
The Potential Role of S-and Fe-Cycling Bacteria on the Formation of Fe-Bearing Mineral (Pyrite and Vivianite) in Alluvial Sediments from the Upper Chicamocha River Basin, Colombia

 ,
,
Abstract
:1. Introduction
2. Background Context
3. Materials and Methods
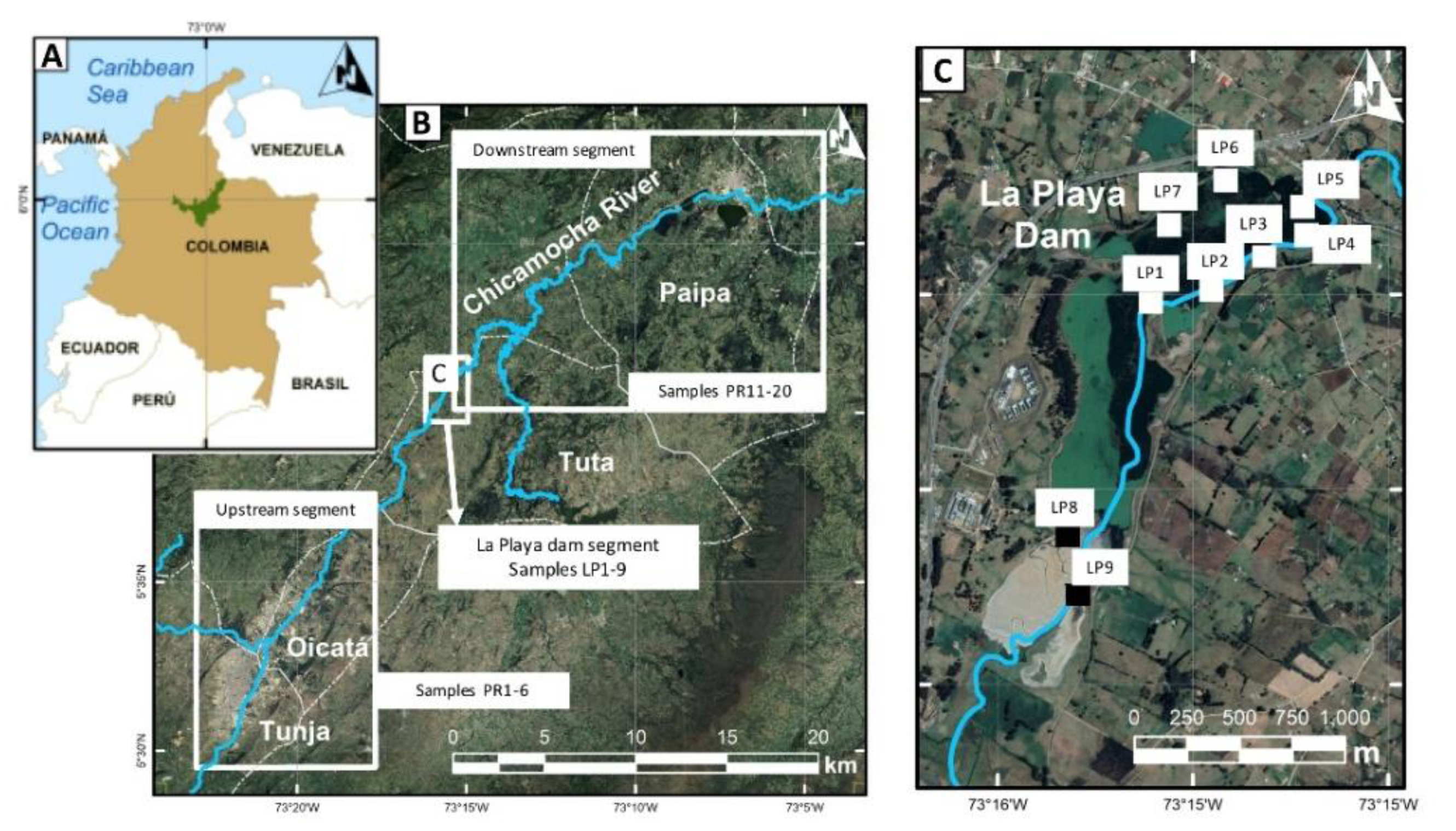
3.1. Materials
3.2. Mineralogical Methods
3.3. Microbiological Methods
4. Results
4.1. Fe-Bearing Minerals of the Sediments
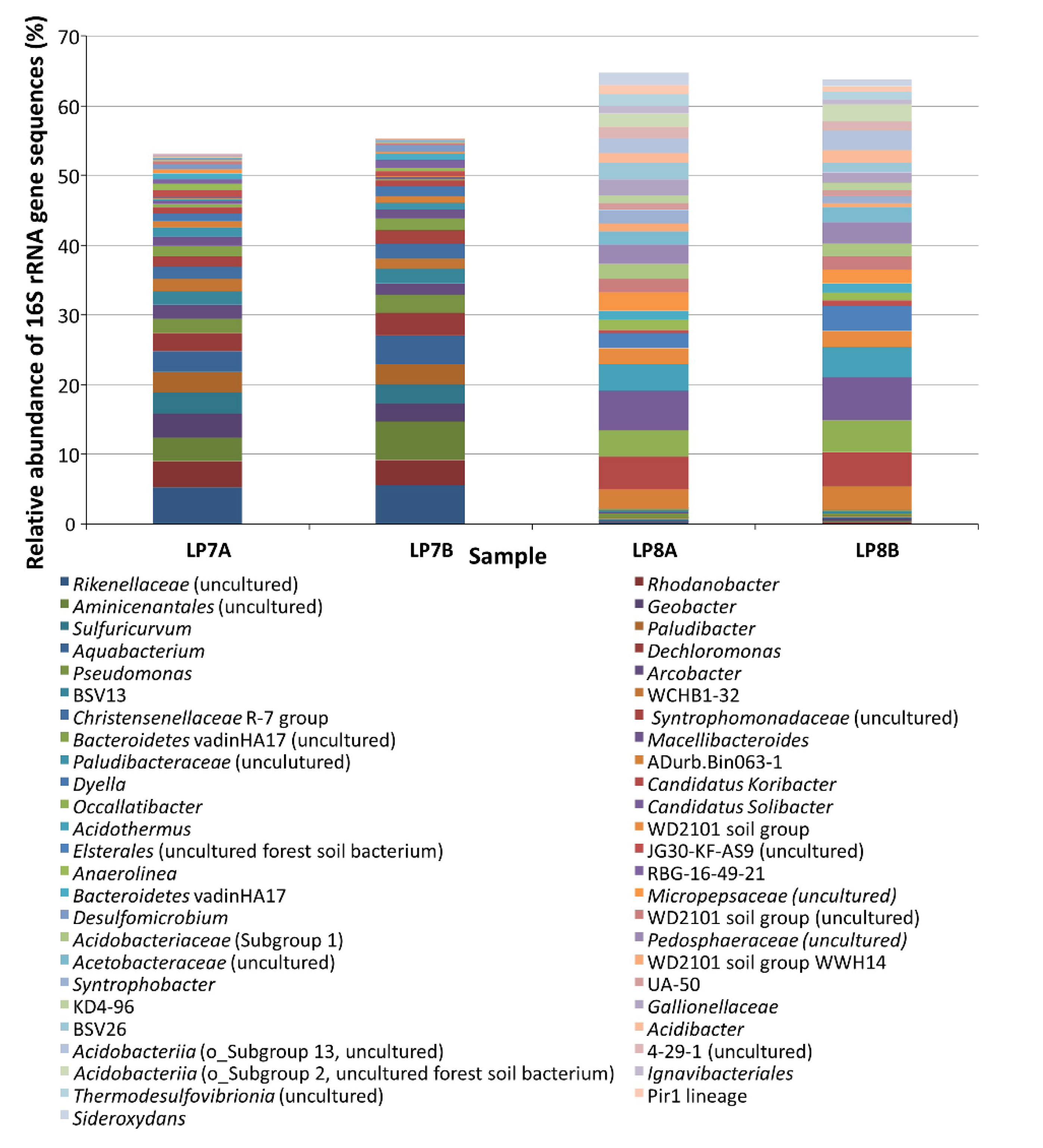
4.2. Bacterial Populations
5. Discussion
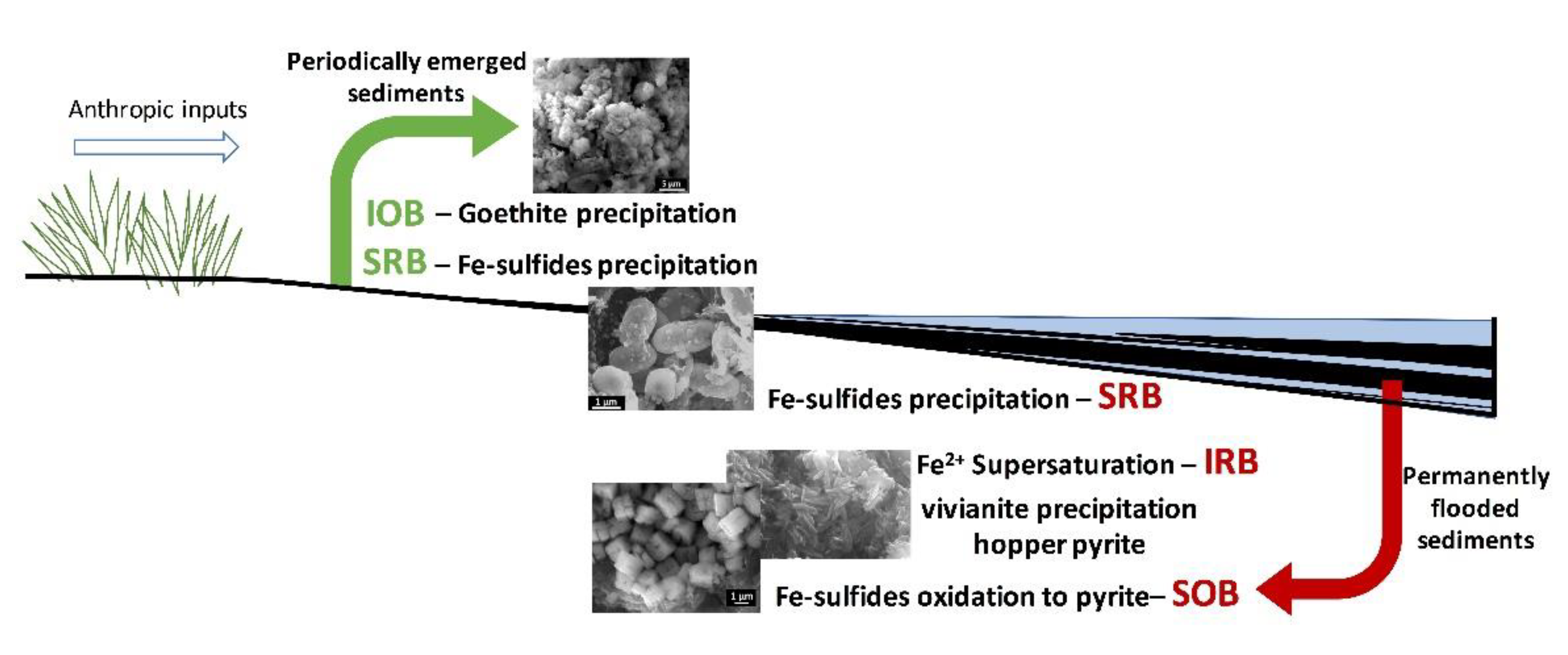
5.1. Mineral Distribution
5.2. The Role of the Bacterial Communities
6. Conclusions
- The high content in bacterial communities from the Bacteroidetes and Firmicutes phyla of the permanently flooded sediments of the La Playa dam reveal anthropic organic matter pollution processes (e.g., the presence of groups commonly found in feces, such as the Christensenellaceae family) and biodegradation associated with eutrophication (Rikenellaceae family, WCHB1-32, BVS13 and Macellibacteroides);
- The composition of the bacterial communities of the periodically emerged and dried sediments in the La Playa dam is characterized by the presence of groups frequently reported in high salinity soils (Verrucomicrobia and Actinobacteria) affected by the alternation of dry and wet periods (Candidatus Koribacter and Candidatus Solibacter) with oxic conditions (Planctomycetes), as well as by the presence of anaerobic microorganisms related to anoxic conditions (Anaerolineaceae);
- Both flooded and periodically emerged sediments show relevant SRB communities involved in the precipitation of Fe-sulfides (Figure 7). SEM images showing cell-shaped aggregates with a pyritic composition support the importance of the bacterial communities in the nucleation and transformation of sulfide minerals. The activity of these bacterial groups in the flooded sediments can be reinforced by syntrophic partners to produce H2 used by SRB (Syntrophomonadaceae) and increase the sulfide availability;
- IRB enrichment in the permanently flooded sediments of the La Playa dam and the presence of IOB in the periodically emerged sediments promotes a greater availability of Fe(II) in the flooded sediments, which favors the precipitation of vivianite by the contribution of microbial iron- and sulfur-reducing processes;
- Bacterial activity should favor supersaturation in Fe(II) (promoted by IRB and SRB) and sulfide (stimulated by SRB and their syntrophic partners that produce H2), which can be associated with the crystallization of hopper pyrite crystals in the permanently flooded sediments;
- Moreover, the SOB presence in the flooded sediments and the presence of Fe(III) due to aerobic conditions and the activity of IOB in the periodically emerged sediments can favor both pyrite crystallization under a high sulfide availability and the oxidation of microbially precipitated monosulfides. Moreover, IOB could enhance the precipitation of goethite in the periodically emerged sediments, even under low oxygen levels.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Noël, V.; Boye, K.; Kukkadapu, R.K.; Bone, S.; Pacheco, J.S.L.; Cardarelli, E.; Janot, N.; Fendorf, S.; Williams, K.H.; Bargar, J.R. Understanding controls on redox processes in floodplain sediments of the Upper Colorado River Basin. Sci. Total Environ. 2017, 603–604, 663–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, G.R.P.; Cuadros, J.; Partiti, C.M.S.; Cohen, R.; Vidal-Torrado, P. Sequential mineral transformation from kaolinite to Fe-illite in two Brazilian mangrove soils. Geoderma 2018, 309, 84–99. [Google Scholar] [CrossRef]
- Li, R.; Gao, L.; Wu, Q.; Liang, Z.; Hou, L.; Yang, Z.; Chen, J.; Jiang, T.; Zhu, A.; Li, M. Release characteristics and mechanisms of sediment phosphorus in contaminated and uncontaminated rivers: A case study in South China. Environ. Pollut. 2021, 268, 115749. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.M.; Griggs, G.B. Reductions in fluvial sediment discharge by coastal dams in California and implications for beach sustainability. J. Geol. 2003, 111, 167–182. [Google Scholar] [CrossRef]
- Khadse, G.K.; Meshram, D.B.; Deshmukh, P.; Labhasetwar, P.K. Water quality of Tehri dam reservoir and contributing rivers in the Himalayan region. India. Sustain. Water Resour. Manag. 2019, 5, 1951–19616. [Google Scholar] [CrossRef]
- Cifuentes, G.R.; Jiménez-Espinosa, R.; Jiménez-Millán, J.; Quevedo, C.P. Damming induced natural attenuation of hydrothermal waters by runoff freshwater dilution and sediment biogeochemical transformations (Sochagota Lake, Colombia). Water 2021, in press. [Google Scholar]
- Azam, H.M.; Finneran, K.T. Fe(III) reduction mediated phosphate removal as vivianite (Fe3(PO4)2·8H2O) in septic system wastewater. Chemosphere 2014, 97, 1–9. [Google Scholar] [CrossRef]
- Kokocinski, M.; Dziga, D.; Spoof, L.; Stefaniak, K.; Jurczak, T.; Mankiewicz-Boczek, J.; Meriluoto, J. First report of the cyanobacterial toxin cylindrospermopsin in the shallow, eutrophic lakes of western Poland. Chemosphere 2009, 74, 669–675. [Google Scholar] [CrossRef]
- Quevedo, C.P.; Jiménez-Millán, J.; Cifuentes, G.R.; Jiménez-Espinosa, R. Clay mineral transformations in anthropic organic matter-rich sediments under saline water environment. Effect on the detrital mineral assemblages in the upper Chicamocha river basin, Colombia. Appl. Clay Sci. 2020, 196, 105576. [Google Scholar] [CrossRef]
- Hoving, A.L.; Sander, M.; Bruggeman, C.; Behrends, T. Redox properties of clay-rich sediments as assessed by mediated electrochemical analysis: Separating pyrite, siderite and structural Fe in clay minerals. Chem. Geol. 2017, 457, 149–161. [Google Scholar] [CrossRef]
- Hu, S.Y.; Evans, K.; Fisher, L.; Rempel, K.; Craw, D.; Evans, N.J.; Cumberland, S.; Robert, A.; Grice, K. Associations between sulfides, carbonaceous material, gold and other trace elements in polyframboids: Implications for the source of orogenic gold deposits, Otago Schist, New Zealand. Geochim. Cosmochim. Acta 2016, 180, 197–213. [Google Scholar] [CrossRef]
- Cornell, R.M.; Schwertmann, U. The Iron Oxides: Structure, Properties, Reactions, Occurrences, and Uses, 2nd ed.; Wiley VCH: Weinheim, Germany, 2003. [Google Scholar]
- Cosmidis, J.; Benzerara, K.; Morin, G.; Busigny, V.; Lebeau, O.; Jézéquel, D.; Noël, V.; Dublet, G.; Othmane, G. Biomineralization of iron-phosphates in the water column of Lake Pavin (Massif Central, France). Geochem. Cosmochim. Acta 2014, 126, 78–96. [Google Scholar] [CrossRef]
- Watanabe, T.; Katayanagi, N.; Agbisit, R.; Llorca, L.; Yasukazu, H.; Susum, A. Influence of alternate wetting and drying water-saving irrigation practice on the dynamics of Gallionella-related iron-oxidizing bacterial community in paddy field soil. Soil Biol. Biochem. 2021, 152, 108064. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, H.; Huang, D.; Xu, C.; Zhu, H.; Zhu, Q. Water managements limit heavy metal accumulation in rice: Dual effects of iron-plaque formation and microbial communities. Sci. Total Environ. 2019, 687, 790–799. [Google Scholar] [CrossRef]
- Ji, Y.; Liu, P.; Conrad, R. Response of fermenting bacterial and methanogenic archaeal communities in paddy soil to progressing rice straw degradation. Soil Biol. Biochem. 2018, 124, 70–80. [Google Scholar] [CrossRef]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef]
- Qi, M.H.; Ma, S.S.; Qu, K.M.; Xin, F.Y. The formation of sulfide in the marine sediments and its relationships to cultivation of shellfish. Mar. Fish. Res. 2004, 25, 85–89. [Google Scholar]
- Niu, Z.S.; Pan, H.; Guo, X.P.; Lu, D.P.; Feng, J.N.; Chen, Y.R.; Tou, F.; Liu, M.; Yang, Y. Sulphate-reducing bacteria (SRB) in the Yangtze Estuary sediments: Abundance, distribution and implications for the bioavailibility of metals. Sci. Total Environ. 2018, 634, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Kiran, M.G.; Pakshirajan, K.; Das, G. Heavy metal removal from multicomponent system by sulfate reducing bacteria: Mechanism and cell surface characterization. J. Hazard. Mater. 2017, 324, 62–70. [Google Scholar] [CrossRef]
- Tarekegn, M.M.; Zewdu, F.; Iniyehu, A. Microbes used as a tool for bioremediation of heavy metal from the environment. Cogent Food Agric. 2020, 6, 1783174. [Google Scholar] [CrossRef]
- Wolfenden, S.; Charnock, J.M.; Hilton, J.; Livens, F.R.; Vaughan, D.J. Sulfide species as a sink for mercury in lake sediments. Environ. Sci. Technol. 2005, 39, 6644–6648. [Google Scholar] [CrossRef]
- Kühl, M.; Jørgensen, B.B. Microsensor measurements of sulfate reduction and sulfide oxidation in compact microbial communities of aerobic biofilms. Appl. Environ. Microbiol. 1992, 58, 1164–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.; Shaman, A.K. An integrated microbial process for the bioremediation of soil contaminated with toxic metals. Nat. Biotechnol. 1998, 16, 572–575. [Google Scholar] [CrossRef]
- Corpoboyacá; Corporación Autónoma Regional de Boyacá; Pedagogical and Technological University of Colombia. Plan de Ordenación y Manejo Ambiental de la Cuenca Alta del Río Chicamocha; Corpoboyacá—Corporación Autónoma Regional de Boyacá; Pedagogical and Technological University of Colombia: Boyacá, Colombia; Universidad Nacional de Colombia: Bogotá, Colombia, 2006. [Google Scholar]
- Márquez, G.; Guillot, G. Ecología y Efecto Ambiental de Embalses—Aproximación a Casos Colombianos; Universidad Nacional de Colombia: Bogotá, Colombia, 2001; Volume 218. [Google Scholar]
- Quevedo, C.P.; Jiménez-Millán, J.; Cifuentes, G.R.; Jiménez-Espinosa, R. Electron microscopy evidence of Zn bioauthigenic sulfides formation in polluted organic matter-rich sediments from the Chicamocha River (Boyacá-Colombia). Minerals 2020, 10, 673. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Manning, P.G.; Prepas, E.E.; Serediak, M.S. Pyrite and vivianite intervals in the bottom sediments of eutrophic Baptiste Lake, Alberta, Canada. Can. Miner. 1999, 37, 593–601. [Google Scholar]
- García-Ruiz, J.M.; Otálora, F. Crystal Growth in Geology: Patterns on the Rocks. In Handbook of Crystal Growth; Nishinaga, T., Rudolph, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; Volume II, pp. 1–43. [Google Scholar]
- Berg, J.S.; Jézéquel, D.; Duverger, A.; Lamy, D.; Laberty-Robert, C.; Miot, J. Microbial diversity involved in iron and cryptic sulfur cycling in the ferruginous, low-sulfate waters of Lake Pavin. PLoS ONE 2019, 14, e0212787. [Google Scholar] [CrossRef]
- Rothe, M.; Kleeberg, A.; Hupfer, M. The occurrence, identification and environmental relevance of vivianite in waterlogged soils and aquatic sediments. Earth Sci. Rev. 2016, 158, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.S.; Duverger, A.; Cordier, L.; Laberty-Robert, C.; Guyot, F.; Miot, J. Rapid pyritization in the presence of a sulfur/sulfate-reducing bacterial consortium. Sci. Rep. 2020, 10, 8264. [Google Scholar] [CrossRef]
- Duverger, A.; Berg, J.S.; Busigny, V.; Guyot, F.; Bernard, S.; Miot, J. Mechanisms of Pyrite Formation Promoted by Sulfate-Reducing Bacteria in Pure Culture. Front. Earth Sci. 2020, 8, 588310. [Google Scholar] [CrossRef]
- Egger, M.; Jilbert, T.; Behrends, T.; Rivard, C.; Slomp, C.P. Vivianite is a major sink for phosphorus in methanogenic coastal surface sediments. Geochim. Cosmochim. Acta 2015, 169, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, N.; Slomp, C.P.; Behrends, T. Vivianite is a key sink for phosphorus in sediments of the Landsort Deep, an intermittently anoxic deep basin in the Baltic Sea. Chem. Geol. 2016, 438, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Liu, J.; Gao, B.; Sillanpää, M. Phosphate substances transformation and vivianite formation in P-Fe containing sludge during the transition process of aerobic and anaerobic conditions. Bioresour. Technol. 2021, 319, 124259. [Google Scholar] [CrossRef] [PubMed]
- Druschel, G.K.; Emerson, D.; Sutka, R.; Suchecki, P.; Luther III, G.W. Low-oxygen and chemical kinetic constraints on the geochemical niche of neutrophilic iron(II) oxidizing microorganisms. Geochim. Cosmochim. Acta 2008, 72, 3358–3370. [Google Scholar] [CrossRef]
- Wu, Y.F.; Xing, P.; Liu, S.; Wu, Q.L. Enhanced Microbial Interactions and Deterministic Successions During Anoxic Decomposition of Microcystis Biomass in Lake Sediment. Front. Microbiol. 2019, 10, 2474. [Google Scholar] [CrossRef]
- Chang, C.H.; Wei, C.C.; Lin, L.H.; Tu, T.H.; Liao, V.H.C. Humic acids enhance the microbially mediated release of sedimentary ferrous iron. Environ. Sci. Pollut. Res. 2016, 23, 4176–4184. [Google Scholar] [CrossRef]
- Li, H.; Xing, P.; Wu, Q.L. Characterization of the bacterial community composition in a hypoxic zone induced by Microcystis blooms in lake Taihu. China. FEMS Microbiol. Ecol. 2012, 79, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Li, B.; Feng, G.; Zhang, C.; He, J.; Li, H.; Zhu, J. Responses of bacterial communities and organic matter degradation in surface sediment to Macrobrachium nipponense bioturbation. Sci. Total Environ. 2021, 759, 143534. [Google Scholar] [CrossRef] [PubMed]
- Timmers, P.H.A.; Vavourakis, C.D.; Kleerebezem, R.; Damsté, J.S.S.; Muyzer, G.; Stams, A.J.M.; Sorokin, D.Y.; Plugge, C.M. Metabolism and Occurrence of Methanogenic and Sulfate-Reducing Syntrophic Acetate Oxidizing Communities in Haloalkaline Environments. Front. Microbiol. 2018, 9, 3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacerda-Júnior, G.V.; Noronha, M.F.; Cabral, L.; Delforno, T.P.; de Sousa, S.T.P.; Fernandes-Júnior, P.I.; Melo, I.S.; Oliveira, V.M. Land Use and Seasonal Effects on the Soil Microbiome of a Brazilian Dry Forest. Front. Microbiol. 2019, 10, 648. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Samaddar, S.; Chatterjee, P.; Ramasamy, K.; Jeon, S.; Sa, T. Structural and functional responses of microbial community with respect to salinity levels in a coastal reclamation land. Appl. Soil Ecol. 2019, 137, 96–105. [Google Scholar] [CrossRef]
- Polivkova, M.; Suman, J.; Strejcek, M.; Kracmarova, M.; Hradilova, M.; Filipova, A.; Cajthaml, T.; Macek, T.; Uhlik, O. Diversity of root-associated microbial populations of Tamarix parviflora cultivated under various conditions. Appl. Soil Ecol. 2018, 125, 264–272. [Google Scholar] [CrossRef]
- Radhakrishnan, M.; Imchen, M.; Kaari, M.; Angamuthu, V.; Venugopal, G.; Shanmugasundaram, T.; Joseph, J.; Balagurunathan, R.; Kumavath, R. Metagenomic insights unveil the dominance of undescribed Actinobacteria in pond ecosystem of an Indian shrine. Meta Gene 2019, 23, 100639. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Ivanova, A.A. Planctomycetes in boreal and subarctic wetlands: Diversity patterns and potential ecological functions. FEMS Microbiol. Ecol. 2019, 95, fiy227. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Hetharua, B.; Lin, L.; Xu, H.; Zheng, T.; He, Z.; Tian, Y. Mangrove Sediment Microbiome: Adaptive Microbial Assemblages and Their Routed Biogeochemical Processes in Yunxiao Mangrove National Nature Reserve, China. Microb. Ecol. 2019, 78, 57–69. [Google Scholar] [CrossRef]
- Cifuentes, G.R.; Jiménez-Millán, J.; Quevedo, C.P.; Gálvez, A.; Castellanos-Rozo, A.; Jiménez-Espinosa, R. Trace element fixation in sediments rich in organic matter from a saline lake in tropical latitude with hydrothermal inputs (Sochagota Lake, Colombia): The role of bacterial communities. Sci. Total Environ. 2021, 762, 143113. [Google Scholar] [CrossRef]
- Iino, T.; Mori, K.; Uchino, Y.; Nakagawa, T.; Harayama, S.; Suzuki, K. Ignavibacterium album gen. nov., sp. nov., a moderately thermophilic anaerobic bacterium isolated from microbial mats at a terrestrial hot spring and proposal of Ignavibacteria classis nov., for a novel lineage at the periphery of green sulfur bacteria. Int. J. Syst. Evol. Microbiol. 2010, 60, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Hazra, C.; Kundu, D.; Chaudhari, A.; Jana, T. Biogenic synthesis, characterization, toxicity and photocatalysis of zinc sulfide nanoparticles using rhamnolipids from Pseudomonas aeruginosa, BS01 as capping and stabilizing agent. J. Chem. Technol. Biotechnol. 2013, 88, 1039–1048. [Google Scholar] [CrossRef]
- Zampieri, B.D.B.; da Costa Andrade, V.; Chinellato, R.M.; Garcia, C.A.B.; de Oliveira, M.A.; Brucha, G.; Fernandes Cardoso de Oliveira, A.J. Heavy metal concentrations in Brazilian port areas and their relationships with microorganisms: Can pollution in these areas change the microbial community? Environ. Monit. Assess. 2020, 192, 512. [Google Scholar] [CrossRef]
- Ming, H.; Jingfeng, F.; Liu, J.; Jie, S.; Zhiyi, W.; Yantao, W.; Dongwei, L.; Mengfei, L.; Tingting, S.; Yuan, J.; et al. Full-length 16S rRNA gene sequencing reveals spatiotemporal dynamics of bacterial community in a heavily polluted estuary, China. Environ. Pollut. 2021, 275, 116567. [Google Scholar] [CrossRef]
- Tu, Y.T.; Chiang, P.C.; Yang, J.; Chen, S.H.; Kao, C.M. Application of a constructed wetland system for polluted stream remediation. J. Hydrol. 2014, 510, 70–78. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, L.; Hou, L.; Yang, Y.; Wu, D.; Meadows, M.; Li, Z.; Tong, C.; Gu, J. Occurrence and distribution of PAHs and microbial communities in nearshore sediments of the Knysna Estuary, South Africa. Environ. Pollut. 2020, 270, 116083. [Google Scholar] [CrossRef]
- Li, C.H.; Yan, K.; Tang, L.S.; Jia, Z.J.; Li, Y. Change in deep soil microbial communities due to long-term fertilization. Soil Biol. Biochem. 2014, 75, 264–272. [Google Scholar] [CrossRef]
- Geesink, P.; Wegner, C.E.; Probst, A.J.; Herrmann, M.; Dam, H.T.; Kaster, A.K.; Küsel, K. Genome-inferred spatio-temporal resolution of an uncultivated Roizmanbacterium reveals its ecological preferences in groundwater. Environ. Microbiol. 2020, 22, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Guo, C.; Yang, C.; Lu, G.; Chen, M.; Dang, Z. Distribution and diversity of bacterial communities and sulfate-reducing bacteria in a paddy soil irrigated with acid mine drainage. J. Appl. Microbiol. 2016, 121, 13143. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Z.; Cai, H.; Lu, W.; Li, J. Long-term agricultural contamination shaped diversity response of sediment microbiome. J. Environ. Sci. 2021, 99, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Andrea, I.; Triana, D.; Sanz, J.L. Bioremediation of acid mine drainage coupled with domestic wastewater treatment. Water Sci. Technol. 2012, 66, 2425–2431. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiao, T.; Ning, Z.; Li, Q.; Xiao, E.; Liu, Y.; Xiao, Q.; Lan, X.; Ma, L.; Lu, F. In-situ remediation of acid mine drainage from abandoned coal mine by filed pilot-scale passive treatment system: Performance and response of microbial communities to low pH and elevated Fe. Bioresour. Technol. 2020, 317, 123985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, W.C.; Liu, Y.; Zhang, H.L.; Zhao, Z.H.; Zou, L.Y.; Shen, Y.C.; Lan, W.S. Impacts of anthropogenic disturbances on microbial community of coastal waters in Shenzhen, South China. Ecotoxicology 2020, 30, 1652–1661. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | SiO2 | Al2O3 | FeO | S | LOI | TOC | Clay | Qz | Py | Viv | Gth | pH | R.P. | E.C. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LP 1 | 61.34 | 19.21 | 4.21 | 27201 | 17.21 | 14.51 | 44 | 44 | 7 | 5 | - | 7.0 | −54 | 1431 |
| LP 2 | 60.83 | 18.74 | 4.38 | 24534 | 18.34 | 14.88 | 42 | 50 | 5 | <5 | - | 7.0 | −32 | 1254 |
| LP 3 | 62.71 | 17.62 | 4.75 | 23244 | 16.99 | 13.99 | 42 | 48 | 5 | 5 | - | 7.1 | −21 | 1572 |
| LP 4 | 60,32 | 19.23 | 3.88 | 18321 | 17.61 | 14.76 | 46 | 43 | 6 | 5 | - | 7.2 | −31 | 1423 |
| LP 5 | 61.12 | 18.31 | 4.04 | 26523 | 18.44 | 14.41 | 43 | 48 | 5 | <5 | - | 7.0 | −12 | 1621 |
| LP 6 | 60.21 | 17.21 | 3.89 | 17453 | 15.33 | 13.80 | 47 | 46 | 5 | <5 | - | 6.9 | −27 | 1599 |
| LP 7 | 63.24 | 18.24 | 4.11 | 21956 | 10.51 | 4.29 | 45 | 46 | 5 | 6 | - | 7.0 | −32 | 1324 |
| LP 8 | 61.31 | 15.99 | 3.81 | 11345 | 16.22 | 11.00 | 45 | 42 | 7 | - | 6 | 7.0 | −5 | 2431 |
| LP 9 | 60.18 | 17.22 | 4.38 | 15781 | 15.43 | 13.80 | 43 | 45 | 5 | - | 7 | 7.0 | 2 | 1854 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quevedo, C.P.; Jiménez-Millán, J.; Cifuentes, G.R.; Gálvez, A.; Castellanos-Rozo, J.; Jiménez-Espinosa, R. The Potential Role of S-and Fe-Cycling Bacteria on the Formation of Fe-Bearing Mineral (Pyrite and Vivianite) in Alluvial Sediments from the Upper Chicamocha River Basin, Colombia. Minerals 2021, 11, 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101148
Quevedo CP, Jiménez-Millán J, Cifuentes GR, Gálvez A, Castellanos-Rozo J, Jiménez-Espinosa R. The Potential Role of S-and Fe-Cycling Bacteria on the Formation of Fe-Bearing Mineral (Pyrite and Vivianite) in Alluvial Sediments from the Upper Chicamocha River Basin, Colombia. Minerals. 2021; 11(10):1148. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101148
Chicago/Turabian StyleQuevedo, Claudia Patricia, Juan Jiménez-Millán, Gabriel Ricardo Cifuentes, Antonio Gálvez, José Castellanos-Rozo, and Rosario Jiménez-Espinosa. 2021. "The Potential Role of S-and Fe-Cycling Bacteria on the Formation of Fe-Bearing Mineral (Pyrite and Vivianite) in Alluvial Sediments from the Upper Chicamocha River Basin, Colombia" Minerals 11, no. 10: 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/min11101148