Possible Physical Mechanisms in the Galaxy to Cause Homochiral Biomaterials for Life

Astrophysics Division, UCLA Physics & Astronomy Department, 405 Hilgard Avenue, Los Angeles, California 90095, USA
Symmetry 2010, 2(3), 1450-1460; https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031450
Submission received: 2 February 2010
/
Revised: 8 May 2010
/
Accepted: 21 June 2010
/
Published: 9 July 2010
(This article belongs to the Special Issue Symmetry of Life and Homochirality)
Abstract
:The origin of homochirality in life remains a mystery that some believe is essential for life, and which may result from chiral symmetry breaking interactions with galactic organic material.
1. Introduction
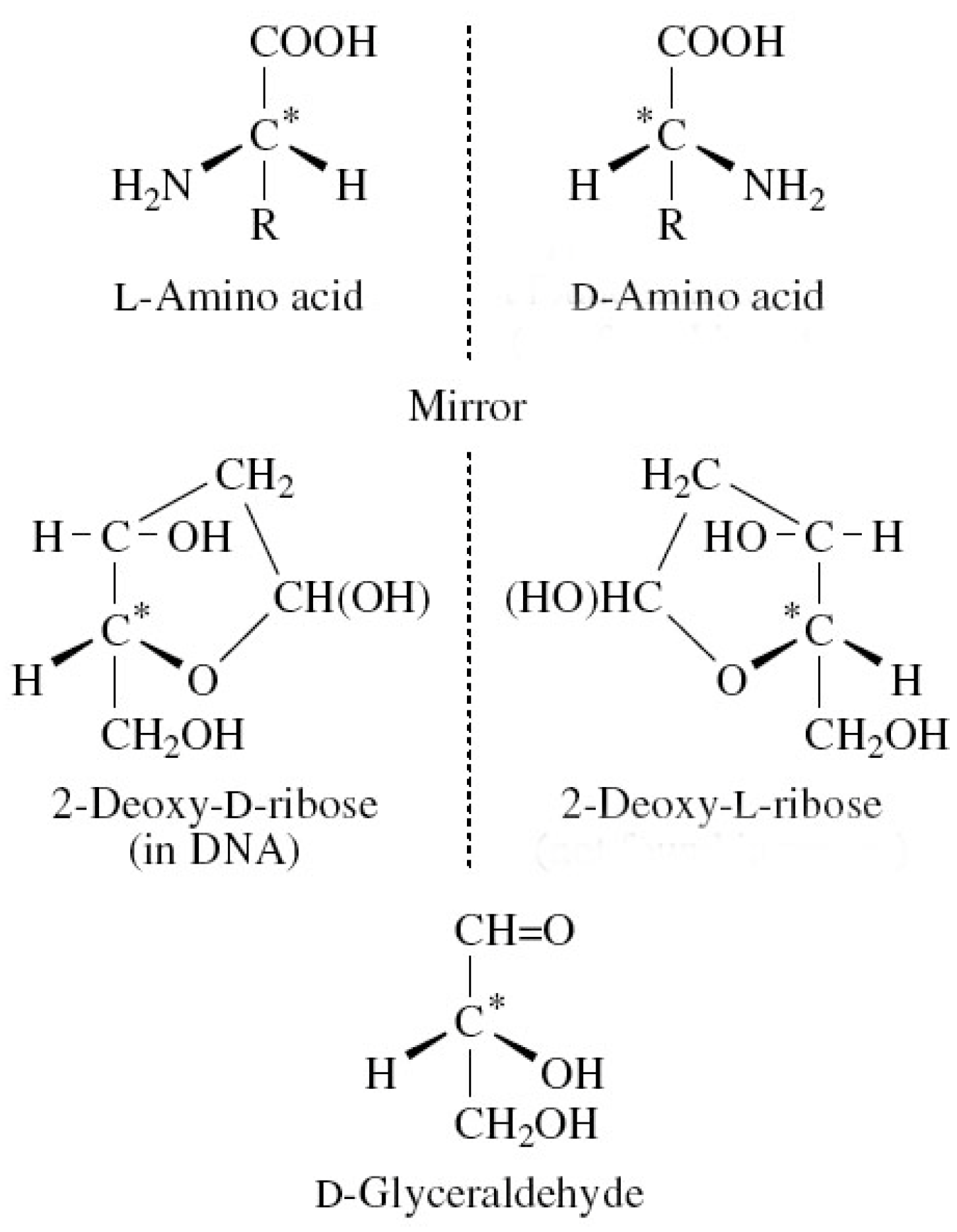
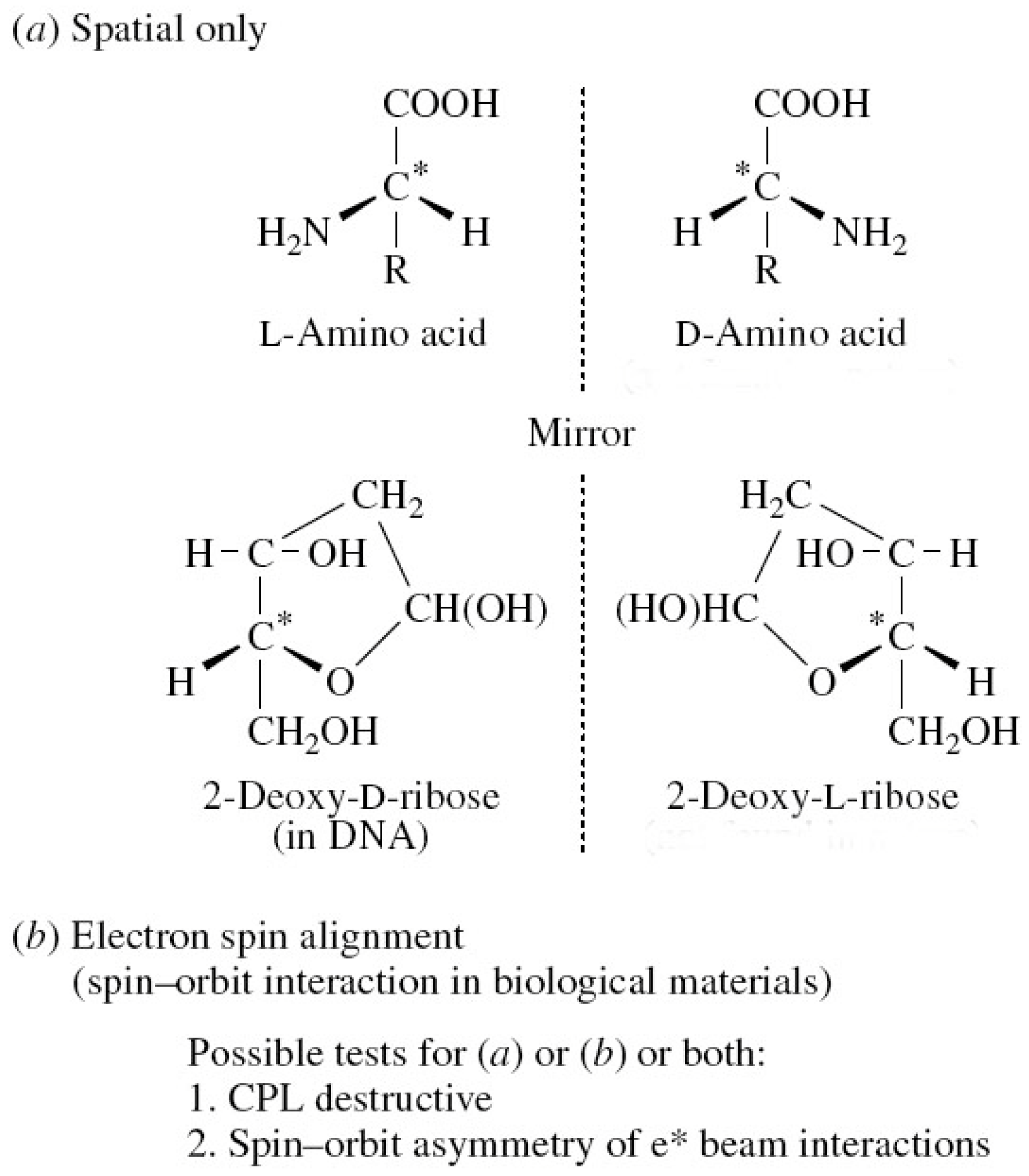
For more than a century, there has been evidence for the chiral nature of life forms on Earth. Pasteur was among the first to point this out (1848–1880), and the universal nature of chiral symmetry breaking in DNA and RNA is now very well established for all life forms. Figure 1 and Figure 2 show how the homochirality is manifest at the molecular level. We note that amino acids are L and DNA is largely D handed.
With the discovery of parity violation within charged current reactions in 1956, and of the weak neutral currents (WNCs) in 1973, two universal symmetry-breaking processes (WNC and β-decay) were uncovered that could have determined the handedness of DNA and RNA. The main problem is the extremely small symmetry-breaking effects (ΔE/kB T ~ 10-17). However there are plausible non-linear mechanisms that could have amplified this small, symmetry-breaking phase transition up to the full symmetry-breaking level observed in life forms [1,2,3,4,5,6,7,8,9,10,11].
For many years, there have been several issues associated with the homochiral structure of biomolecules, as first observed by Pasteur in 1848:
- (a)
- Is a homochiral structure necessary for life as we know it?
- (b)
- Did homochirality precede the formation of life (homochiral prebiotic medium hypothesis)?
- (c)
- Is there any reasonable physical mechanism that could have produced the large chiral symmetry breaking in the prebiotic medium or in the observed homochiral structure?
- (d)
- Is the homochiral structure an accident that occurred in biological systems, which was later amplified?
- (e)
- Can the homochirality be used as a signature for existing, or previous, living systems in the solar system or other parts of our galaxy?
- (f)
- Are there any experiments that can be carried out now to clarify the origin of homochirality?
- (g)
Recently, there has been increasing interest in the chiral nature or handedness of biomolecules. In fact, there are some who claim that the complex biomolecular structure of life must have arisen from a “chiral pure” medium [12,13]. This may be a precondition for the emergence of self-replicating biomolecular systems. Table 1 lists some of these ideas, which have been put forward largely by W. Bonner and V. Goldanskii [12,13]. We also point out a recent review in Reference [14]; see also Reference [15] for more recent details. Life on Earth is likely to have originated between 3.8 and 3.5 billion years ago. This estimate of time, along with the previous concept of the prebiotic medium, leaves a small window of 300 million years or less for life to have emerged from that prebiotic medium. Indeed, some speculate that the time could be less than 10 million years. Recently, some evidence has been obtained for an excess of L amino acids in the Murchison meteorite [37]. This and other observations may point to an extraterrestrial origin of homochirality [37].
We now turn to the experiments that study possible asymmetry processes. Over a period of 20 years, many experiments have been carried out (see Reference [12] for a nearly complete list). However, it appears that nearly every positive effect that was observed has turned out to be incorrect. In Table 2, we list some experimental results that are not yet refuted or are in direct conflict with previous null effects. The observation that circularly polarized light destroys L and D isomers selectively (entry 1 in Table 2) is now well established.
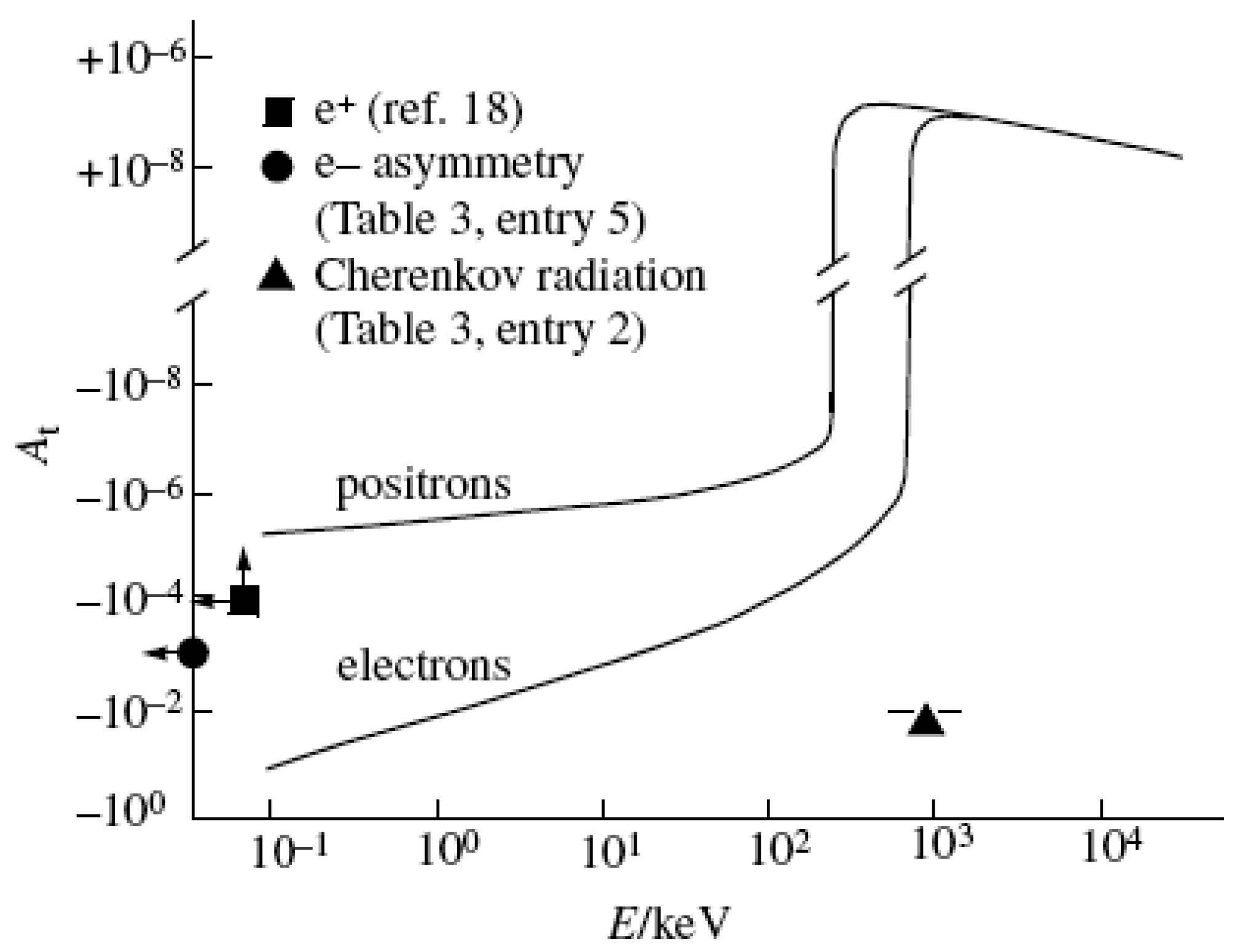
The other experiment to which we will refer is that of entry 2 (also in Table 2), the study of Cherenkov light in L or D material with chiral e- (β-decay) [20]. The authors claim an effect of perhaps 10-2 magnitude. This is, in our belief, far too large an effect, but a future study of Cherenkov radiation from L or D materials with polarized e- beams could be promising.
In Figure 3, we show the current limits or observations of asymmetry. The e+ measurement of Gridley et al. [18] is a very nice experiment. See also references [16] and [17]. Our conclusion is that no present experiment has reached the level of sensitivity needed to observe an effect. Therefore, it is premature to count out the weak force as a determining factor in the origin of homochirality in life.
2. Organic Molecules in Space and the Possible Role of Nearby Supernovae and Neutron Stars
One of the main themes of recent attempts to understand life on Earth is the likelihood that most of the early organic material on Earth was brought in by comets and asteroids. Reference [15] gives a nice introduction, from different points of view, to this concept. There are some interesting “large numbers” to consider in this regard:
- (1)
- The estimated amount of dust matter in the galaxy is ~10-4 mG, or ≤ 107 solar masses, largely in the form of dust grains. A fraction of that material is in the form of organic materials [26].
- (2)
- (3)
- In a molecular cloud with a density of 104 M/cm3 and a radius of 1 parsec, there could be a complex of organic matter equal to 100 solar masses.
- (4)
- The Earth revolves around the galaxy every ~250 million years, and it likely encounters several dense layers of molecular clouds in this trajectory.
- (5)
- It is likely that large quantities of organic material were deposited in the Earth in the first billion years.
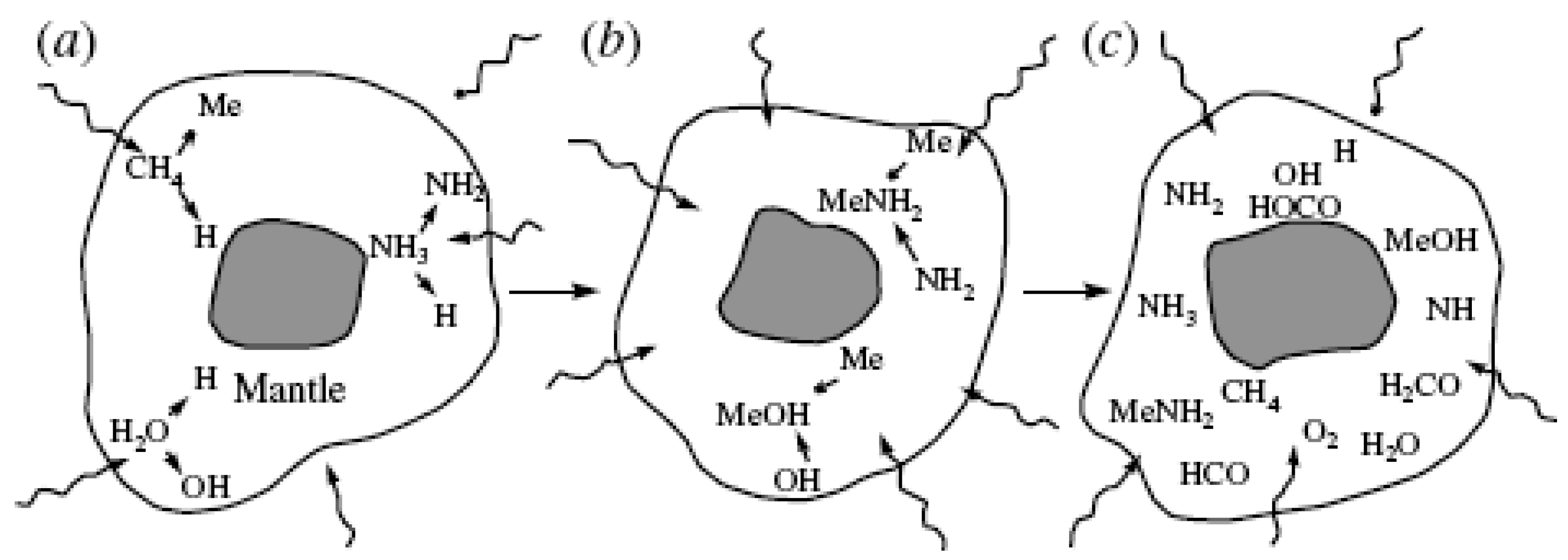
The above information is obtained by observing the infrared scattering of dust in the galaxy and by modeling various UV-driven processes here on Earth [26]. Ultraviolet photo processing plays an important role in the organic chemistry of the dust particles [27]; see also Reference [28]. Figure 4 shows the nature of a dust grain with prebiotic molecules inside [29].
We discuss here two scenarios where chiral interactions in the ISM could have led to a preponderance of homochiral molecules. These two concepts are outlined in Table 3, with Figure 5 giving a fairly complete description of the hypothesis. The second hypothesis is illustrated in Figure 6, where the possibility of the relative survival of the e± polarization deep within the cloud is also illustrated. Of course, during this time, the effect of the WNC can be driving the system towards a homochiral state.
Let us consider the rate of these three effects:
- (1)
- For absorption and a supernova 1 parsec away (or inside a 1-parsec dense cloud), the number of interactions will be ~10-3 /kg of material for 100 MO of organic material (which would be 1012 g of organic matter that is active), therefore the positron from the interactions would lose energy at a rate of 10-19 MeV/cm, and thus travel over a parsec. ( is an antielectron neutrino).
- (2)
- For the coherent vx + N → vx + N, and for the carbon in the hydrocarbons, we would have ~102 more or ~1014 grams of active material. Note that νx stands for all types of neutrinos. This effect could be very important in light of the small energy difference that separates L and D molecules, and the possibility of large coherent effects.
- (3)
- For the Al26 over the half-life, there would be ~1050 decays producing ~1050 positrons that lose energy at the rate of 10-19 MeV/cm; for MeV positrons, the range would be on the order of a parsec (ignoring possible magnetic-field effects).
2.1. Direct Interaction of the Supernova II Neutrino
Consider the example where 0.001 MO of Al26 is produced and assume (MO is the mass of the sun), for simplicity, that the energy of the e+ is 1 MeV and is contained in the gas cloud. Assume that the cloud has a density of 104 atoms/cm3 and that 10-3 of the atoms are organic. The stopping power for e+ would then be
and for a density of ρ = 104 atoms/cm3 ~ 10-17 g/cm3, we find
and for an average energy exchange of 10 eV, we have
For 0.001 MO of Al26 and a 10-3 organic fraction, we obtain a total of ~1050 collisions of polarized positrons, with organic materials in the cloud, assuming all of the e+ stop in the cloud (we assume that only one of the collisions can result in spin exchange). There will also be the same order of polarized photons from the e+e- → γγ annihilation. It is estimated in [17] and [24] that the asymmetry due to the weak interaction would be of order 10-11 to 10-6, depending on the positron energy {it scales like α2[α/(ν/c)]2}. Thus, it takes N ~1022 interactions for the asymmetric to become statistically important. In this example there are far more interactions.
More recently we have considered the main effects of a SN II explosion in a hydrocarbon cloud. We estimate that the larger anti-neutrino interaction rate would cause a chirality breaking effect and estimate this effect in Table 4. See Reference [34].
It appears that supernova neutrinos could include a chiral symmetry breaking in the ISM that could be transferred to the biomolecules of life. I wish to thank Goldanskii for some very useful conversations while he was a Regents Lecturer at UCLA and I regret his passing; please see Reference [5].
3. Other Calculations of Chiral Symmetry Breaking
The process shown in Figure 6 could come about in similar ways. Here are two examples:
- (a)
- Effects of cosmological neutrinos on the discrimination between the two enantiomers of a chiral molecule [35]. This concept is similar in spirit to that of Cline and the authors reference that work [34]. The basic concept in this work is that cosmological neutrinos and antineutrinos interact with the electrons in organic materials in the galaxy. This results in a split in the energy of the different chiral systems. This energy split is enhanced by the contributions if all the electrons in the molecules, and other mechanisms, up to the point of a larger chiral symmetry breaking.
- (b)
- A relativistic neutron fireball from a supernova explosion is a possible source of chiral influence [36]. This concept is also similar to the one by Cline [34]. However, in this case, it is the decay of neutrons that leads to polarized electrons that destroy organic material differently for L and D enantiomers. This is shown in Figure 3 and explained in the text. This work studies the effect of neutrinos from the supernova fireball with a Lorentz factor of one hundred. The relativistic electron-proton plasma (from the neutron decays) is slowed down by collective effect. There is high chiral efficiency for such electrons. The electron interactions and the photons produced both help destroy one chiral state, leading to the dominance of the other. This idea is very similar to that of Cline [34].
4. Summary of Concepts of the Production of Homochiral Molecules in the Galaxy
References and Notes
- Kondepudi, D.D.; Nelson, G.W. Weak neutral currents and the origin of biomolecular chirality. Nature 1985, 314, 438–441. [Google Scholar] [CrossRef]
- Zel’dovich, Y.B.; Mikhailov, A.S. Fluctuation kinetics of reactions. Sov. Phys. Usp. 1987, 30, 977–992. [Google Scholar] [CrossRef]
- Avetisov, V.A.; Goldanskii, E.I.; Kuz’min, V.V. Handedness, Origin of life and evolution. Phys. Today 1991, 44, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Salam, A. The role of chirality in the origin of life. J. Mol. Evol. 1991, 33, 105–113. [Google Scholar] [CrossRef]
- Avetisov, V.; Goldanskii, V. Mirror symmetry breaking at the molecular level. Proc. Natl. Acad. Sci. USA 1996, 93, 11435–11442. [Google Scholar] [CrossRef] [PubMed]
- Hegstrom, R.Q.; Rein, D.W.; Sandars, P.G.H. Calculation of the parity nonconserving energy difference between mirror-image molecules. J. Chem. Phys. 1980, 73, 2329–2341. [Google Scholar] [CrossRef]
- Mason, S.F.; Tranter, G.E. The electroweak origin of biomolecular handedness. Proc. R. Soc. London 1985, A397, 45–65. [Google Scholar] [CrossRef]
- Cline, D.B.; Liu, Y.; Wang, H. Simulation of a weak interaction induced chiral transition in a pre-biotic medium. In 30 Years of Weak Neutral Currents; Proceedings of the International Symposium on the Discovery of Weak Neutral Currents: The Weak Interaction Before and After, Santa Monica, Santa Monica, CA, USA, 3–5 February 1993; Mann, A.K., Cline, D.B., Eds.; AIP Conference Proceedings 300: New York, NY, USA, 1994; pp. 499–505. [Google Scholar]
- Ulbricht, T.L.V. Asymmetry: The non-conservation of parity and optical activity. Quart. Rev. 1959, 13, 48–60. [Google Scholar] [CrossRef]
- Vester, F.; Ulbricht, T.L.V.; Krauch, H. Optical activity and parity violation in β-decay. Naturwissenschaften 1958, 40, 68. [Google Scholar]
- Yamagata, Y. A hypothesis for the asymmetric appearance of biomolecules on Earth. J. Theor. Biol. 1966, 11, 495–498. [Google Scholar] [CrossRef]
- Bonner, W. The origin and amplification of biomolecular chirality. Origins Life Evol. B. 1991, 21, 59–111. [Google Scholar] [CrossRef]
- Goldanskii, V.I.; Kuz’min, V.V. Spontaneous Breaking of mirror symmetry in nature and the origins of life. Sov. Phys. Usp. 1989, 32, 1–29. [Google Scholar] [CrossRef]
- Keszthely, L. Origin of the homochirality of biomolecules. In Quarterly Rev. Biophys.; Goldstein, S., Horwich, A., Leslie, A., Lilley, D., Norden, B., Wagner, G., Eds.; University of Chicago: Chicago, IL, USA, 1995. [Google Scholar]
- Cline, D.B. Proceedings of the 1st Symposium on the Physical Origins of Homochirality of Life, Santa Monica, CA, USA, February 1995; AIP Conference Proceedings 379: New York, NY, USA, 1995.
- Hegstrom, R.A. Weak neutral currents and β radiolysis effects on the origin of biomolecular chirality. Nature 1985, 315, 749–750. [Google Scholar] [CrossRef]
- Meiring, W.R. Nuclear β-decay and the origin of biomolecular chirality. Nature 1987, 329, 712–714. [Google Scholar] [CrossRef]
- Gidley, D.; Rich, A.; Vanhouse, J.; Zitzewitz, P.W. β-decay and the origins of biological chirality: Experimental results. Nature 1982, 297, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Norden, B. Was photoresolution of amino acids the origin of optical activity in life? Nature 1977, 266, 567–568. [Google Scholar] [CrossRef] [PubMed]
- Garay, A.; Ahigren-Beckendorf, J.A. Differential interaction of chiral β-particles with enantiomers. Nature 1990, 346, 451–453. [Google Scholar] [CrossRef]
- Akaboshi, M.; Kawai, K.; Maki, H.; Erlich, W.; Honda, Y. Selective decomposition of either enantiomer or aspartic acid irradiated with 60CO-γ-rays in the mixed aqueous solution with D- or L-alanine. Origins Life Evol. B. 1990, 20, 111–119. [Google Scholar] [CrossRef]
- Conte, E. Investigation on the chirality of electrons from 90Sr-90Y Beta-decay and their asymmetrical interactions with D- and L-Alanines. Nuovo. Cimento 1985, 44, 641–647. [Google Scholar]
- Campbell, D.M.; Farago, P.S. Spin-dependent electron scattering from optically active molecules. Nature 1985, 318, 52–53. [Google Scholar] [CrossRef]
- Cline, D.B.; Liu, Y.; Wang, H. Effect of a chiral impulse on the weak interaction induced handedness in a pre-biotic medium. Origins Life Evol. B. 1995, 25, 201–209. [Google Scholar] [CrossRef]
- Park, J.; Cheng, C.W.; Cline, D.; Liu, Y.; So, H.; Wang, H. Origins of homochirality: A simulation by electrical circuit. Proc. SPIE 1995, 2551, 40–52. [Google Scholar]
- Greenberg, J.M.; Mendoza-Gomez, C.X.; Pirronello, V. The Chemistry of Life’s Origins; Kluwer Academic Publishing: Dordrecht, The Netherlands, 1993; pp. 1–32. [Google Scholar]
- Khare, B.U.; Sagan, C. Experimental interstellar organic chemistry, preliminary findings. Molecules in the Galactic Environment; Proceedings of a Symposium Held at the University of Virginia, Green Bank, VA, USA, 4–7 November, 1971; Gordon, M.A., Lewis, E., Eds.; Wiley: New York, NY, USA, 1973. [Google Scholar]
- Chyba, C.; Sagan, C. Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the signs of life. Nature 1992, 355, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Whittet, D.; Chiar, J. Cosmic Evolution of the Biorganic Elements and Compounds. Astro. Rev. 1993, 5, 1. [Google Scholar]
- Cline, D.B. Homochiral prebiotic molecule formation in dense molecular clouds. UCLA Preprint No. PPH0072.
- Chela-Flores, J. The origin of chirality in protein amino acids. Chirality 1994, 6, 165–168. [Google Scholar] [CrossRef]
- Pospelov, M. On the Scattering of Polarized Particles by Chiral Molecules. In Proceedings of the 1st Symposium on the Physical Origins of Homochirality of Life, Santa Monica, CA, USA, February 1995; Cline, D.B., Ed.; AIP Conference Proceedings 379: New York, NY, USA, 1995. [Google Scholar]
- Thiemann, W.H.P.; Meierhenrich, U. ESA-rosetta Mission. Adv. Space Res. 2001, 27, 323. [Google Scholar] [CrossRef]
- Cline, D.B. Supernova antineutrino interactions cause chiral symmetry breaking and possibly homochiral biomaterials for life. Mendeleev Commun. 2004, 14, 301–304. [Google Scholar] [CrossRef]
- Bargueño, P.; Gonzalo, I. Effect of cosmological neutrinos on discrimination between the two enantiomers of a chiral molecule. Origins Life Evol. B. 2006, 36, 171–176. [Google Scholar] [CrossRef]
- Gusev, G.A.; Saito, T.; Tsarev, V.A.; Uryson, A.V. A relativistic neutron fireball from a supernova explosion as a possible source of chiral influence. Origins Life Evol. B. 2007, 37, 259–266. [Google Scholar] [CrossRef]
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.; Chrysostomou, A.; Hough, J.H.; Gledhill, T.M.; McCall, A.; Clark, S.; Ménard, F.; Tamura, M. Circular polarization in star-formation regions: Implications for biomolecular homochirality. Science 1998, 281, 672–674. [Google Scholar] [CrossRef] [PubMed]
- Root-Bernstein, R.S. Simultaneous origin of homochirality, the genetic code and its directionality. BioEssays 2007, 29, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Molecular handedness of life: significance of RNA amnioacylation. J. Biosci. 2009, 34, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Breslow, R.; Levine, M.; Cheng, Z.L. Imitating prebiotic homochirality on earth. Orig. Life Evol. Biosph. 2010, 40, 11–26. [Google Scholar] [CrossRef]
Figure 1.
Examples of biomolecules that are isomers; some of which exist in nature and some that do not.
Figure 1.
Examples of biomolecules that are isomers; some of which exist in nature and some that do not.

Figure 2.
Various components of symmetry breaking in molecules.

Figure 3.
Most recently published calculations of the expected asymmetry for the scattering of e* from chiral molecules. We also include some recent measurements of the limits on such an asymmetry.
Figure 3.
Most recently published calculations of the expected asymmetry for the scattering of e* from chiral molecules. We also include some recent measurements of the limits on such an asymmetry.

Figure 4.
The structure of grains, when they initially accrete, is inferred from laboratory simulations, in which mixtures of water, methane, ammonia, and other simple molecules are subjected to UV irradiation at 10 K. (a) Each grain begins as a silicate core that condensed in the atmosphere of a cool giant star. Around this core, a mantle of ice forms. Ultraviolet radiation breaks some of the mantle molecules into radicals or reactive molecular fragments. (b) The radicals can then recombine in new ways. (c) Over a longer period, the continued UV irradiation of the grain can give rise to ever more complex mixtures of molecules and radicals. Data from Reference 29, which was used in the talk of Greenburg M. at the Homochiralty Symp., Santa Monica, CA, USA, 1995.
Figure 4.
The structure of grains, when they initially accrete, is inferred from laboratory simulations, in which mixtures of water, methane, ammonia, and other simple molecules are subjected to UV irradiation at 10 K. (a) Each grain begins as a silicate core that condensed in the atmosphere of a cool giant star. Around this core, a mantle of ice forms. Ultraviolet radiation breaks some of the mantle molecules into radicals or reactive molecular fragments. (b) The radicals can then recombine in new ways. (c) Over a longer period, the continued UV irradiation of the grain can give rise to ever more complex mixtures of molecules and radicals. Data from Reference 29, which was used in the talk of Greenburg M. at the Homochiralty Symp., Santa Monica, CA, USA, 1995.

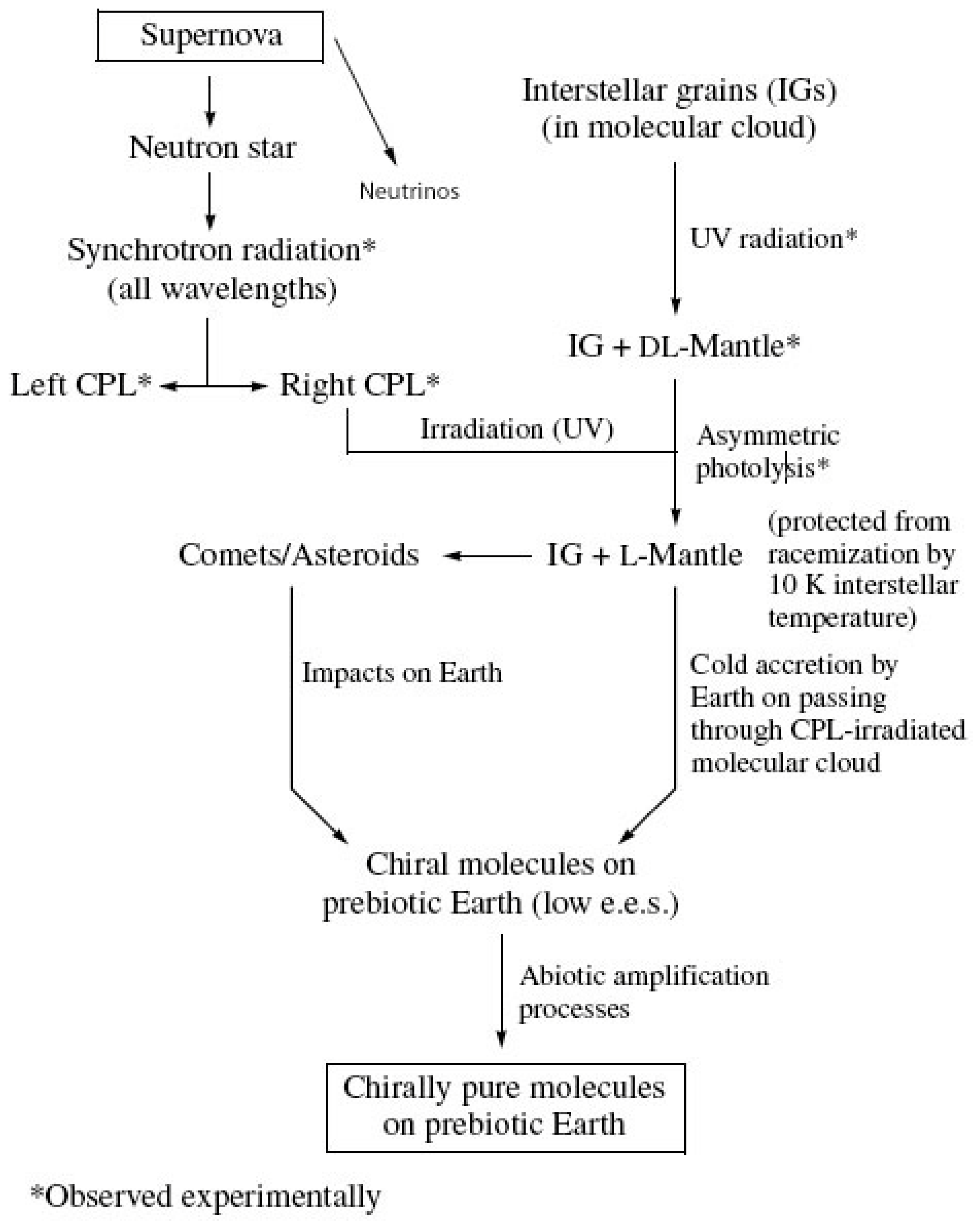
Figure 5.
Possible extraterrestrial origin of terrestrial homochiralty.

Figure 6.
(a) Schematic representation of the effect of a supernova II on the organic molecules in a dense molecular cloud. (b) Estimated polarization of the β* decay particles as the energy decreases, by ionization, in H2O and in the dense molecular cloud. An observation of polarized e+ is made by the Michigan group in Figure 6.
Figure 6.
(a) Schematic representation of the effect of a supernova II on the organic molecules in a dense molecular cloud. (b) Estimated polarization of the β* decay particles as the energy decreases, by ionization, in H2O and in the dense molecular cloud. An observation of polarized e+ is made by the Michigan group in Figure 6.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Trends in life origins.
| 1 | DNA: Self replication would not work with heterochiral systems (50% L and 50% D). |
| 2 | Errors in DNA Replication: Without a pure chiral structure, the error rate in replication would be unacceptable for long-lived systems (higher animal forms, trees, etc.). |
| 3 | In a prebiotic medium, homochirality must have been either (a) or (b) |
| (a) Established in a very short time on Earth (≤ 100 Million years) | |
| (b) Existed in Interstellar Medium (ISM) organic materials near the solar system |
Table 2.
Experiments where a chiral effect has been observed (and not refuted).
| Experiment | Beam/Source | Target | Detection | Comments | Reference | |
|---|---|---|---|---|---|---|
| 1 | CPL on L/D/ photo absorption | UV/keV radiation | DC tartaric acid, DL alanine, DL glutamic | Observe destructive difference in one chirality | To be expected from optical activity | Norden [19] |
| 2 | e- target → Č light | p32 source and Ca137 (no chiral electrons) | R- or S-PBA | Observe different Č light intensity due to chiral electrons | Effect too large to be due to Č radiation from spin effects | Garay et al. [20] |
| 3 | Co60 → γ + (L,D) | Co60 γs | D or L alanine | Observe different amounts of L,D after irradiation | Other experiments did not produce this effect | Akabosh et al. [21] |
| 4 | Introduction of L/D/ chiral particles | Sr90, Y90 β-decay | Y90 D or L alanine | Detects effects by electron spin resonance technique | The electron spin resonance may be sensitive to spin dependence | Conte et al. [22] |
| 5 | Low-energy polarized e- beam | GaAs polarized source 5 eV | Camphor L,D | Observe different electron polarization and beam attenuation in L/D | At such low energy, asymmetry may be too large | Campbell et al. [23] |
Table 3.
Possible sources of CPL (UV/keV) radiation in dense molecular clouds.
| Primordial Soup – Molecular Clouds (ISM) | |
|---|---|
| 1 | Synchronotron radiation from neutron stars ([12,26]) |
| CPL helicity depends on position (i.e., above or below star) | |
| In principle, this mechanism works. However, on the 250-Myr orbit around the galaxy, this effect is expected to average out. | |
| 2 | Radiation from weak interaction processes [30]-injection into molecular cloud |
| Processes: supernova II interaction, Al26 from nearby supernovas, etc.; | |
| Because of grain structure, dE/dx will be very different from that of solids, gases, or liquids; | |
| Always gives the same chiral symmetry breaking; | |
| An act as a chiral impulse along with WNC. |
Table 4.
Estimated rate of νe interactions in the dense presolar cloud.
| Assume: Cloud density 104 protons/cm3 [fraction of organic material] 10-3-- Range of e+ 1 parsec; Number of interactions ~ 1035 = Number of e+ for interaction with organics in cloud > 1030 We estimate that only 1022 interactions are necessary to produce asymmetry. From this, it is clear a much larger asymmetry is produced. |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Cline, D.B. Possible Physical Mechanisms in the Galaxy to Cause Homochiral Biomaterials for Life. Symmetry 2010, 2, 1450-1460. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031450
AMA Style
Cline DB. Possible Physical Mechanisms in the Galaxy to Cause Homochiral Biomaterials for Life. Symmetry. 2010; 2(3):1450-1460. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031450
Chicago/Turabian StyleCline, David B. 2010. "Possible Physical Mechanisms in the Galaxy to Cause Homochiral Biomaterials for Life" Symmetry 2, no. 3: 1450-1460. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031450