Symmetries of the Central Vestibular System: Forming Movements for Gravity and a Three-Dimensional World

Neuro-Otology Department, Legacy Research Center, 1225 NE 2nd Avenue, Portland, Oregon 97232, USA
*
Author to whom correspondence should be addressed.
Symmetry 2010, 2(3), 1544-1558; https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031544
Submission received: 18 May 2010
/
Accepted: 16 July 2010
/
Published: 22 July 2010
(This article belongs to the Special Issue Feature Papers: Symmetry Concepts and Applications)
Abstract
:Intrinsic dynamics of the central vestibular system (CVS) appear to be at least partly determined by the symmetries of its connections. The CVS contributes to whole-body functions such as upright balance and maintenance of gaze direction. These functions coordinate disparate senses (visual, inertial, somatosensory, auditory) and body movements (leg, trunk, head/neck, eye). They are also unified by geometric conditions. Symmetry groups have been found to structure experimentally-recorded pathways of the central vestibular system. When related to geometric conditions in three-dimensional physical space, these symmetry groups make sense as a logical foundation for sensorimotor coordination.
{kind=link}
{kind=link}
{kind=link}
1. Sensorimotor Integration and the Central Vestibular System
The experience of rocking in a rocking chair integrates information from several sensory receptors, as picking up a pen integrates the action of several fingers. Although one knows that the rocking chair rotates in a certain way, to experience the rocking motion involves neural activity flowing in sensory systems. The visual scene recedes as one rocks back and approaches as one rocks forward. This visual flow contributes to the experience of motion. Yet one can also feel the rocking motion with the eyes closed, as one can still pick up the pen with the forefinger lifted away from it. There remain other senses with which to experience the rocking motion. These senses include touch in the feet on the floor and in the skin next to the chair, musculoskeletal sensations in the feet if they are pushing or moving with the chair, and inner ear sensory receptors that register inertial stimuli. Several senses act together to grasp an experience of motion. Both the experience of motion and the movement of grasping with the hand are guided in their coordination by spatial geometry.
The commonality of spatial geometry to the various types of sensory information and body movements provides a framework for their integration in neural centers and pathways. For example, although optic flow and inertial head movement sensations are physiologically and anatomically distinct, each must conform to the three-dimensional motion dynamics that stimulate sensation. Head movements result in both optic flow and inertial sensation from the inner ear. Since optic flow and inertial sensation can provide similar motion information, they converge on many neurons—that is, they connect to many of the same neurons in such a way that their activity is combined [1,2]. However, the head movements can be passive or intentional and can arise from rotations about different body axes; to take these factors into account in action and perception, the central vestibular system (CVS) changes the origin of sensorimotor reference frames [3,4,5,6,7,8]. By actively and circumstantially combining the different sensory and motor information, the CVS facilitates the ability to grasp a motion experience with whatever sensory information is relevant and available. The CVS occupies a confluence of the spinal cord, direct neural connections from the inertial sensory receptors of the inner ear, and the accessory optic system, which carries optic flow [1,2,9,10], and it provides spatially-organized information to the hippocampus, thalamus, and cerebral cortex [11,12,13,14]. Spatially-related symmetry groups have been observed which organize the anatomy and physiology of the CVS [15,16,17].
Clinical observations provide some of the clearest evidence of the role of the CVS in balance and posture. Inertial information passes through and is organized in the CVS on its way to the spinal cord, in the vestibulospinal pathways. Patients with balance disorders display a wide range of balance deficits. Some have been shown to arise from a lack of inertial information and others from other dysfunctions of the peripheral sensory receptors of the inner ear. Some disorders are caused by dysfunction of the CVS itself. However, one of the most important points here is that, even lacking inertial information from the inner ear or having inaccurate information from the inner ear, the CVS and other sensorimotor integration centers can function normally or almost normally to organize the remaining sensorimotor information.
The organization of sensorimotor information depends on the ongoing activity. You coordinate the muscles and senses differently depending on whether you are rocking, running, hammering, or writing. Just as different movements are salient in each of these activities, so are different senses. Balance is more crucial to running than to rocking. Fine finger movements are more crucial to writing than to hammering. For each activity, we put ourselves into an appropriate state of sensory and motor coordination—a sensorimotor state—that prioritizes motor goals and coordinates the senses and many muscles around the activity. In all these cases, the sensorimotor system must satisfy geometric constraints in order to perform actions and to make sense of neural activity arising from sensory receptors. Even in the case of rocking in a rocking chair, a failure of sensorimotor coordination can lead to an acute sense of disorientation, for example, if a person misconstrues the position of the eyes or the neck.
The neck plays a crucial sensorimotor role, somewhat analogous to the eye muscles. In both cases, a sensory receptor is borne on a movable globe: the retina in the eye and the inertial endorgans in the head. In order to integrate visual information with movement, it is necessary to know where the eye is; in order to integrate inertial information with movement, it is necessary to know where the neck has turned the head. Visual information is doubly dependent on the neck, because the eye is in the head. For these reasons, knowledge of neck orientation is thought to be the crucial factor in motion sickness, a failure of sensorimotor integration in which the CVS plays a strong part [18,19].
Sensorimotor functions shade into cognition without a clear boundary. For example, the vestibular system is implicated in construing two-dimensional line drawings as depicting depth in three dimensions [20]. There is experimental and clinical evidence that the vestibular system has a substantive role in cognition [11]. Lakoff & Johnson [21] emphasize the role of spatial metaphor in language and cognition; such use of spatial metaphor may also depend on the physiological role of the CVS in sensorimotor spatial functions, besides hippocampal, thalamic, and cerebral cortex regions to which it connects [11,12,13,14]. Llinás [22] argues that nervous systems evolved for movement, so it would not be surprising to find the spatial and geometrical framework necessary for movement throughout neural functions. In this paper we address how spatial and geometrical frameworks are expressed in vestibular pathways and communicated to other neural centers.
2. Anatomy, Physiology, and Behavior
Anatomically and physiologically, the CVS is a set of nuclei and parts of the cerebellum, along with pathways connecting them together and to the rest of the nervous system [2]. (Physiology is anatomy when it is alive. The two are distinguished primarily by the experimental methods used to observe them.) Evolutionarily, the CVS is ancient, and the vestibulo-cerebellum is the oldest part of the cerebellum [23]. Vestibular pathways connect to neural centers essential to basic behavioral functions such as the spinal cord for body movements and the oculomotor centers for eye movements.
Studying anatomy and physiology can lead to an understanding of the way nervous systems design and initiate actions and construct perceptions and responses. What kind of causality operates in sensorimotor systems? For example, one billiard ball knocking another, hot air causing a balloon to rise, and radioactive decay are best described by differing causal logics: mechanics, thermodynamics, and quantum mechanics, respectively. This section presents three aspects of the causal logic of sensorimotor neurobiology—connection, construction, and modularity—and relates them to symmetry groups.
One aspect is that neural connections carry information essential for behavior. Lesions that disrupt connections prevent information from being communicated. The importance of connectivity has led the input-output relationships in particular experimental paradigms to be studied with great quantitative exactitude. Neural pathways typically consist of neuronal axons originating in one anatomically and physiologically distinct neural center or population and projecting onto one or more different neural centers, sometimes at great distances through the body. Translational symmetry groups of two-dimensional arrays of neurons illuminate the properties of these mappings; they have been used to understand properties of mappings in the visual system [24,25,26], and the entorhinal cortex of the hippocampus [27,28]. The "source" and "target" neuronal populations in these pathways do not consist of identical neurons. Rather, a population consists of neurons with similar morphology and activity patterns, but in which each neuron is an individual.
A second aspect of the relationship between anatomy/physiology and behavior is that neural systems have a certain amount of autonomy and display active behaviors. This autonomous aspect is necessary to construct a probable world, along with perceptions and actions to fit. Constructive perception is not an input-output process [29,30,31]. Rather, designing perceptions and actions to conform to three-dimensional spatial geometry may involve a template, and the spatially-related symmetry groups in vestibular pathways may serve as such guidelines (§4). Furthermore, symmetries of networks of differential equations are related to Hopf bifurcations [32] and, as a result, the symmetry group of an interconnected neural network governs the patterns of neural oscillation that will arise in that network. Although spontaneous oscillatory behavior is often associated with the properties of individual neurons in a population [33,34,35], it is not limited to that level of analysis. Given a pathway inducing a network of relationships on a target population, the symmetry group of the pathway can present a menu of geometrically-appropriate patterns for the nervous system to produce spontaneously and constructively [36]. Depending on the behavior of interacting networks, these patterns can be reflected in behaviors such as gaits and head movements. In the first case we describe below, the disynaptic projection from semicircular canal nerves to neck motor neurons (§4.2), the symmetries are related to sensory pathways; however, the oscillations in the network can arise independently of any particular sensory event.
A third aspect of the causal logic of nervous systems is the interaction of strongly interconnected but distinct anatomical/physiological and behavioral modules. While the word "modularity" is often associated with genetics, the fact of modularity is far more general in biology [37,38,39]. Behavioral modules include, for example, walking and grasping with the hand; these two can be performed separately or in various combinations. Basic anatomical units include the neuron and the organism itself. A neuron, in particular, is an almost-complete biological unit with autonomy near that of an organism; it should not be thought of as a passive element in an electrical circuit. Neurons participate, in turn, in anatomically and physiologically patterned modules, including neuronal populations and neural centers. Separate but interacting pathways with distinct symmetry groups present different menus of possible behaviors [32]. In designing and initiating actions and constructing perceptions and responses, nervous systems combine intricately interacting possibilities.
In Section 4, we present two distinct vestibular pathways, to the spinal cord and cerebellum, which have different symmetry groups and likely interact in many whole-body behaviors. Before presenting the exact symmetries of the pathways, we will describe the approximate symmetries of the sensory organs whose modalities are integrated in the CVS.
3. Multimodal Sensory Organization
We have seen in the previous sections that vestibular pathways communicate sensory information relevant to balance, gaze, and movement, organize that information for behavior, and play a role in the initiation of perception and action. The CVS integrates sensory modalities, including visual, somatosensory, and inertial, in the activity in its pathways. (Here "integration" means bringing together, in various ways, as the movements of the fingers are integrated in tying a knot.) Especially in rotational and visual pathways, information is often organized by the CVS according to three planes: one more or less horizontal when the head is held upright and two vertical, at right and left diagonals to the forward and right-left directions. This organization into three planes of rotation (or equivalently, three axes) facilitates the combination of multiple sensory modalities and the interweaving of sensory and motor functions. The three rotational planes form a basis for rotations in three-dimensional physical space.
Visual information in the accessory optic system is organized into these three planes [9,10]. When the eye rotates in one plane, the visual field flows in the opposite direction without deformation. It is only optic flow information that reaches the CVS, not visual patterns such as print.
We will review the anatomy of the inertial sensory receptor, because it is less familiar than that of the visual receptor, the eye. As the retina sits in the eye, the inertial receptors in the inner ear sit in a bony labyrinth, which is part of the skull. There are several "sensory epithelia" that sit like patches of moss on curving wet rock, in curves of this bony labyrinth [40]. Each sensory epithelium consists of cells arranged in two-dimensional layers, including a layer of cells with sensory hairs. The curve in which the "saccule"—one of the sensory epithelia—sits is more vertical, and the curve in which the "utricle" sits is more horizontal, so that the utricle is more involved in registering horizontal motions. The sensory hairs of the saccule and utricle are surrounded by a gel that is, in turn, weighted with lumps of calcium carbonate called otoliths. Together, the saccule and utricle are called the otolith organs. These otolith organs register gravitation and linear motions in the various directions.
As a later evolutionary development [41], the bony labyrinth in which the inertial receptors sit has become shaped so that fluid flows along three narrow, almost circular canals, like hula hoops. (For a graceful and thorough introduction to these endorgans and their evolution, see [42,43].) The fluid in these semicircular canals moves when the head rotates with a component in the plane of the hoop-like canal, deforming a diaphragm (called the cupula), which contains oriented sensory hair cells. In mammals, there are three of these canals on each side of the head, approximately in the three planes in which sensorimotor information is organized: one near horizontal and two vertical. The semicircular canals form a relatively straightforward biophysical system, registering only acceleration to a first and good approximation, with balanced inhibitory and excitatory responses on the right and left sides to interrelate the right and left sets of three canals each [42,43]. The near-orthogonality of the semicircular canals was a key to early understanding that the canals registered rotation in three spatial dimensions, thought of in terms of the pitch, roll, and yaw of a sailing ship and, later, an airplane [44]. Understanding of their sensory function then led to the realization that many central pathways are also organized according to the same three planes [42,43]. While it is tempting to think that the nervous system evolved in the order of human understanding --sensory endorgans first—it may not be so. It is more important for the central pathways to maintain three-dimensional geometric requirements for sensorimotor function than for the endorgans to be in three orthogonal planes. The endorgans and central pathways certainly co-evolved, and perhaps the pathway symmetries have been a bit in advance.
The symmetries of the inner ear labyrinths, including the semicircular canals, are approximate, as is, for example, the mirror symmetry of one's right and left hands. However, the pattern of musculoskeletal connectivity of the two hands has a right-left reflection symmetry that is exact. Similarly, the structural symmetries of neural pathway connectivity can be exact even if the anatomical symmetry of sensory organs is only approximate.
Pathways of the CVS reach out to the accessory optic system, inner ear, and spinal cord on the sensory side. Activity then flows within the CVS and to other neural centers, such as the thalamus and cerebral cortex, reticular formation, and motor centers in the spinal cord and oculomotor nuclei.
4. Pathway Symmetries
To find symmetries of pathways, we consider transformations that are natural to the anatomy and physiology of the neuronal populations that give rise to the pathway. Permutations of the rotational planes are natural to the pathways we have investigated. In addition, the target neuronal population and the pathway itself suggest natural transformations. The next two subsections present specifics on two pathways.
4.1. A Canal-Neck Pathway
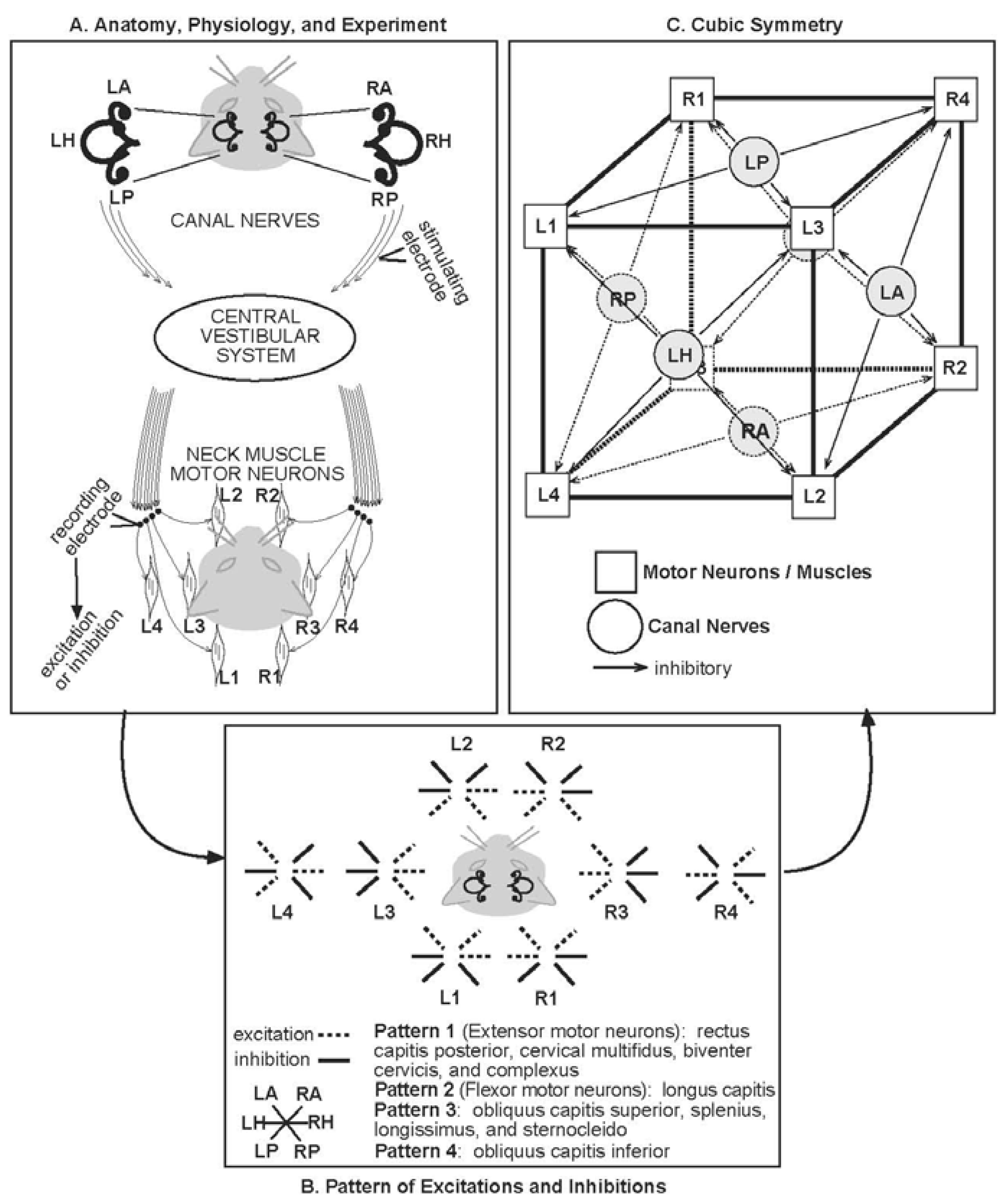
To perform its many sensory and motor functions, the head must often be moved relative to the shoulders by the neck muscles. The CVS integrates senses relevant to head movement and is strongly connected to neck motor neurons, the neurons that directly activate specific muscles. One way to probe the connection between the CVS and neck muscles is to electrically stimulate the nerves leading from the semicircular canals to the CVS and record the effect of that stimulation on neck motor neuron activity (Figure 1A).
The semicircular canals are convenient markers of the rotational planes in which much sensorimotor integration is organized. In Figure 1A, the canals are shown from above and exaggerated in the cat's head. There are three on the left (L) and three on the right (R). The horizontal canals (LH, RH) are approximately in the plane of the page. There are two vertical canals on each side, anterior (A) and posterior (P). The vertical canals (LA, LP, RA, RP) are projected onto the plane of the page. LA and RP are roughly in the same plane (LARP), which is transverse to the plane RA and LP approximately share (RALP).
Although in Figure 1A the stimulating electrode is only shown on the right, all six canal nerves were stimulated in the experiments [45,46,47]. There are left-right connections in the CVS which allow stimulation from all six canal nerves to affect all the neck motor neurons recorded. For the disynaptic pathway—one synapse in the CVS then one between the CVS neuron and the neck motor neuron—the effect of stimulation was a temporary increase (excitation) or decrease (inhibition) of activity in the motor neuron. The many motor neurons are represented as four dots on each side, connecting to schematic muscles. Eight patterns of excitation and inhibition were found, four on each side, which divided the various anatomical muscles into eight types (Figure 1B). These eight patterns are all the possible combinations of (excitation, inhibition) pairs in the three rotational planes, horizontal, LARP, and RALP.
The eight muscle types provide further natural transformations, besides the permutations of the rotational planes. These transformations include front-back and right-left reflections. Permutations of the muscle types form rotations like those of the eight vertices of a cube [15]. While the nervous system may segregate sub-patterns in ways as yet unforeseen, the pattern shown in Figure 1B has the symmetry group of the cube, the 11-element octahedral group O [36] (Figure 1C). The eight muscle types (L1-4, R1-4) are displayed on the vertices of the cube, and the six canals (LA, LH, LP, RA, RH, RP) occupy one face each. The pattern of Figure 1B allows the muscle types and canals to be arranged such that the four muscle types on each face are inhibited by stimulating the nerve of the canal on that face. (Stimulating the same canal nerve excites the remaining four muscle types.) It can then be readily demonstrated that symmetries of the cube, with the identified faces and vertices, correspond exactly with symmetries of the canal-neck pathway.
The cube is a three-dimensional object with three rotational planes, like the physical head. The octahedral symmetry of the canal-neck pathway suggests it provides three-dimensional geometrical constraints for head movements. Note that the neck muscles are arrayed around the neck as around a cylinder, and they are more parallel to the neck than perpendicular to it. They do not resemble a set of gears for turning in the three rotational planes. The rotational symmetry that we discovered in the Shinoda et al. [45,46,47] data was therefore a surprise, even though geometrically-accurate head movements had long been observed.
How would it benefit a nervous system to embody geometrical requirements that are followed ineluctably by all physical objects? Mechanical motions obey conservation of angular momentum, which is related by the Noether theorem to continuous rotational symmetries [48]. In the canal-neck pathway between discrete canals and discrete muscles, the CVS has formed a pattern that provides a discrete skeleton of these continuous rotational symmetries, which may allow the nervous system to anticipate the conservation of angular momentum. The symmetry group of the canal-neck pathway may thus provide a way for the organism to anticipate and to use body physics to its own advantage, in addition to integrating sensory and motor functions in such a way as to conform to three-dimensional geometric requirements.
4.2. Canal Pathway to the Uvula-Nodulus (CVOUN Pathway)
Physically, all rotations are in three-dimensional space, so the most straightforward way to cover them would be to build cubic symmetry into each pathway. However, the nervous system has a different and modular logic. Indeed, that is the question of interest: what is the logical structure by which the nervous system constructs perception and action? It is clear that the nervous system and the physical world do not match perfectly, because we make errors, we have perceptual illusions, and we get motion sickness [31,49]. Part of this difference in logical structure may arise from functions being handled by multiple pathways, each with different symmetries.
A second vestibular pathway in which a functionally-relevant symmetry is found runs from the vestibular nuclei to the uvula-nodulus, an evolutionarily ancient part of the cerebellum [23]. More completely, it can be considered the Canal pathway from the Vestibular nuclei via the inferior Olive to the Uvula-Nodulus or CVOUN pathway. Pathways from the vestibular nuclei to the cerebellum may have an even more pervasive influence on sensorimotor states than those to the spinal cord, as activity from the cerebellum reaches many neural centers besides the spinal cord.
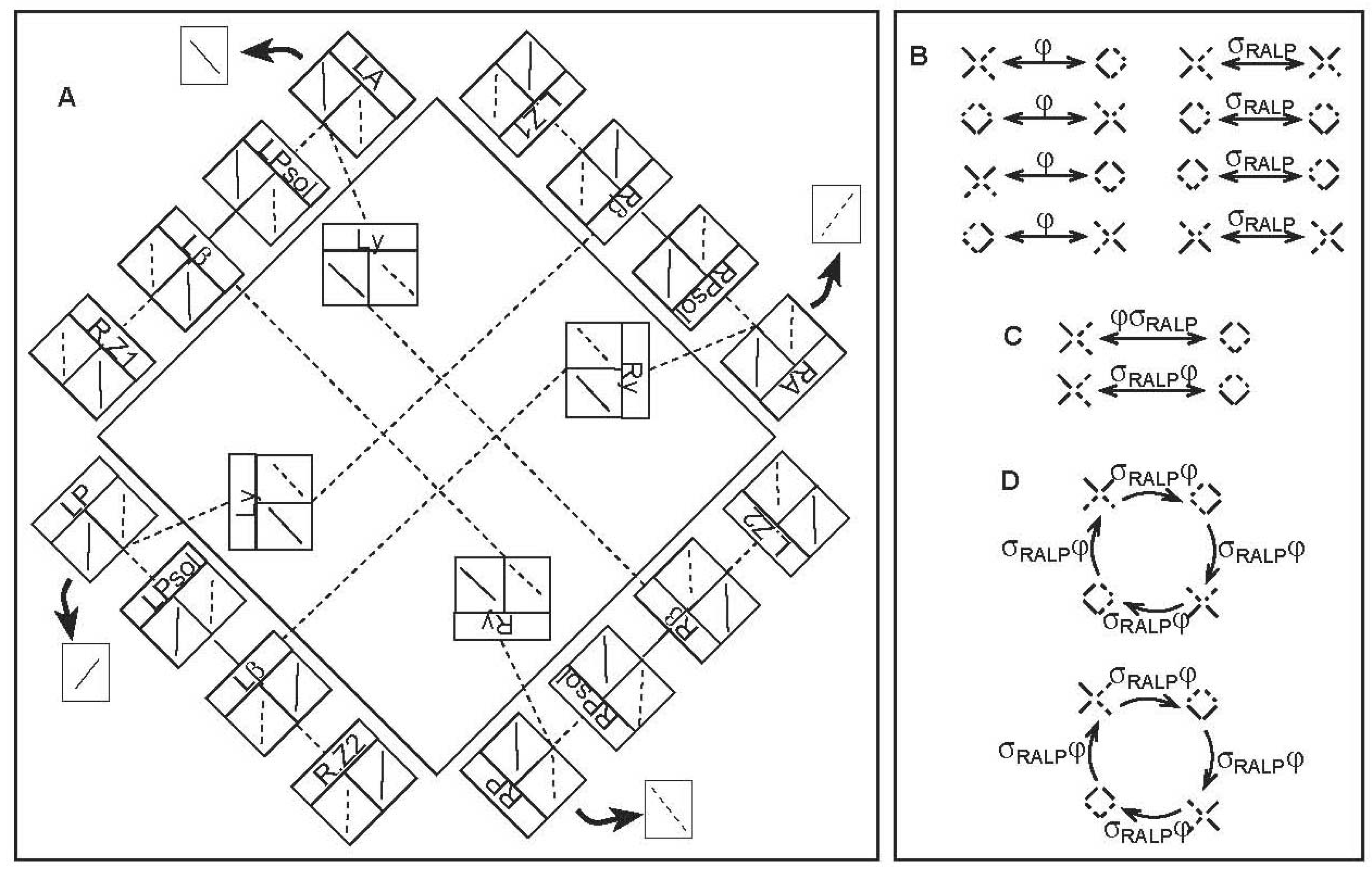
Unlike the canal-neck pathway, the CVOUN pathway has intermediate connections that are known to some precision [2]. The analysis of the symmetries relies, therefore, on the full anatomy and physiology rather than the pattern of activity in the target population of neurons [16] (Figure 2A). Also unlike the canal-neck pathway, sensory information from only the two vertical rotation planes, RALP and LARP, is included in this pathway. In Figure 2A, the anatomical pathways are drawn schematically, with an indication of the physiological recordings in the boxes.
Which transformations are reasonable to use on a pathway? This question arises here more sharply than in the canal-neck pathway. For example, the physical paths followed by nerves (axon bundles) are approximately symmetric in many cases, as the right and left hands and even the palm lines are often approximately symmetric. However, we are not interested in approximate symmetries that probably have little relevance to neural function; we are interested in exact symmetries with probable functional relevance. Such a symmetry is likely to arise among physiologically-natural transformations such as exchange of excitation and inhibition σ and exchange of rotational planes φ.
Consider the pathways diagrammed in Figure 2A. There are four parts, one mainly along each side of the large square. They start from the left and right anterior and posterior semicircular canals (LA, LP, RA, RP). Each box represents a neural center or part of a neural center. The neural centers are in the same sequence on each side of the large square. The four parts of the pathway interconnect, as shown by the dashed (excitatory) and solid (inhibitory) lines, in the same pattern on each side of the large square. There are four target zones in the uvula-nodulus: left and right zones 1 and 2 (L.Z1, R.Z1, L.Z2, R.Z2). Each part passes through the parasolitarius (Psol) of the vestibular nuclei (shown at the very bottom of the vestibular nuclei in Figure 1A), then the β nucleus of the inferior olive (a somewhat whimsically named neural center whose function may be rhythmic). The four parts are interconnected via the y-group of the vestibular nuclei, as represented by boxes in the middle of the large square. Note that the four parts separate into interconnecting pairs. The upper right and lower left are interconnected with each other and not with the other two. Further, the information that passes through these two interconnecting parts concerns only the RALP rotation plane; the plane is indicated by the angle of the lines in the boxes, lower left to upper right. (The line on the left tells the response to left tilt, excitatory dashed or inhibitory solid. The line on the right tells the response to right tilt.) Similarly, the upper right and lower left interconnecting parts of the pathway concern the LARP rotation plane alone.
The particular way the pathway connects its exclusively RALP and exclusively LARP parts determines the symmetry group of the pathway. Since the RALP and LARP parts do not interconnect, it is natural to consider exchange of excitation and inhibition on the RALP part only, σRALP, and on the LARP part only, σLARP. We will consider the transformations of the CVOUN pathway effected by φ, σRALP, and σLARP. Because of the similarity in pattern of the RALP and LARP parts of the pathway, σRALP and σLARP are conjugate by φ, and the three transformations can follow each other in compositions, so that they form a group.
Because the response to tilt is opposite in the two canals in a single rotational plane (RALP or LARP) and because this pairing of canal planes is established in the CVOUN pathway (Figure 2A), we can summarize the physiological state of each half of the pathway by a polarized pair of excitation and inhibition: one side is excited, and the other side is inhibited, of both RALP and LARP. This is denoted in Figure 2 by a diagonal line for each rotational plane, upper left to lower right for LARP and upper right to lower left for RALP, with the ends solid or dashed for inhibition or excitation, respectively. (Note that a similar polarization occurs in the canal-neck pathway, but in a very different way. There, each canal nerve is electrically excited, and the resulting response via the CVS is opposite, excitation and inhibition, in the two opposite canals of the same rotational plane.) Therefore, the physiological state of the CVOUN pathway can be summarized as a four-part diagram consisting of diagonal lines. The actual physiological responses to natural stimuli look like x's. For example, the response to a left tilt shown in the left-hand box (shown right-side-up by an arrow) under the semicircular canal at the beginning of each part of the pathway in Figure 2A can be combined into an x with two solid lines on the left and two dashed lines on the right. In order to consider the symmetries of the pathway, we will contemplate what happens to such a physiological state under the transformations φ, σRALP, and σLARP.
For each of the four parts of the CVOUN pathway, there are four possible states, two rotational planes and excitation or inhibition. Because the pairs must have the same rotational plane and be opposite in excitation and inhibition, there are only eight possible states of the whole CVOUN pathway. Since φ, σRALP, and σLARP are of order two, the square of each being the identity, they each divide the eight states of the CVOUN pathway into four pairs. The actions of φ and σRALP are shown in Figure 2B. (Further possibilities are explored in [17].) The transformations φ and σRALP do not commute, as shown in Figure 2C. Rather, σRALPφ and φσRALP are transformations of order four, as shown in Figure 2D.
The symmetry group of the CVOUN pathway is that of the square, D8, the dihedral group, including discrete rotations. As a subgroup of O, the symmetry group of the cube, it fixes one axis, presenting the symmetries of the faces orthogonal to that axis. (The subgroup that preserves the plane through the center of the cube, orthogonal to the axis, would also include also a vertical reflection.) In the CVOUN case, since transformations spatially permute only the vertical canal planes, we can identify this axis with the vertical axis through the head and the symmetry group D8 with symmetries in the horizontal plane.
Because we live and have evolved in a gravitational field, the horizontal plane plays a privileged role in sensorimotor behavior. It makes sense that the nervous system would have a separate module for the horizontal plane. In fact, the two vertical canals (leaving the horizontal plane fixed or tacit) evolved first [50]. Perhaps the horizontal plane was previously established by touch or other senses. The neck, and presumably the canal-neck pathway, is not found in most fish. It may be that a CVS pathway symmetry corresponding to full three-dimensional movement only developed in land animals with necks.
5. Sensorimotor States, Pathway Symmetries, and Causal Logic
The CVS occupies a juncture of the nervous system between sensory and motor, where it plays a strong role in shaping sensorimotor states. Symmetries relating to the three-dimensional geometry of physical space, into which perception and action must fit, are established in the anatomical and physiological connections of CVS pathways. These symmetries present the nervous system with menus of oscillations that can be used to compose perceptions and actions. The two CVS pathway symmetries we have presented differ, as befits a modular nervous system. We arrive at a picture in which sensorimotor states arise as or correspond to a patchwork of neural states, unified by related symmetries.
For example, reaching for an object often involves both a twist of the body and a glance to find the object in three-dimensional space. When the trunk maintains its vertical axis, even though the head and hand may move in a vertical direction, the behavior may be designed both in two and in three dimensions, in a coordinated fashion. A breakdown in such coordination may result in perceptual illusions and in motion sickness.
Complete sensorimotor coordination will presumably rely upon further CVS pathways, whose symmetries have not yet been evaluated. Will their symmetry groups always be subgroups of O, the symmetry group of the cube, which is found in the canal-neck pathway? A particularly salient question regards the symmetries of otolith pathways, which presumably evolved first [41]. There is extensive convergence between otolith and canal afferents in the CVS. As the semicircular canals evolved, convergence may have allowed canal activity to piggy-back on otolith pathways, using and differentiating their symmetries. In any case, separation of the two types of pathway is equivocal, and their symmetries are expected to be related.
The importance of symmetry in mechanics and therefore in sensorimotor behavior is well-established, in the form of conservation laws of energy and linear and angular momentum. These conservation laws are related to continuous symmetries by the Noether theorem [48]. For example, a continuous rotation about a vertical axis is related to its angular momentum. We have shown that in both the canal-neck and the CVOUN pathways there are discrete rotational symmetries. Because sensory receptors, neural pathways, and muscles are discrete, these discrete rotations may be the nervous system's skeleton for Noether symmetries. There is an additional interpretation which may be true at the same time and make the causal picture more interesting. A symmetry in a network of ordinary differential equations leads to oscillatory solutions [32]. Just as Noether's law shows that where there is a symmetry in physics, there is a conservation law, this relationship between discrete symmetry and oscillations would imply that where there is a symmetry in a neural center or pathway, there are symmetry subgroups shared among behaviors. The reverse would be that where there is symmetry in biological behavior, there is very likely to be an underlying neural symmetry, and it should be sought. It is possible that even an exactly symmetric behavior could be produced by heterogeneous neural connections, perfectly orchestrated, but a simpler and more elegant system would be to have behavior facilitated by the inherent structure of neural systems that bring it about. The pathway symmetries we have presented here induce networks on their targets, with symmetries. These symmetries are shared among oscillations that may arise in the pathway [17,36]. Oscillations of this sort may present a menu of sensorimotor options, already consistent with geometric requirements of physical space.
Our knowledge of the logical nature of causality in nervous systems is nascent and tentative. Symmetries of CVS pathways have been discussed in this review with regard to three facets of causal logic in nervous systems: connection, construction or autonomy, and modularity. The occurrence of CVS pathway symmetries in fundamental sensorimotor functions such as locomotion and gaze leads us to believe that they may play a major role in both initiating and shaping many sensorimotor behaviors.
Acknowledgments
We thank Thomas Chartrand for conversations about the Noether approach to symmetries and the contrast between that and the Golubitsky approach.
References and Notes
- Goldberg, J.M.; Fernández, C. The Vestibular System. In Handbook of Physiology—The Nervous System; Brookhart, J.M., Mountcastle, V.B., Darian-Smith, I., Eds.; American Physiological Society: Bethesda, MD, USA, 1984; Volume III, pp. 977–1022. [Google Scholar]
- Barmack, N.H. Central Vestibular System: Vestibular Nuclei and Posterior Cerebellum. Brain Res. Bull. 2003, 60, 511–541. [Google Scholar] [CrossRef]
- Boyle, R.; Belton, T.; McCrea, R.A. Responses of Identified Vestibulospinal Neurons to Voluntary and Reflex Eye and Head Movements in the Alert Squirrel Monkey. Ann. NY Acad Sci. 1996, 781, 244–263. [Google Scholar] [CrossRef] [PubMed]
- McCrea, R.A.; Gdowski, G.T.; Boyle, R.; Belton, T. Firing Behavior of Vestibular Neurons during Active and Passive Head Movements: Vestibulo-Spinal and Other Non-Eye-Movement Related Neurons. J. Neurophys. 1999, 82, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.E.; Cullen, K.E. Selective processing of vestibular reafference during self-generated head motion. J. Neurosci. 2001, 21, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.E.; Cullen, K.E. Dissociating Self-Generated from Passive Applied Head Motion: Neural Mechanisms in the Vestibular Nuclei. J. Neurosci. 2004, 24, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Cullen, K.E. Sensory signals during active versus passive movement. Curr. Opin. Neurobiol. 2004, 14, 698–706. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, M.K.; Cullen, K.E. Eye, Head, and Body Coordination during Large Gaze Shifts in Rhesus Monkeys: Movement Kinematics and the Influence of Posture. J. Neurophysiol. 2007, 97, 2976–2991. [Google Scholar] [CrossRef]
- Simpson, J.I. The Accessory Optic System. Ann. Rev. Neurosc. 1984, 7, 13–41. [Google Scholar] [CrossRef]
- Giolli, R.A.; Blanks, R.H.; Lui, F. The Accessory Optic System: Basic Organization with an Update on Connectivity, Neurochemistry, and Function. Prog. Brain Res. 2005, 151, 407–440. [Google Scholar]
- Hanes, D.A.; McCollum, G. Cognitive-Vestibular Interactions: A Review of Patient Difficulties and Possible Mechanisms. J. Vest. Res. 2006, 16, 75–91. [Google Scholar]
- Fukushima, K. Corticovestibular Interactions: Anatomy, Electrophysiology, and Functional Considerations. Exp. Brain Res. 1997, 117, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Brandt, T.; Schnautzer, F.; Hamilton, D.A.; Brüning, R.; Markowitsch, H.J.; Kalla, R.; Darlington, C.; Smith, P.; Strupp, M. Vestibular Loss Causes Hippocampal Atrophy and Impaired Spatial Memory in Humans. Brain 2005, 128, 2732–2741. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.F.; Horii, A.; Russell, N.; Bilkey, D.K.; Zheng, Y.; Liu, P.; Kerr, D.S.; Darlington, C.L. The Effects of Vestibular Lesions on Hippocampal Function in Rats. Prog. Neurobiol. 2005, 75, 391–405. [Google Scholar] [CrossRef] [PubMed]
- McCollum, G.; Boyle, R. Rotations in a Vertebrate Setting: Evaluation of the Symmetry Group of the Disynaptic Canal-neck Projection. Biol. Cybern. 2004, 90, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Foster, I.Z.; Hanes, D.A.; Barmack, N.H.; McCollum, G. Spatial Symmetries in Vestibular Projections to the Uvula-Nodulus. Biol. Cybern. 2007, 96, 439–453. [Google Scholar] [CrossRef] [PubMed]
- McCollum, G. Spatial Symmetry Groups as Sensorimotor Guidelines. J. Vest. Res. 2007, 17, 347–359. [Google Scholar]
- Lackner, J.R.; DiZio, P. Altered Sensory-Motor Control of the Head as an Etiological Factor in Space-Motion Sickness. Percept. Motor Skills 1989, 68, 784–786. [Google Scholar] [CrossRef]
- DiZio, P.; Lackner, J.R. Sensorimotor Aspects of High-Speed Artificial Gravity: III. Sensorimotor Adaptation. J. Vest. Res. 2002–2003, 12, 291–299. [Google Scholar]
- Clément, G.; Fraysse, M.J.; Deguine, O. Mental Representation of Space in Vestibular Patients with Otolithic or Rotatory Vertigo. Neurorep. 2009, 20, 457–461. [Google Scholar] [CrossRef]
- Lakoff, G.; Johnson, M. Metaphors We Live By; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Llinás, R.R. I of the Vortex; MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Crosby, E.C. Comparative Aspects of Cerebellar Morphology. In Neurobiology of Cerebellar Evolution and Development; Llinás, R., Ed.; American Medical Association: Chicago, IL, USA, 1969. [Google Scholar]
- Bressloff, P.C.; Cowan, J.D.; Golubitsky, M.; Thomas, P.J.; Wiener, M.C. Geometric Visual Hallucinations, Euclidean Symmetry, and the Functional Architecture of Striate Cortex. Phil. Trans. R. Soc. Lond. B 2001, 356, 299–330. [Google Scholar] [CrossRef]
- Bressloff, P.C.; Cowan, J.D. A Spherical Model of Orientation and Spatial-Frequency Tuning in a Cortical Hypercolumn. Phil. Trans. R. Soc. Lond. B 2002, 357, 1643–1667. [Google Scholar] [CrossRef] [PubMed]
- Bressloff, P.C. Spontaneous Symmetry Breaking in Self-Organizing Neural Fields. Biol. Cybern. 2005, 93, 256–274. [Google Scholar] [CrossRef]
- Chastain, E.; Liu, Y. Firing Fields of Dorsocaudal Medial Entorhinal Cortex as a Context-Independent Spatial Map; Number 06-02; Robotics Institute Technical Report: Pittsburgh, PA, USA, January 2006. [Google Scholar]
- Chastain, E.; Liu, Y. Quantified Symmetry for Entorhinal Spatial Maps. Neurocomputing 2007, 70, 1723–1727. [Google Scholar] [CrossRef]
- Rock, I. The Perception of Movement. In Indirect Perception; Rock, I., Palmer, S., Eds.; MIT Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Gregory, R.L. Knowledge in Perception and Illusion. Phil. Trans. R. Soc. Lond. B 1997, 352, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Holly, J.E.; McCollum, G. Constructive Perception of Self-Motion. J. Vest. Res. 2008, 18, 249–266. [Google Scholar]
- Golubitsky, M.; Stewart, I. The Symmetry Perspective: From Equilibrium to Chaos in Phase Space and Physical Space; Birkhauser Verlag: Basel, Switzerland, 2002. [Google Scholar]
- Llinás, R.R. The Intrinsic Electrophysiological Properties of Mammalian Neurons: Insights into Central Nervous System Function. Science 1988, 242, 1654–1664. [Google Scholar] [CrossRef] [PubMed]
- Llinás, R.R.; Ribary, U.; Jeanmonod, D.; Kronberg, E.; Mitra, P.P. Thalamocortical Dysrhythmia: A Neurological and Neuropsychiatric Syndrome Characterized by Magnetoencephalography. Proc. Nat. Acad. Sci. USA 1999, 96, 15222–15227. [Google Scholar] [CrossRef] [PubMed]
- Varela, F.; Lachaux, J.-P.; Rodriguez, E.; Martinerie, J. The Brainweb: Phase Synchronization and Large-Scale Integration. Nat. Rev. Neurosci. 2001, 2, 229–239. [Google Scholar] [CrossRef]
- Golubitsky, M.; Shiau, L.J.; Stewart, I. Spatiotemporal Symmetries in the Disynaptic Canal-Neck Projection. SIAM J. Appl. Math. 2007, 67, 1396–1417. [Google Scholar] [CrossRef]
- Carroll, S.B. Evo-Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell 2008, 134, 25–36. [Google Scholar] [CrossRef]
- Wagner, G.P.; Pavlicev, M.; Cheverud, J.M. The Road to Modularity. Nat. Rev. Genetics 2007, 8, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Wagner, G.P. Examining the Modularity Concept in Evolutionary Psychology: The Level of Genes, Mind, and Culture. J. Cult. Evol. Psychol. 2003, 1, 135–165. [Google Scholar] [CrossRef]
- Uzun-Coruhlu, H.; Curthoys, I.S.; Jones, A.S. Attachment of the Utricular and Saccular Maculae to the Temporal Bone. Hearing Res. 2007, 233, 77–85. [Google Scholar] [CrossRef]
- Fritsch, B.; Beisel, K.W.; Jones, K.; Fariñas, I.; Maklad, A.; Lee, J.; Reichardt, L.F. Development and Evolution of Inner Ear Sensory Epithelia and their Development. J. Neurobiol. 2002, 53, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Graf, W.M. Vestibular System. In Evolution of Nervous Systems: A Comprehensive Reference; Kaas, J.H., Krubitzer, L.A., Eds.; Academic Press: Amsterdam, The Netherland, 2007. [Google Scholar]
- Graf, W.; Klam, F. Le Système Vestibulaire: Anatomic Fontionelle et Comparée, Evolution et Développement. C. R. Palevol 2006, 5, 637–655. [Google Scholar] [CrossRef]
- Mach, E. Grundlinien der Lehre von den Bewegungsempfindungen (1875); Young, L.R.; Henn, V.; Scherberger, H., Translators; Leipzig: Engelmann, IL, USA, 2001. [Google Scholar]
- Shinoda, Y.; Sugiuchi, Y.; Futami, T.; Ando, N.; Kawasaki, T. Input Patterns and Pathways from Six Semicircular Canals to Motoneurons of Neck Muscles. I. The Multifidus Muscle Group. J. Neurophysiol. 1994, 72, 2691–702. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, Y.; Sugiuchi, Y.; Futami, T.; Kakei, S.; Izawa, Y.; Na, J. Four Convergent Patterns of Input from the Six Semicircular Canals to Motoneurons of Different Neck Muscles in the Upper Cervical Cord. Ann. NY Acad. Sci. 1996, 781, 264–75. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, Y.; Sugiuchi, Y.; Futami, T.; Ando, N.; Yagi, J. Input Patterns and Pathways from Six Semicircular Canals to Motoneurons of Neck Muscles. II. The Longissimus and Semispinalis Muscle Groups. J. Neurophysiol. 1997, 72, 2691–702. [Google Scholar] [CrossRef]
- Goldstein, H. Classical Mechanics, 2nd ed.; Addison-Wesley Publishing Company: Reading, MA, USA, 1980. [Google Scholar]
- Lackner, J.R.; DiZio, P. Vestibular, Proprioceptive, and Haptic Contributions to Spatial Orientation. Annu. Rev. Psychol. 2005, 56, 115–47. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.R.; Leverenz, E.L.; Bialek, W.S. The Vertebrate Inner Ear; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
Figure 1.
Canal-Neck Pathway Symmetry. This figure shows the way the symmetry occurs in the anatomy and physiology. Note that the semicircular canals (LH, LA, LP, RA, RP, RH) and the eight muscle types (L1-4 and R1-4) occur in each panel. Panel A provides a schematic diagram of the anatomy, along with the physiological experiment that revealed the symmetries. The three semicircular canals are shown exaggerated in the cat's head at the top. The sites of stimulation and recording are shown. Panel B depicts the recorded patterns of excitation and inhibition. The anatomically distinct muscles are divided into types by these patterns, as listed. Panel C displays a clear demonstration that the pattern in panel B has the symmetry group of the cube. L-left; R-right; A-anterior; H-horizontal; P-posterior (Parts of A and B are modified from [15]. C is modified from [36]).
Figure 1.
Canal-Neck Pathway Symmetry. This figure shows the way the symmetry occurs in the anatomy and physiology. Note that the semicircular canals (LH, LA, LP, RA, RP, RH) and the eight muscle types (L1-4 and R1-4) occur in each panel. Panel A provides a schematic diagram of the anatomy, along with the physiological experiment that revealed the symmetries. The three semicircular canals are shown exaggerated in the cat's head at the top. The sites of stimulation and recording are shown. Panel B depicts the recorded patterns of excitation and inhibition. The anatomically distinct muscles are divided into types by these patterns, as listed. Panel C displays a clear demonstration that the pattern in panel B has the symmetry group of the cube. L-left; R-right; A-anterior; H-horizontal; P-posterior (Parts of A and B are modified from [15]. C is modified from [36]).

Figure 2.
CVOUN Pathway and Its Transformations. A. Schematic diagram of the anatomy and physiology of the pathway. Excitatory connections are dashed, while inhibitory connections are solid. (Modified from [16]) B-D. Transformations of the state of the pathway shown in A, summarized as a four-part diagram composed of diagonal lines. An upper-left lower-right diagonal represents the LARP plane. An upper-right lower-left diagonal represents the RALP plane. Excitation is dashed; inhibition is solid. B. Order two transformations φ and σRALP. C. Non-commutativity of φ and σRALP. D. Transformations of the order four transformation σRALPφ (B-D modified from [17]).
Figure 2.
CVOUN Pathway and Its Transformations. A. Schematic diagram of the anatomy and physiology of the pathway. Excitatory connections are dashed, while inhibitory connections are solid. (Modified from [16]) B-D. Transformations of the state of the pathway shown in A, summarized as a four-part diagram composed of diagonal lines. An upper-left lower-right diagonal represents the LARP plane. An upper-right lower-left diagonal represents the RALP plane. Excitation is dashed; inhibition is solid. B. Order two transformations φ and σRALP. C. Non-commutativity of φ and σRALP. D. Transformations of the order four transformation σRALPφ (B-D modified from [17]).

© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
McCollum, G.; Hanes, D.A. Symmetries of the Central Vestibular System: Forming Movements for Gravity and a Three-Dimensional World. Symmetry 2010, 2, 1544-1558. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031544
AMA Style
McCollum G, Hanes DA. Symmetries of the Central Vestibular System: Forming Movements for Gravity and a Three-Dimensional World. Symmetry. 2010; 2(3):1544-1558. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031544
Chicago/Turabian StyleMcCollum, Gin, and Douglas A. Hanes. 2010. "Symmetries of the Central Vestibular System: Forming Movements for Gravity and a Three-Dimensional World" Symmetry 2, no. 3: 1544-1558. https://0-doi-org.brum.beds.ac.uk/10.3390/sym2031544