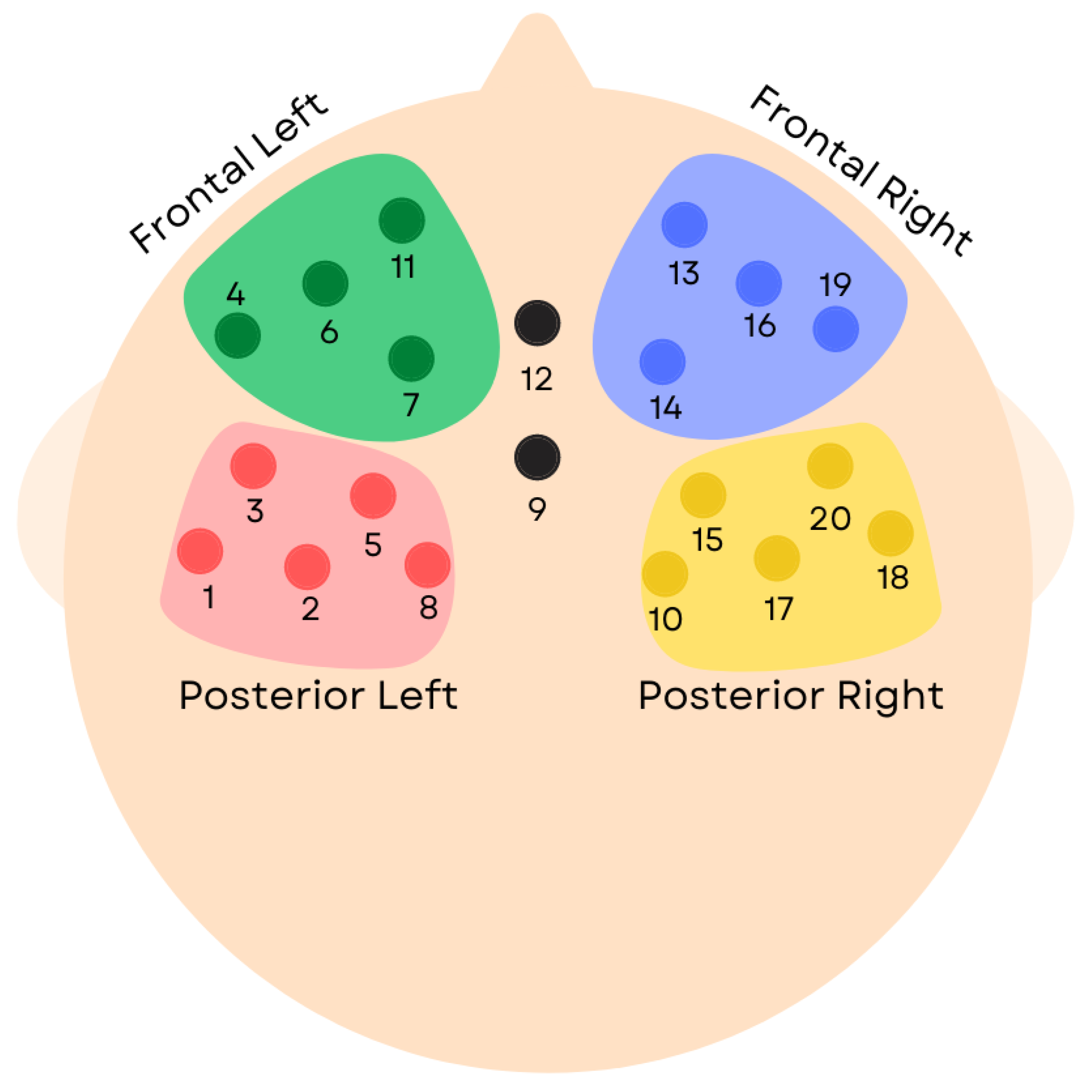
2.1. Participants
2.2. Procedure
2.3. Behavioural Coding of Mutual Gaze
2.4. fNIRS Data Preprocessing
fNIRS Signals
2.5. Analyses
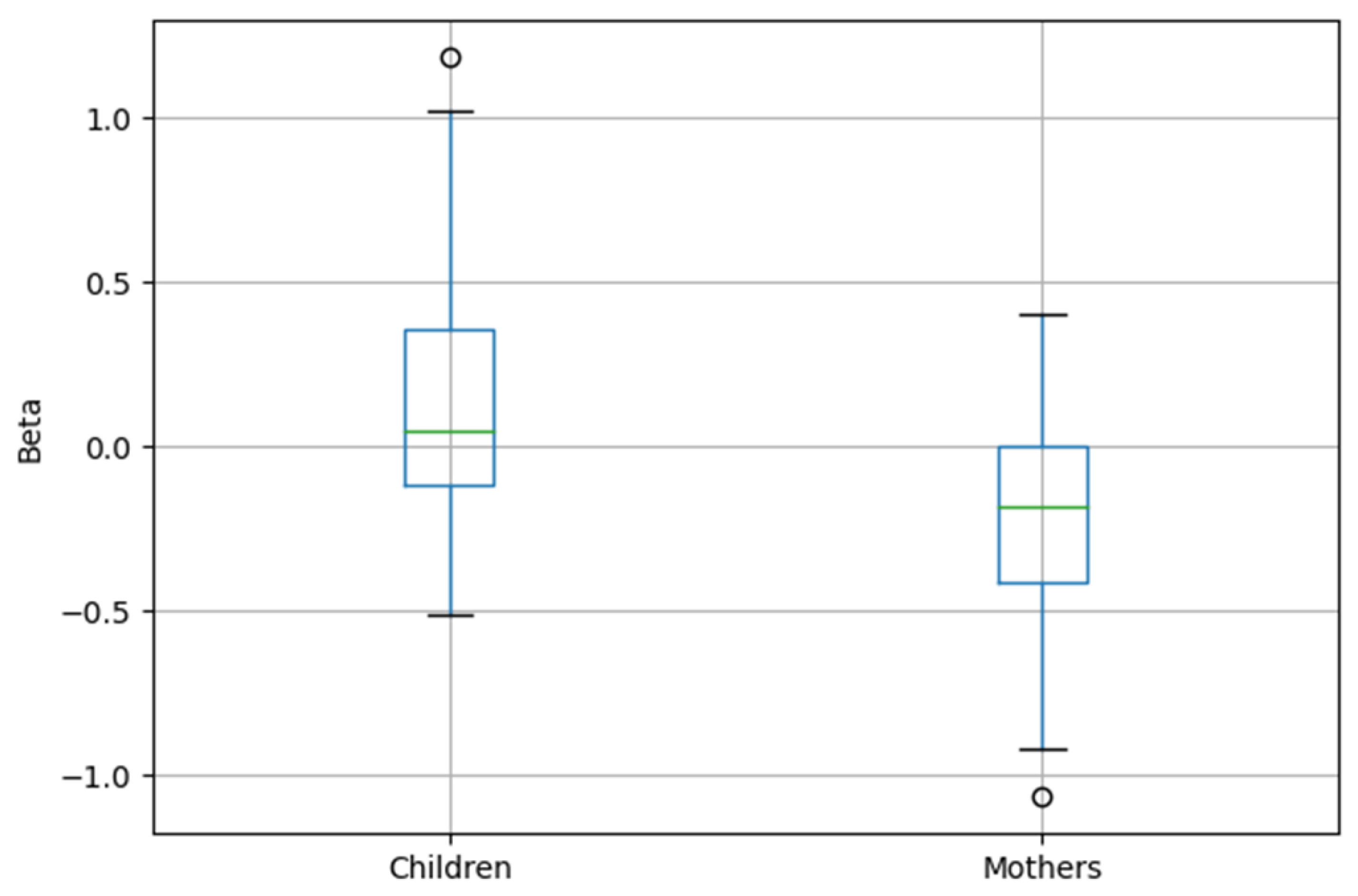
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atzil, S.; Hendler, T.; Feldman, R. The brain basis of social synchrony. Soc. Cogn. Affect. Neurosci. 2014, 9, 1193–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atzil, S.; Gendron, M. Bio-behavioral synchrony promotes the development of conceptualized emotions. Curr. Opin. Psychol. 2017, 17, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, H.N.; Menna, R. Physical and relational aggression in young children: The role of mother–child interactional synchrony. Early Child Dev. Care 2013, 183, 207–222. [Google Scholar] [CrossRef]
- Habersaat, S.; Monnier, M.; Peter, C.; Bolomey, L.; Borghini, A.; Despars, J.; Pierrehumbert, B.; Müller-Nix, C.; Ansermet, F.; Hohlfeld, J. Early mother-child interaction and later quality of attachment in infants with an orofacial cleft compared to infants without cleft. Cleft-Palate-Craniofacial J. 2013, 50, 704–712. [Google Scholar] [CrossRef]
- De Falco, S.; Emer, A.; Martini, L.; Rigo, P.; Pruner, S.; Venuti, P. Predictors of mother–child interaction quality and child attachment security in at-risk families. Front. Psychol. 2014, 5, 898. [Google Scholar] [CrossRef] [Green Version]
- Bornstein, M.H. Handbook of Parenting: Volume 4 Social Conditions and Applied Parenting; Psychology Press: London, UK, 2005. [Google Scholar]
- Bornstein, M.H.; Suwalsky, J.T.D.; Putnick, D.L.; Gini, M.; Venuti, P.; de Falco, S.; Heslington, M.; de Galperín, C.Z. Developmental Continuity and Stability of Emotional Availability in the Family: Two Ages and Two Genders in Child-Mother Dyads from Two Regions in Three Countries. Int. J. Behav. Dev. 2010, 34, 385–397. [Google Scholar] [CrossRef]
- Garralda, E.; Raynaud, J.P. (Eds.) Brain, Mind, and Developmental Psychopathology in Childhood; Jason Aronson, Inc.: Lanham, MD, USA, 2012; pp. 3–30. [Google Scholar]
- Dexter, C.A.; Wong, K.; Stacks, A.M.; Beeghly, M.; Barnett, D. Parenting and attachment among low-income African American and Caucasian preschoolers. J. Fam. Psychol. 2013, 27, 629. [Google Scholar] [CrossRef] [Green Version]
- Koike, T.; Tanabe, H.C.; Sadato, N. Hyperscanning neuroimaging technique to reveal the “two-in-one” system in social interactions. Neurosci. Res. 2015, 90, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Legerstee, M.; Haley, D.W.; Bornstein, M.H. (Eds.) The Infant Mind: Origins of the Social Brain; Guilford Press: New York, NY, USA, 2013. [Google Scholar]
- Feldman, R. Interactive synchrony: A biobehavioral model of mutual influences in the formation of affiliative bonds in healthy and pathological development. Neuropsychiatr. L’Enfance L’Adolescence 2012, 5, S2. [Google Scholar] [CrossRef]
- Bell, M.A. Mother-child behavioral and physiological synchrony. Adv. Child Dev. Behav. 2020, 58, 163–188. [Google Scholar]
- Gardner, F. Methodological issues in the direct observation of parent–child interaction: Do observational findings reflect the natural behavior of participants? Clin. Child Fam. Psychol. Rev. 2000, 3, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Venuti, P. Symmetry in Infancy: Analysis of Motor Development in Autism Spectrum Disorders. Symmetry 2009, 1, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Carollo, A.; Lim, M.; Aryadoust, V.; Esposito, G. Interpersonal synchrony in the context of caregiver-child interactions: A document co-citation analysis. Front. Psychol. 2021, 12, 701824. [Google Scholar] [CrossRef]
- Azhari, A.; Leck, W.Q.; Gabrieli, G.; Bizzego, A.; Rigo, P.; Setoh, P.; Bornstein, M.H.; Esposito, G. Parenting stress undermines mother-child brain-to-brain synchrony: A hyperscanning study. Sci. Rep. 2019, 9, 11407. [Google Scholar] [CrossRef] [Green Version]
- Leong, V.; Byrne, E.; Clackson, K.; Georgieva, S.; Lam, S.; Wass, S. Speaker gaze increases information coupling between infant and adult brains. Proc. Natl. Acad. Sci. USA 2017, 114, 13290–13295. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.; Schleihauf, H.; Kayhan, E.; Matthes, D.; Vrtička, P.; Hoehl, S. The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex 2020, 124, 235–249. [Google Scholar] [CrossRef]
- Piazza, E.A.; Hasenfratz, L.; Hasson, U.; Lew-Williams, C. Infant and Adult Brains Are Coupled to the Dynamics of Natural Communication. Psychol. Sci. 2020, 31, 6–17. [Google Scholar] [CrossRef]
- Astolfi, L.; Toppi, J.; Fallani, F.D.V.; Vecchiato, G.; Cincotti, F.; Wilke, C.T.; Yuan, H.; Mattia, D.; Salinari, S.; He, B.; et al. Imaging the social brain by simultaneous hyperscanning during subject interaction. IEEE Intell. Syst. 2011, 26, 38. [Google Scholar] [CrossRef] [Green Version]
- Koike, T.; Sumiya, M.; Nakagawa, E.; Okazaki, S.; Sadato, N. What makes eye contact special? Neural substrates of on-line mutual eye-gaze: A hyperscanning fMRI study. Eneuro 2019, 6. [Google Scholar] [CrossRef]
- Gergely, G.; Watson, J.S. Early socio-emotional development: Contingency perception and the social-biofeedback model. In Early Social Cognition: Understanding Others in the First Months of Life; Rochat, P., Ed.; Lawrence Erlbaum Associates Publishers: Mahwah, NI, USA, 1999; pp. 101–136. [Google Scholar]
- Liu, N.; Mok, C.; Witt, E.E.; Pradhan, A.H.; Chen, J.E.; Reiss, A.L. NIRS-Based Hyperscanning Reveals Inter-brain Neural Synchronization during Cooperative Jenga Game with Face-to-Face Communication. Front. Hum. Neurosci. 2016, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Reindl, V.; Gerloff, C.; Scharke, W.; Konrad, K. Brain-to-brain synchrony in parent-child dyads and the relationship with emotion regulation revealed by fNIRS-based hyperscanning. NeuroImage 2018, 178, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Azhari, A.; Gabrieli, G.; Bizzego, A.; Bornstein, M.H.; Esposito, G. Probing the association between maternal anxious attachment style and mother-child brain-to-brain coupling during passive co-viewing of visual stimuli. Attach. Hum. Dev. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Azhari, A.; Bizzego, A.; Esposito, G. Father-child dyads exhibit unique inter-subject synchronisation during co-viewing of animation video stimuli. Soc. Neurosci. 2021, 16, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Durnford, J.R.; Balagtas, J.P.M.; Azhari, A.; Lim, M.; Gabrieli, G.; Bizzego, A.; Esposito, G. Presence of parent, gender and emotional valence influences preschoolers’ PFC processing of video stimuli. Early Child Dev. Care 2020, 1–11. [Google Scholar] [CrossRef]
- Azhari, A.; Wong, A.W.T.; Lim, M.; Balagtas, J.P.M.; Gabrieli, G.; Setoh, P.; Esposito, G. Parents’ past bonding experience with their parents interacts with current parenting stress to influence the quality of interaction with their child. Behav. Sci. 2020, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Bizzego, A.; Gabrieli, G.; Azhari, A.; Setoh, P.; Esposito, G. Computational methods for the assessment of empathic synchrony. In Progresses in Artificial Intelligence and Neural Systems; Springer: Berlin/Heidelberg, Germany, 2021; pp. 555–564. [Google Scholar]
- Bornstein, M.H.; Maurice Haynes, O.; O’Reilly, A.W.; Painter, K.M. Solitary and Collaborative Pretense Play in Early Childhood: Sources of Individual Variation in the Development of Representational Competence. Child Dev. 1996, 67, 2910. [Google Scholar] [CrossRef] [PubMed]
- Azhari, A.; Bizzego, A.; Esposito, G. Parent-Child Dyads with Greater Parenting Stress Exhibit Enhanced Inter-brain Synchrony During Shared Play. ResearchSquare 2022. [Google Scholar] [CrossRef]
- Gamer, M.; Lemon, J.; Fellows, I.; Singh, P. Solomon Coder (Version Beta: 22 March 2017): A Simple Solution for Behaviour Coding. Available online: https://solomon.andraspeter.com (accessed on 22 March 2017).
- Gamer, M.; Lemon, J.; Fellows, I.; Singh, P. Various Coefficients of Inter-rater Reliability and Agreement. Available online: https://cran.r-project.org/web/packages/irr/irr.pdf (accessed on 28 September 2012).
- Quaresima, V.; Ferrari, M. Functional Near-Infrared Spectroscopy (fNIRS) for Assessing Cerebral Cortex Function During Human Behavior in Natural/Social Situations: A Concise Review. Organ. Res. Methods 2019, 22, 46–68. [Google Scholar] [CrossRef]
- Morais, G.A.Z.; Scholkmann, F.; Balardin, J.B.; Furucho, R.A.; de Paula, R.C.V.; Biazoli, C.E.; Sato, J.R. Non-neuronal evoked and spontaneous hemodynamic changes in the anterior temporal region of the human head may lead to misinterpretations of functional near-infrared spectroscopy signals. Neurophotonics 2017, 5, 011002. [Google Scholar]
- Pollonini, L.; Olds, C.; Abaya, H.; Bortfeld, H.; Beauchamp, M.S.; Oghalai, J.S. Auditory cortex activation to natural speech and simulated cochlear implant speech measured with functional near-infrared spectroscopy. Hear. Res. 2014, 309, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, D280–D298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholkmann, F.; Wolf, M. General equation for the differential pathlength factor of the frontal human head depending on wavelength and age. J. Biomed. Opt. 2013, 18, 105004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, J.; Zhang, X.; Noah, J.A.; Ono, Y. Frontal temporal and parietal systems synchronize within and across brains during live eye-to-eye contact. Neuroimage 2017, 157, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Koike, T.; Tanabe, H.C.; Okazaki, S.; Nakagawa, E.; Sasaki, A.T.; Shimada, K.; Sugawara, S.K.; Takahashi, H.K.; Yoshihara, K.; Bosch-Bayard, J.; et al. Neural substrates of shared attention as social memory: A hyperscanning functional magnetic resonance imaging study. Neuroimage 2016, 125, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachat, F.; Hugueville, L.; Lemaréchal, J.D.; Conty, L.; George, N. Oscillatory Brain Correlates of Live Joint Attention: A Dual-EEG Study. Front. Hum. Neurosci. 2012, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Wass, S.V.; Clackson, K.; Georgieva, S.D.; Brightman, L.; Nutbrown, R.; Leong, V. Infants’ visual sustained attention is higher during joint play than solo play: Is this due to increased endogenous attention control or exogenous stimulus capture? Dev. Sci. 2018, 21, e12667. [Google Scholar] [CrossRef] [Green Version]
- Kuboshita, R.; Fujisawa, T.X.; Makita, K.; Kasaba, R.; Okazawa, H.; Tomoda, A. Intrinsic brain activity associated with eye gaze during mother-child interaction. Sci. Rep. 2020, 10, 18903. [Google Scholar] [CrossRef]
- Farran, D.C.; Hirschbiel, P.; Jay, S. Toward Interactive Synchrony: The Gaze Patterns of Mothers and Children in Three Age Groups. Int. J. Behav. Dev. 1980, 3, 215–224. [Google Scholar] [CrossRef]
- Messer, D.J.; Vietze, P.M. Timing and transitions in mother-infant gaze. Infant Behav. Dev. 1984, 7, 167–181. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Hiraki, K. Prefrontal cortex and executive function in young children: A review of NIRS studies. Front. Hum. Neurosci. 2013, 7, 867. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, E.; Chisum, H.J.; Townsend, J.; Cowles, A.; Covington, J.; Egaas, B.; Harwood, M.; Hinds, S.; Press, G.A. Normal Brain Development and Aging: Quantitative Analysis at in Vivo MR Imaging in Healthy Volunteers. Radiology 2000, 216, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Knickmeyer, R.C.; Gouttard, S.; Kang, C.; Evans, D.; Wilber, K.; Smith, J.K.; Hamer, R.M.; Lin, W.; Gerig, G.; Gilmore, J.H. A Structural MRI Study of Human Brain Development from Birth to 2 Years. J. Neurosci. 2008, 28, 12176–12182. [Google Scholar] [CrossRef] [PubMed]
- Bunge, S.A.; Dudukovic, N.M.; Thomason, M.E.; Vaidya, C.J.; Gabrieli, J.D.E. Immature Frontal Lobe Contributions to Cognitive Control in Children. Neuron 2002, 33, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Durston, S.; Thomas, K.M.; Yang, Y.; Ulug, A.M.; Zimmerman, R.D.; Casey, B.J. A neural basis for the development of inhibitory control. Dev. Sci. 2002, 5, F9–F16. [Google Scholar] [CrossRef]
- Feldman, R. Social Behavior as a Transdiagnostic Marker of Resilience. Annu. Rev. Clin. Psychol. 2021, 17, 153–180. [Google Scholar] [CrossRef]
- Helminen, T.M.; Kaasinen, S.M.; Hietanen, J.K. Eye contact and arousal: The effects of stimulus duration. Biol. Psychol. 2011, 88, 124–130. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Coefficient | t | p-Value | |
|---|---|---|---|
| Intercept | 0.21 | 1.989 | 0.049 |
| Mother | −0.39 | −3.827 | <0.001 |
| Frontal Right | −0.1 | −0.788 | 0.432 |
| Posterior Left | −0.17 | −1.128 | 0.262 |
| Posterior Right | 0.02 | 0.14 | 0.889 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azhari, A.; Bizzego, A.; Balagtas, J.P.M.; Leng, K.S.H.; Esposito, G. Asymmetric Prefrontal Cortex Activation Associated with Mutual Gaze of Mothers and Children during Shared Play. Symmetry 2022, 14, 998. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14050998
Azhari A, Bizzego A, Balagtas JPM, Leng KSH, Esposito G. Asymmetric Prefrontal Cortex Activation Associated with Mutual Gaze of Mothers and Children during Shared Play. Symmetry. 2022; 14(5):998. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14050998
Chicago/Turabian StyleAzhari, Atiqah, Andrea Bizzego, Jan Paolo Macapinlac Balagtas, Kelly Sng Hwee Leng, and Gianluca Esposito. 2022. "Asymmetric Prefrontal Cortex Activation Associated with Mutual Gaze of Mothers and Children during Shared Play" Symmetry 14, no. 5: 998. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14050998