Machine Learning Algorithms Applied to Identify Microbial Species by Their Motility

1
Astrobiology Group, Center of Astronomy and Astrophysics, Technical University Berlin, 10623 Berlin, Germany
2
Section Geomicrobiology, GFZ German Center for Geosciences, 14473 Potsdam, Germany
3
Department of Experimental Limnology, Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), 16775 Stechlin, Germany
4
School of the Environment, Washington State University, Pullman, WA 99164, USA
*
Author to whom correspondence should be addressed.
Life 2021, 11(1), 44; https://0-doi-org.brum.beds.ac.uk/10.3390/life11010044
Submission received: 14 December 2020
/
Revised: 7 January 2021
/
Accepted: 8 January 2021
/
Published: 12 January 2021
(This article belongs to the Section Astrobiology)
Abstract
:(1) Background: Future missions to potentially habitable places in the Solar System require biochemistry-independent methods for detecting potential alien life forms. The technology was not advanced enough for onboard machine analysis of microscopic observations to be performed in past missions, but recent increases in computational power make the use of automated in-situ analyses feasible. (2) Methods: Here, we present a semi-automated experimental setup, capable of distinguishing the movement of abiotic particles due to Brownian motion from the motility behavior of the bacteria Pseudoalteromonas haloplanktis, Planococcus halocryophilus, Bacillus subtilis, and Escherichia coli. Supervised machine learning algorithms were also used to specifically identify these species based on their characteristic motility behavior. (3) Results: While we were able to distinguish microbial motility from the abiotic movements due to Brownian motion with an accuracy exceeding 99%, the accuracy of the automated identification rates for the selected species does not exceed 82%. (4) Conclusions: Motility is an excellent biosignature, which can be used as a tool for upcoming life-detection missions. This study serves as the basis for the further development of a microscopic life recognition system for upcoming missions to Mars or the ocean worlds of the outer Solar System.
1. Introduction
Every planetary mission so far has used optics for direct imaging at macroscopic scales. However, imaging was used mainly for geological purposes in these missions, and so microscopes on rovers or landers have not been employed to detect life [1]. One major problem with using microscopic in-situ observations to search for microbial organisms is that there are not many features that distinguish cells from abiotic sediment particles. The unambiguous distinction of primitive life forms such as bacteria and archaea from mineral particles even with an electron microscope is not always possible [2]. Thus, to detect life unambiguously, chemical composition or microbial motility has to be added as a criterion [1].
Microbial motility is the property of the movement of a cell using its own power. Motility is common to all three domains of life (bacteria, archaea, and eukaryotes) and is stimulated by environmental conditions [3]. This ability to move is crucial for many organisms because it enables them to avoid toxins and find new resources and nutrients. Many prokaryotes use flagella for propulsion. Flagella are filamentous appendages that rotate at up to 300 revolutions per second and propel the cell through liquid media at up to 60 cell lengths per second [4]. The amino acid sequence of flagellin—an essential protein of the flagella—is highly conserved in bacterial species, suggesting that flagellar motility evolved early and has deep phylogenetic roots. The fundamental differences of the flagellar protein structure in archaea and bacteria indicate that flagellar motility evolved separately in these two prokaryotic cell types, presumably after they diverged over 3.5 billion years ago [4]. The main characteristic of bacterial motility is speed and the motion pattern. A quantitative description of the distribution of rotational motions and the speed in spatiotemporal space describes the bacterial movement and allows the characterization of different motility behaviors. The values of these parameters are characteristic not only for each specific species but also for the environmental conditions. Different forms of motility patterns have been qualitatively described by Reference [5]. The strains we used in our experiments show dominantly run-and-tumble motion (Escherichia coli and Bacillus subtilis), or a run-and-reverse-flick pattern (Pseudoalteromonas haloplanktis and Planococcus halocryophilus), but an extensive quantitative evaluation is missing. Previous research indicates that the average translational velocity of all bacteria seems to increase as the environmental temperature increases [6].
The main unknown question, assuming the existence of extraterrestrial microbial life, is whether it would move using its own propulsion (like using flagella or a similar system) and, if so, how it might move. Because motility is such an advantageous characteristic and one which evolved very early in the evolution of life on Earth, we suggest that it is likely that extraterrestrial life has developed this property as well [4]. If so, then the distinction between Brownian motion and microbial motility should be straightforward and is expected to have high accuracy when applying machine learning algorithms.
It would be ideal to make the distinction with a minimum set of parameters. This is important because one of the challenges of planetary missions is the constrained link budgets. Only limited amounts of data can be transmitted from a lander—either via a relay or directly—to Earth. Therefore, it is crucial that most of the data can be analyzed autonomously. It is also advantageous to determine beforehand which data should be transmitted autonomously and which not. An optimal engineering solution is a trade-off between the computing power required for the autonomous processes and the link budget, which requires detailed knowledge of the mission concept. The software solution needs to be a trade-off between false positive and false negative rates (depending on the link budget, a bias toward false positive might be preferable, but in any case, the ground operators should re-evaluate the transferred data). This study aims to identify life based on motility by using supervised machine learning algorithms, showing that identification of microbial species is possible by analyzing their motility and collecting detailed quantitative data, which complement the qualitative descriptions performed in previous attempts. Here, we address the identification of motility with tools that can be extended to use in space missions.
2. Materials and Methods
2.1. Microorganisms Used
Aside from the well-known Escherichia coli bacterium, which has been commonly used in many previous experiments, we have also tested our detection technique using Bacillus subtilis, Planococcus halocryophilus, and Pseudoalteromonas haloplanktis. B. subtilis is a common and resistant soil bacterium. P. halocryophilus and P. haloplanktis are adapted to salty and cold environments, making them suitable analog organisms for life detection on Mars and the ocean worlds such as Europa and Enceladus.
Escherichia coli is the most widely studied prokaryotic organism. It is a rod-shaped, facultative anaerobe gram-negative bacterium, which lives in the guts of warm-blooded organisms. The organisms are about 2 µm long, 1 µm in diameter, and are motile due to the possession of flagella [4]. It is a peritrichous bacterium, meaning it has 5–10 flagella, which are randomly distributed across the cell surface [7]. We used E. coli K-12 wild-type (DSM 498) in our study, obtained from Dirk Wagner, Helmholtz Centre Potsdam GFZ German Research Centre for Geosciences.
Bacillus subtilis is a facultative anaerobic bacterium and one of the best-studied gram-positive organisms. B. subtilis occurs in the natural soil environment and is also found in human’s gastrointestinal tract [8]. It is rod-shaped and can survive in extreme conditions in terms of temperatures and desiccation because it is able to form endospores. It is typically 4 to 10 µm long and 0.25 to 1 µm in diameter [9]. We used the type strain B. subtilis Marburg (DSM 10), strain designation Marburg, obtained from Dirk Wagner, Helmholtz Centre Potsdam GFZ German Research Centre for Geosciences. This strain moves by using approximately 20 flagella per cell, which are non-randomly distributed over the cell surface [10].
Pseudoalteromonas haloplanktis is a curved, rod-shaped psychrophilic bacterium isolated from Antarctica. It is typically 1.2 to 2.3 µm long and 0.5 to 0.6 µm wide. This strictly aerobic extremophile is gram-negative, non-spore-forming, and motile, employing a single polar flagellum [11]. It has its growth optimum at 26 °C and can grow at temperatures as low as 0 °C [12]. Due to its ability to survive in cold environments, we used the type strain Pseudoalteromonas haloplanktis 545 (DSM 6060), obtained from DSMZ (Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures GmbH).
Planococcus halocryophilus is a gram-positive bacterium. It is aerobic and found in the arctic permafrost. The cells are non-spore-forming cocci, which occur individually or in pairs, and have a diameter of between 0.8 and 1.2 µm. The bacterium is motile and thrives in environments of high salinity and low temperature [13]. It grows at temperatures as low as −10 °C and is metabolically active at temperatures as low as −25 °C [14]. Cells remain motile under all growth conditions [14]. The strain was first isolated from a permafrost active layer-soil in Ellesmere Island, Eureka, Canada. We used the type strain Planococcus halocryophilus Or1 (DSM 24743), obtained from DSMZ (Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures GmbH).
2.2. Incubation of Microorganisms
Before conducting the microscopic measurements at different temperatures, the microbial strains were allowed to grow to the late exponential phase. E. coli and B. subtilis were both grown for 24 h at 30 °C in standard nutrient broth S1. P. haloplanktis was incubated for 48 h at 25 °C in bacto marine broth (DIFCO 2216), and P. halocryophilus was grown for 48 h at 25 °C in trypticase soy yeast extract medium.
2.3. Measurements
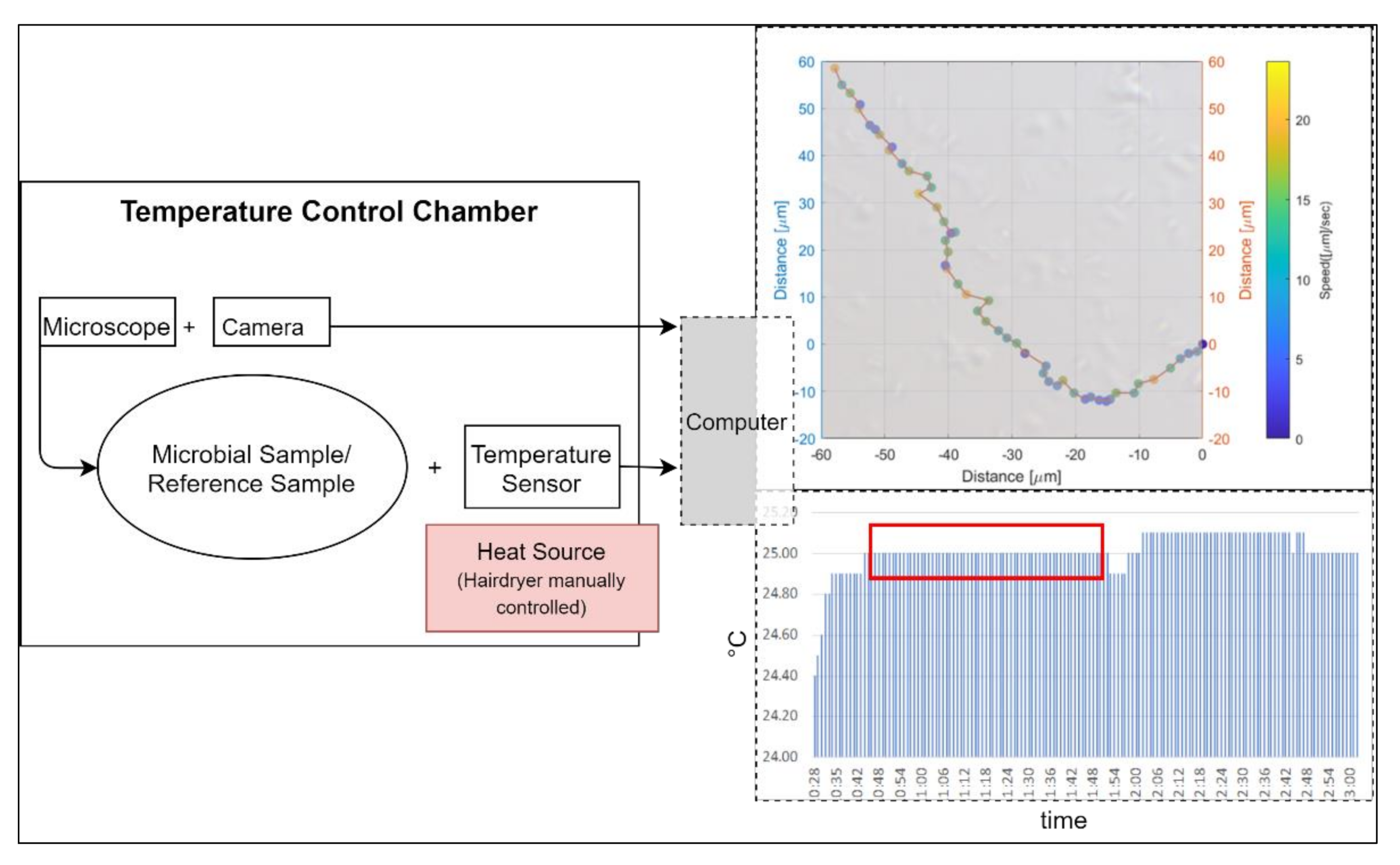
We used a Primo Star Full Köhler phase contrast microscope with a 40× objective for the observations. Connected with the microscope was an AxioCam 105 color camera, which transmitted the pictures live to a computer. The microscope in combination with the digital camera leads to a pixel scale of 0.11 µm restricted by the Abbe diffraction limit (approximately 0.4 µm). We used a Neubauer counting chamber with a depth of 10 µm for the observations of the microbial sample and of a milky solution as a reference sample. The microscopic measurements were performed at temperatures of 25 °C, 30 °C, 35 °C, and 40 °C. To maintain a stable temperature, we used a thermal insulating chamber consisting of 4 cm polystyrene walls and a 5 mm insulating aluminum layer. We used a DOSTMANN P700 Universal Precision Thermometer. The probe of the thermometer was attached to the slide of the microscope. The chamber enclosed the microscope and the camera. We used a Philips ThermoProtect 2100 W hairdryer for controlling the temperature in the chamber. The setup of the temperature control chamber can be seen in Figure 1.
Due to time constraints, we tracked 50 individual movement pathways over ten seconds for each species, which was approximately 9.4% of the total individual E. coli cells observed, 10% of the total individual P. halocryophilus cells, 12% of the total individual P. haloplanktis cells, and approximately 40% of the total individual B. subtilis cells. Exposure times varied between species (and therefore optimal lighting for best image quality). The obtained videos had between 6.94 and 5.41 frames per second (in each case, the shutter speed was fast enough that the microbes could be clearly seen in the individual images, without smearing out).
Video files (.tiff) were imported for tracking into the software Fiji/ImageJ. For this, we used the manual tracking plugin, for which we calibrated the time interval and the x/y calibration according to the parameters of the microscopic recordings. With the software, we obtained the speed and the x/y-coordinates of the microbes for each video frame by clicking on their center with the mouse pointer.
2.4. Errors
We obtained images of 2560 × 1920 pixels with our system. Due to the digital zoom in the Fiji software, we could determine the location of the microbes with a ±1 pixel (=0.11 µm) accuracy, which is, depending on the frames rate of the videos, equivalent to a ±0.62 µm/s accuracy for E. coli and B subtilis, ±0.73 µm/s for P. halocryophilus, and ±0.76 µm/s for P. haloplanktis. Another source of error in tracking is that we only tracked the microbes in two dimensions, even though they were in a 10 micrometer (about ten microbial body lengths) deep medium. This limitation applies to all observed microbes. Another potential error was that for observations of this kind, externally induced shifts by convection must be prevented. Therefore, we observed a reference sample (a milk solution in the motility medium) on the stage with the same configuration of heating source, thermal chamber walls, and microscope, and adjusted until no convective motion in the medium occurred before the start of each run.
2.5. Classes and Features of Data
We worked with aggregates (defined as the averaged properties of the observed cells over the length of the recordings) of features over the recorded time to avoid biases caused by methodological errors, such as variations of exposure time from recording to recording. At the same time, it is important to provide the tracking and classification algorithms only with information that does not give them a priori information about the observed species, such as by an increase in the occurrence of a specific species in a certain coordinate range. Therefore, no coordinate data was passed on to the algorithms.
Two classes were used to classify the cells and the abiotic particles with the supervised learning algorithms: The first class consisted of the four observed bacterial species at 25 °C, and the second class consisted of the simulated mobility traces due to Brownian motion at the same temperature. Subsequently, the cells were classified into four classes representing the used species.
The microbes and the simulated particles were tracked using individual motility/mobility traces over ten seconds. Depending on the species and exposure time, between 54 and 69 consecutive images were taken for each microbial pathway. Two consecutive images were needed to determine the speed and three consecutive images for the angle change (e.g., a ten-second image sequence of 54 consecutive images resulted in 53 speed information and 52 angle information units for each microbial pathway).
To ensure that the different exposure times do not provide the classifiers with a priori information, we determined the average values of the microbial pathway features over ten seconds. We intended to select properties that classify the motion as simple as possible to minimize the computational requirements. A too complex analysis of the motion behavior (including complex consecutive motion properties) would risk overfitting certain features that might work well for our data set but fail for some other data set. A general problem with classifications is that it is not known in advance what the most suitable features are for classification. In our case, this means: what are the motility features leading to the best accuracy of classifying microbial species, and how to classify best biotic vs. abiotic particles? The features we used were: (1) mean speed of the particle, (2) standard deviation of mean particle speed, (3) relative amount of clockwise directional change, (4) relative amount of counterclockwise directional change, (5) relative amount of low (<20°) directional change (6) average directional changing angle, (7) standard deviation of directional changing angles, (8) relative amount of low speed (set arbitrarily at < 1.4 µm/s and used as a tunable parameter), (9) mean distance of particles after ten seconds.
Measured angles during movement are taken from three consecutive coordinates based on the motility behavior of the microbes: The angles here are the angles that form the two sides and . 1 is the coordinate of the microbe in the first video frame, 2 in the following one, and 3 in the one thereafter (see Figure 2).
Besides the features (1), (2), (6), and (7) (mean speed and angles, respectively, and their standard deviations), which are the most apparent properties to quantify the microbial pathways, there are additional ways to quantify motility patterns. Bacteria moving near surfaces experience hydrodynamic forces that attract them towards the surface and cause them to move in circular trajectories [15]. Due to this phenomenon, we also quantified the direction of the feature with (3) and (4). In feature (5), we defined a directional change of <20° as a straight movement. The definition of low speed <1.4 µm/s is added as a feature (8). This is an empirical parameter determined after the observations were concluded, and it is suitable for classification because of its statistically significant variations between bacterial species. The speed of 1.4 µm/s equals approximately the mean displacement per second of a particle with a diameter of 0.5 µm at 25 °C. Feature (9) is a way to easily quantify the straightness of an object’s average movement over the observation period.
It is crucial to select features that contribute most to the predicted output. Irrelevant features in the data can decrease the accuracy of the classification. Feature selection means less redundant data and, therefore, less impact from noise, more accurate models, and faster training of algorithms. To remove irrelevant features, we implemented an exhaustive feature selector for sampling and evaluating all possible feature combinations to classify the species. Calculating all nine features from the observed coordinates of the respective frames required very little computer power.
We used 10-fold cross-validation for all algorithms, during which the dataset was shuffled randomly and split into ten groups. Each group was used once as a test group, with the remaining groups being used as training data. A model fitting the training set was evaluated in the test set. The result of the evaluation was kept. The model was then discarded, and the next group was used as a test group. Then, the quality of the algorithm was evaluated by the respective individual test results. The accuracy of the algorithm was then determined for all test groups. This increases the accuracy of estimating how the models are expected to perform with data not used during the training of the model. We used Python 3.7 in the scientific environment Spyder for running the algorithms. The classifiers were Logistic Regression Classifiers (LRC), Linear Discriminant Analysis (LDA), K-Nearest Neighbor Classifiers (KNN), Classification and Regression Trees (CART), Naïve Bayes (NB), and Support Vector Machines (SVM). For detailed information on these classifiers, see Supporting Information and [16].
2.6. Simulation of Brownian Motion
Einstein (1905) developed a statistical mechanics theory for Brownian motion [17]. It states that the mean displacement of a Brownian particle is proportional to the square root of the elapsed time. That is where the mean displacement squared is , the diffusivity is and the elapsed time is . The diffusivity is given by:
with (Boltzmann’s constant), being the temperature, and μ being the ratio of the particle’s drift speed to an applied force, which can be calculated by . is the dynamic viscosity of the fluid, and is the particle’s radius. This leads to:
Based on these equations, we simulated the movements of particles due to Brownian motion at the temperatures investigated with Matlab. We ran simulations with viscosities for water at 25 °C (η = 0.89 mPa*s) assuming spherical particles, with radii between 0.25 µm and 1 µm. The advantage of the simulations is that one can easily vary all relevant parameters. An additional benefit is that massive data sets can be quickly generated to train supervised learning procedures.
3. Results
3.1. Motility Parameters
In the simulations, the mean speed due to Brownian motion was 3.03 at 25 °C with a standard deviation of . The mean distance a particle traveled during ten seconds was 4 with a standard deviation of 2.5 , and the relative amount of low speed (<1.4 µm/s) was with a standard deviation of .
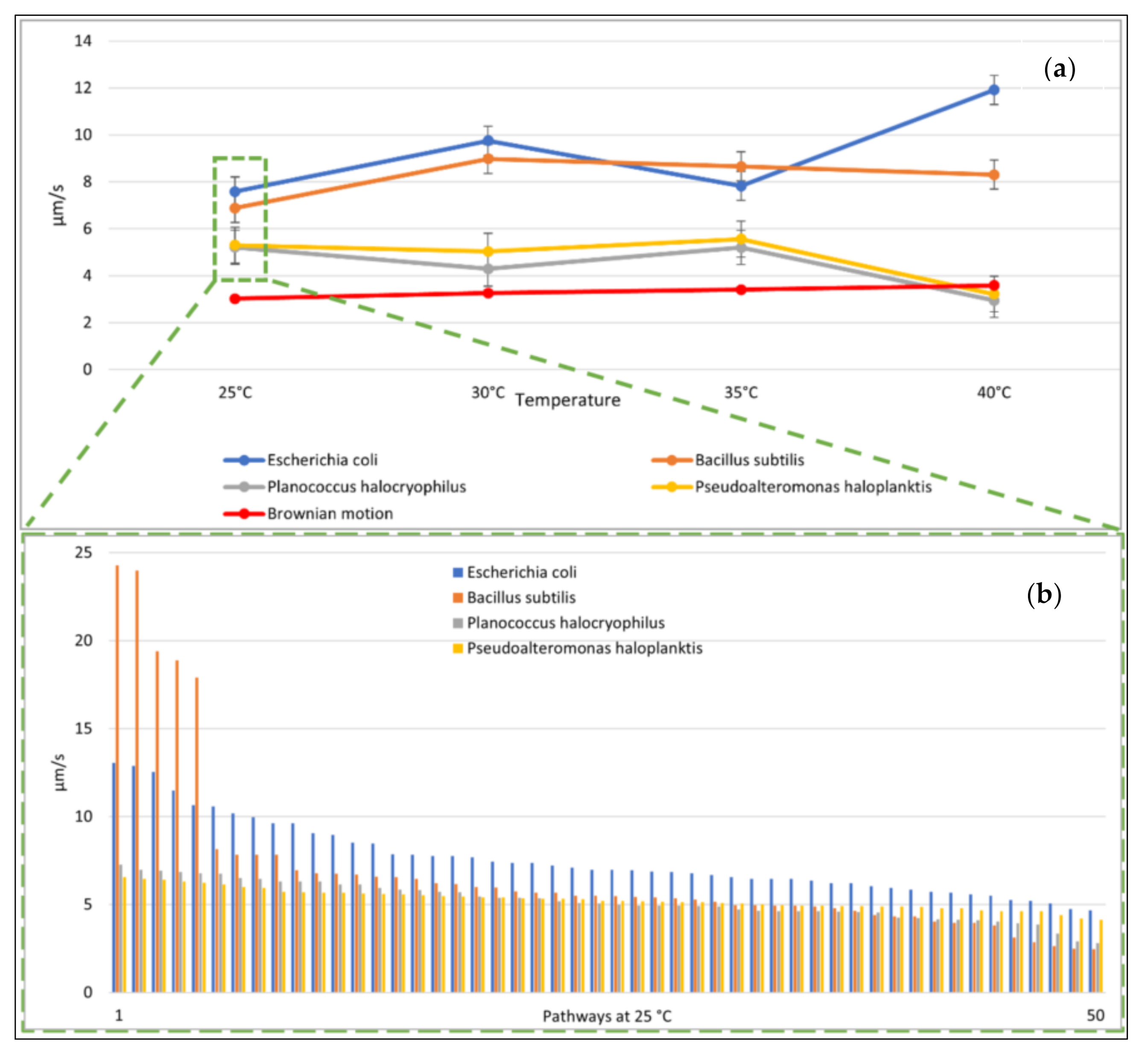
The observed mean speeds of the tracked bacteria varied with microbial species. E. coli was the fastest observed among all bacteria, with a mean speed of at 25 °C. The mean speed of P. haloplanktis was measured to be at 25 °C and P. halocryophilus had a mean speed of (Figure 3a and Supporting Information). B. subtilis were moving the fastest and had a mean speed of at 25 °C. (Figure 3b). At 25 °C, the movements due to (the simulated) Brownian motion were clearly slower than the microbial movements.
We heated the liquid water medium and observed the bacterial species additionally at 30 °C, 35 °C, and 40 °C. The change of mean speed with temperature was variable for the different species tested. E. coli was the only species that had its highest speed at 40 °C. All other bacteria tested had no consistent behavior with regard to an increase in temperature. However, as expected, (the simulated) Brownian motion increased with temperature. For P. halocryophilus and P. haloplanktis, we observed a similar relationship of mean speed as function of temperature. However, we noticed that both microbial species had speeds similar to (the simulated) Brownian motion at increased temperatures, which was likely the case because these elevated temperatures were out of their viability range, and the cells were not anymore self-propelled but dead (see Figure 3a).
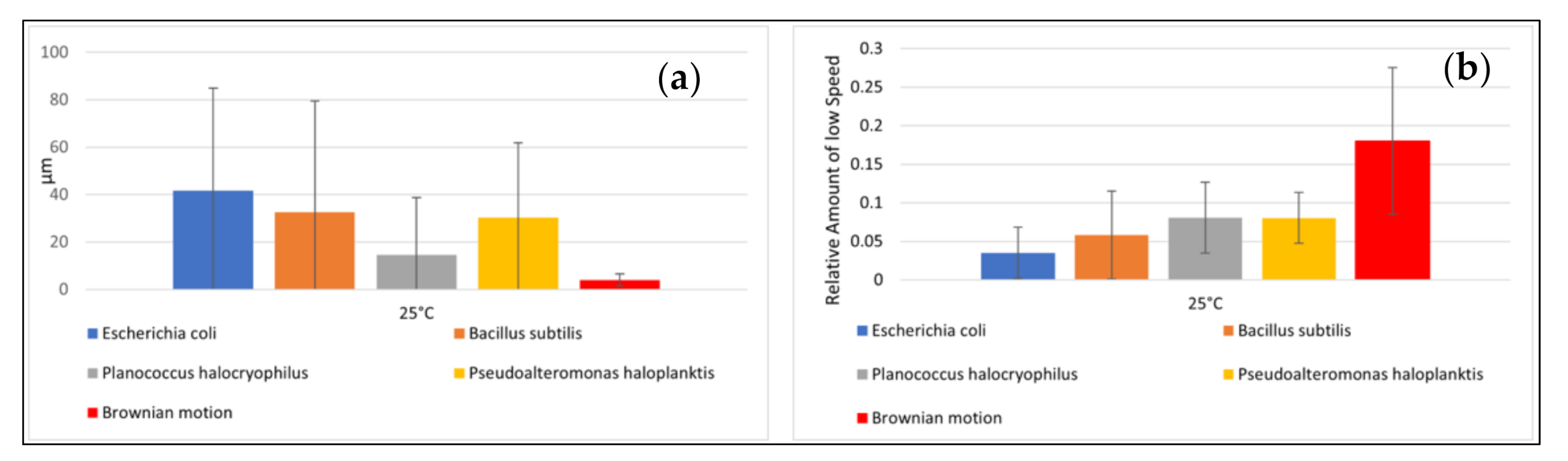
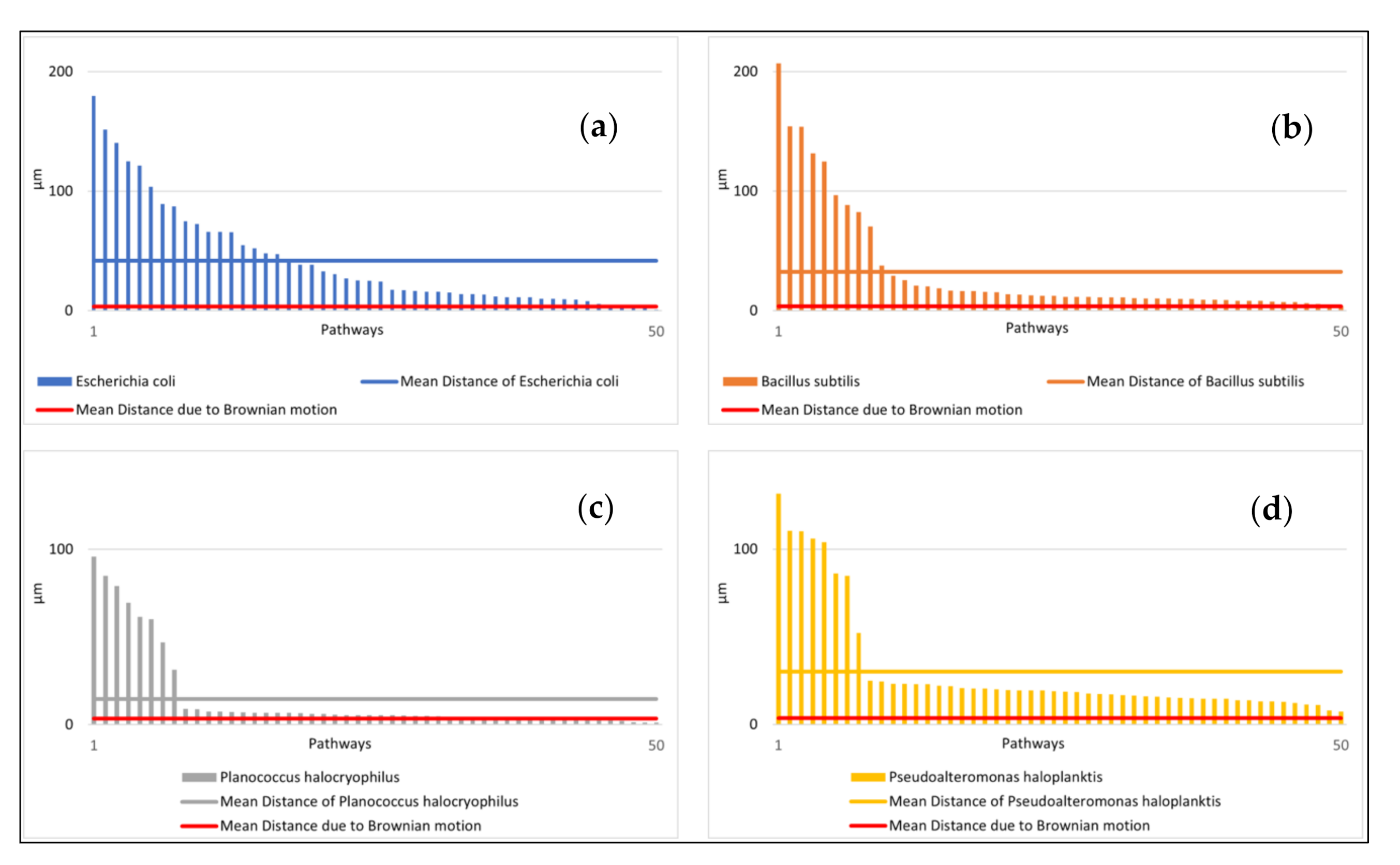
The speeds of the microbes indicated large standard deviations. The mean distance from the starting point after ten seconds and the relative amount of small speed also showed high standard deviations (Figure 4a,b). The distance from the starting point for each microbial pathway and the relative amount of small speed are shown in Figure 5 and Figure 6. Comparisons to normal distribution, histograms of the microbial speed distribution, and Q-Q plots are shown in the Supporting Information.
The mean distance that the microbes traveled after ten seconds at 25 °C correlated well with the respective mean speeds of the microbes tested (e.g., the Pearson correlation coefficient was 0.83). E. coli achieved a mean distance of 41.70 µm and B. subtilis a mean distance of 32.67 µm. P. halocryophilus advanced after ten seconds 14.59 µm, and P. haloplanktis 30.22 µm. P. haloplanktis and P. halocryophilus had approximately the same mean speed, but P. halocryophilus covered less distance than P. haloplanktis, which likely means it made more turns and stops during its movement.
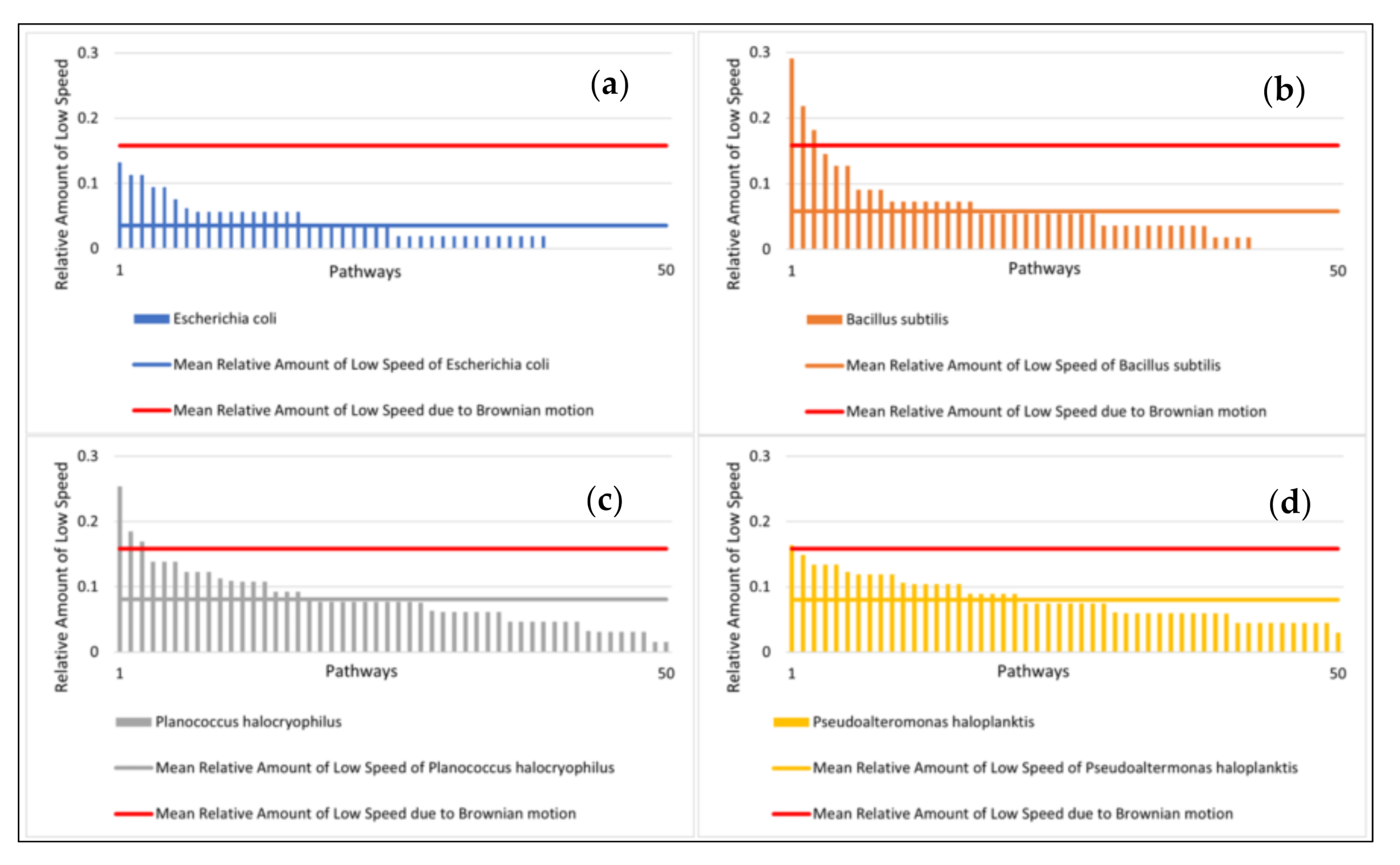
The mean relative amount of low speed (<1.4 µm/s) at 25 °C was highest for P. haloplanktis and P. halocryophilus with 8.0% and 8.1%, respectively. B. subtilis exhibited a mean relative amount of low speed of 5.8% and E. coli of 3.5% (Figure 6, mean values for the simulated pathways are also included in the figure).
Our observations of the features “standard deviation of particle speed”, “relative amount of low directional change (<20°)”, “average directional angle”, and “standard deviation of directional changing angle” did not show any substantial differences on the species level and thus were not substantial enough to be useful for the automated classification.
3.2. Results of Classification Algorithms
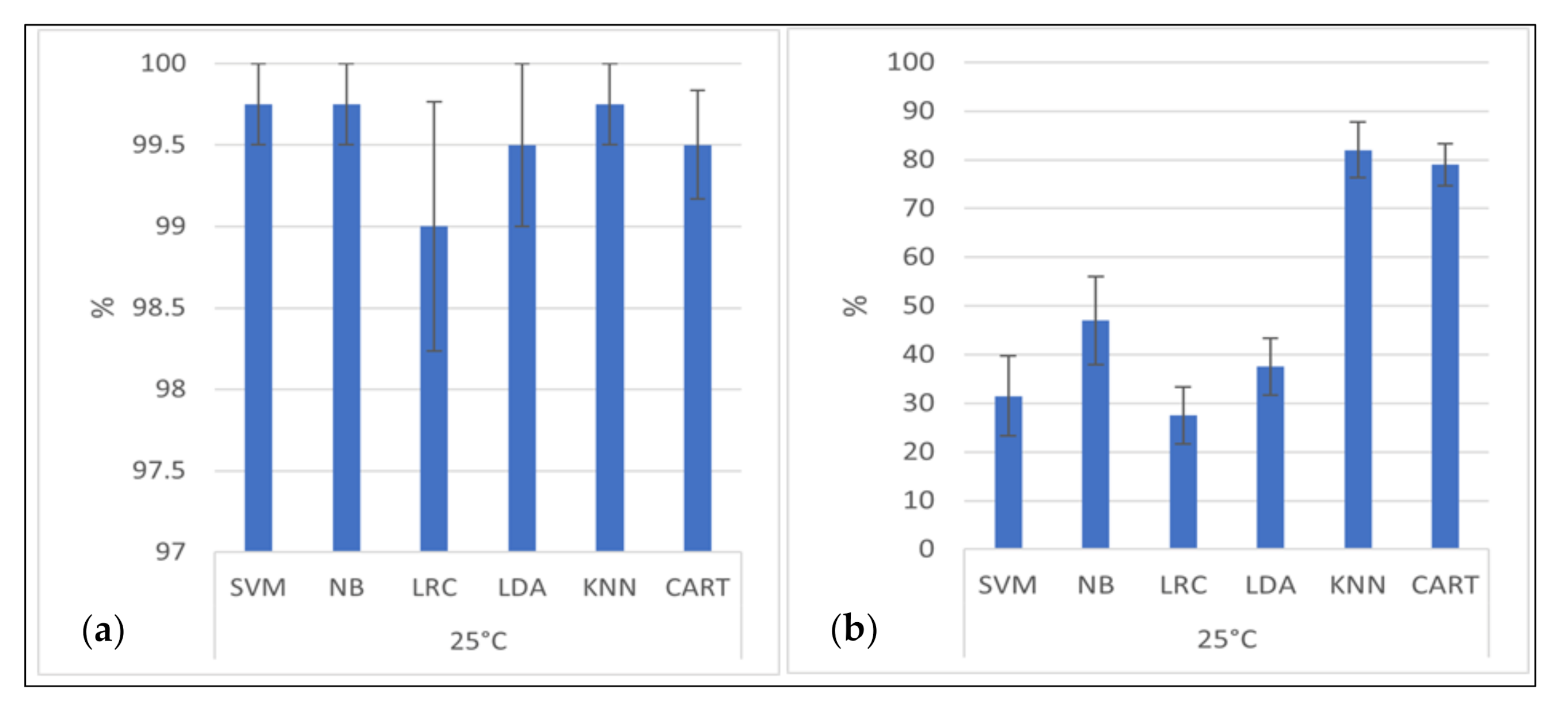
All classification algorithms were shown to have higher detection accuracy than 98% for distinguishing between observed bacteria and the simulated Brownian motion (Figure 7a). The best accuracy is that of the classification algorithm based on Support Vector Machines, Naïve Bayes, and K-Nearest Neighbor (with K = 1) (99.75% ± 0.25% classification accuracy). Six of the seven classifiers reveal the “mean distance after ten seconds” tracking parameter as the most successful classification scheme when only one parameter is used. Only the Linear Discriminant Analysis Classifier achieved a better classification accuracy with the feature mean speed.
The classification accuracies for the assignments of the four bacterial species are shown in Figure 7b, which highlight the usefulness of the algorithms with the optimum feature selection. The best classification accuracy has the K-Nearest Neighbor Classifier (KNN, with K = 1) with an accuracy of 82% ± 5.7%.
The optimal feature for the highest detection accuracy and the second-highest detection accuracy was the relative amount of low speed (Table 1). The KNN Classifier and the CART Classifier were not only with their optimal feature selection (relative amount of low speed) superior to the other algorithms but also with their respective second-, third-, and fourth best feature selection.
We note that especially for the algorithms with high classification accuracy, i.e., KNN and CART, only a few features were selected. The KNN Classifier (with K = 1, see Supporting Material) used for its most successful classification results the features relative amount of low speed, relative amount of clockwise directional change, relative amount of counterclockwise directional change, and relative amount of low directional change. The CART Classifier, which delivered the second-best classification results, used the features relative amount of low speed, average directional angle, and relative amount of counterclockwise directional change (Table 1).
4. Discussion and Conclusions
As shown in Figure 3, Figure 4, Figure 5 andFigure 6, there is a substantial amount of noisiness in the data. Our objective was to discriminate life from non-life using multiple independent criteria. The discrimination is challenging because it is difficult to apply a weighting procedure and motility is not easily traceable to basic physical principles. However, our results show that it is possible to differentiate between the movement of microbial cells and abiotic particle movements (simulated Brownian motion) with very high confidence and accuracy of > 99%. This indicates that Machine Learning is very helpful for this kind of data processing. All tested algorithms successfully distinguished biotic motions from the simulated movements of abiotic particles of similar size. The mean square displacement of particles due to Brownian motion was proportional to the elapsed time, which made the mean distance of particles traveled after ten seconds the most important feature for this purpose. However, it was challenging to distinguish between different bacterial species due to variations in the investigated features, which was observed for all four tested species. The distance advanced by the microbial species after ten seconds correlated only to a limited degree to their respective mean speeds. A feature that seems to be superior for the classification of the bacterial species was the relative amount of low speed (which is a tunable parameter and best classification results were achieved for speeds of <1.4 µm/s). This feature was particularly suitable for the classification of bacterial species when using the KNN or the CART algorithm.
All of the used features for the classification (biotic vs. abiotic, and species identification) are simple statistical values, which can be determined relatively easily automatically without high computational power through two-dimensional observations of the motility pathways. Thus, a quantitative analysis of the movement of microbes may also be a way to screen samples for certain specific microbes if sufficient quantitative data on microbial motility are available.
3D-microscopy approaches used to track microbes like digital holographic microscopy or defocused image methods are a trade-off between performance and ease of use [18,19]. Microbial motility occurs in nature in three dimensions, and a 3D observation most likely will increase the identification accuracy. Thus, it will be instructive to conduct similar experiments, but in 3D, in the future. Nevertheless, the classification with our more straightforward 2D approach with much less data processing still provided acceptable accuracies and results, especially when distinguishing microbial motility from Brownian motion of particles similar in size to microbes. A trade-off between technical requirements must be considered when designing space missions to search for life in extraterrestrial bodies such as Mars or Europa. The maximal data volume capability of NASA’s Europa-Lander for its microscope is 302 Mbit/s. Ten-second recordings of our observations have a data size of approximately 3600 Mbit. This shows that the transmission of longer recordings, which will undoubtedly be necessary, is impractical, at least with our current laboratory hardware. The output of all the processed data, however, has a data size of less than 9 Mbit. Thus, if the data could be processed onboard [20] rather than being transferred to ground control and analyzed on Earth, then this is a feasible way to conduct the motility analyses.
Liquid water is a necessary condition for the existence of life as we know it [21]. Thus, our experiments to expose microbes to liquid water as a medium is appropriate. However, during a space mission to Mars or the subsurface oceans of the icy satellites, this may pose an additional challenge because of the possible instability of the liquid water sample during microscopic observations. To observe the motility of particles (and possible microbes) within the sample, it is crucial that the system does not exhibit any movements induced by external forces or internal forces, for example, vibrations of the system due to movements of actuators on the lander. Additionally, it is conceivable that magnetic forces could interact with the sample, provided it contains ferromagnetic elements. Possible external electromagnetic forces or forces induced by components in the lander could also lead to unwanted movements in the sample. Therefore, the microscopic system must have sufficient shielding. Alternatively, it may be possible to subtract the effect of external motions from the microbial motility behavior. This seems to be a feasible approach for background laminar flow after more insights about the interaction of bacteria and fluid flow are gained because laminar flow equations are well known and understood. This will be more challenging for other external forces such as background turbulent flow or magnetic fields that fluctuate with time.
Thermal stability is one of the most crucial aspects that need to be considered when conducting a microscopic detection of motility. Under specific scenarios, for example, when a frozen sample is obtained from an icy moon, it may be necessary to heat the sample. The heating process is dynamic, and the Brownian motion of any liquid will accelerate during heating. Uneven heating of the solvent (and possible evaporation) will cause a flow that can affect any microbes in the fluid and their movements. Microbes that could be detected under “stable” thermal conditions could be non-distinguishable from inanimate matter when there is a bulk flow forced by heat transfer (convection).
Viscosity, temperature, and particle size are also important. To make quantitative comparisons to Brownian motion, accurate information is needed about the sample’s viscosity, the temperature, and the particle sizes observed. The latter can be done using a filter system that only allows certain particle sizes to pass into the liquid sample. Viscosity and temperature must be determined in-situ.
We have observed that microbes behave differently at different temperatures. This is also true for other stimuli (chemical, light). This characteristic behavior for each species could be exploited for a microscopic life detection device when combined with machine learning techniques. The variability of the environmental conditions, not only the temperature but also inducing chemical microbial stimuli, is worthy of further investigation. We expect that we might be able to observe putative microbes moving toward or away from the provided stimulus. Morphological observations may be an additional criterion of whether life is indeed present. The result of this further research may eventually allow the design of a motility-based instrument suitable as a life-detection device. Such a device might not only be suitable for space missions but also for microbial species identification on Earth, for example, to distinguish pathogenic from non-pathogenic microorganisms in a water sample.
Supplementary Materials
The following are available online at http://0-dx-doi-org.brum.beds.ac.uk/10.14279/depositonce-10638.3 and https://0-www-mdpi-com.brum.beds.ac.uk/2075-1729/11/1/44/s1. Supporting information for machine learning algorithms applied to identify microbial species by their motility: Discussion of the simulation of Brownian motion (Text S1, Figure S1), mean speed distribution (Figure S2), and used classifiers (Table S1). Data Set S1: Motility Data of the four species. Data Set S2: Aggregated motility data over a period of ten seconds for the four species. Data Set S3: Motility data for the microbes for Figure 3a and Figure 4a,b. Data Set S4: Aggregated motility data of microbes and aggregated motility data of simulated biotic movements. Data Set S5: Python code of the KNN Classifier. Data Set S6: Python code of the CART Classifier. Data Set S7: Python code of the LDA Classifier. Data Set S8: Python code of the LRC Classifier. Data Set S9: Python code of the NB Classifier. Data Set S10: Python code of the SVM Classifier. Data Set S11: Matlab file for the creation of movement due to Brownian motion and of its aggregated motility information. Data Set S12: Matlab file for the calculation of the motility information of the X/Y-information of the particle observations. Deployment of the aggregated motility information. Data Set S13: Detailed Information of classification results “biotic vs abiotic”. Data Set S14: Detailed Information of species classification results.
Author Contributions
Conceptualization, M.R. and D.S.-M.; methodology, M.R.; software, M.R.; validation, M.R., J.S., and D.S.-M.; formal analysis, M.R.; investigation, M.R.; resources, D.S.-M.; data curation, M.R. and J.S.; writing—original draft preparation, M.R.; writing—review and editing, M.R., J.S., and D.S.-M.; visualization, M.R.; supervision, D.S.-M.; project administration, M.R.; funding acquisition, M.R., J.S., and D.S.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a Proof-of-Concept Grant from the European Research Council (ERC - GAP 957272 PORTPATH). Max Riekeles acknowledges a Doctoral Scholarship from the Friedrich Ebert Foundation.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data, classification algorithms and results, and the result and codes of the simulations that support the findings of this study are available online at http://0-dx-doi-org.brum.beds.ac.uk/10.14279/depositonce-10638.3.
Acknowledgments
The authors would like to thank Nilton Rennó and Julian Zimmert for helpful conversations and feedback.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nadeau, J.L.; Bedrossian, M.; Lindensmith, C.A. Imaging technologies and strategies for detection of extant extraterrestrial microorganisms. Adv. Phys. X 2018, 3, 1424032. [Google Scholar] [CrossRef] [Green Version]
- Thomas-Keprta, K.L.; Clemett, S.J.; Bazylinski, D.A.; Kirschvink, J.L.; McKay, D.S.; Wentworth, S.J.; Vali, H.; Gibson, J.E.K.; Romanek, C.S. Magnetofossils from Ancient Mars: A robust biosignature in the Martian meteorite ALH84001. Appl. Environ. Microbiol. 2002, 68, 3663–3672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, R.D. Motility. J. Cell Biol. 1981, 91, 148s–155s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, M.T.; Brock, T.D.; Martinko, J.M. Brock Biology of Microorganisms, 14th ed.; Pearson: Boston, MA, USA, 2015; ISBN 0321897390. [Google Scholar]
- Mitchell, J.G.; Kogure, K. Bacterial motility: Links to the environment and a driving force for microbial physics. FEMS Microbiol. Ecol. 2006, 55, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, W.R.; Doetsch, R.N. Temperature effects on bacterial movement. Appl. Environ. Microbiol. 1977, 34, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, M.; Koirala, S.; Picton, D.; Strahl, H.; Hoskisson, P.A.; Rao, C.V.; Gillespie, C.S.; Aldridge, P.D. Growth rate control of flagellar assembly in Escherichia coli strain RP437. Sci. Rep. 2017, 7, 41189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dijl, J.M.; Hecker, M. Bacillus subtilis: From soil bacterium to super-secreting cell factory. Microb. Cell Fact. 2013, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sneath, P.H.A.; McGowan, V.; Skerman, V.B.D. Approved lists of bacterial names. Int. J. Syst. Evol. Microbiol. 1980, 30, 225–420. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Kearns, D.B. The structure and regulation of flagella in Bacillus subtilis. Annu. Rev. Genet. 2014, 48, 319–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zobell, C.E. A List of Marine Bacteria Including Descriptions of Sixty New Species; University of California Press: Berkley/Los Angeles, CA, USA, 1944. [Google Scholar]
- Wilmes, B.; Hartung, A.; Lalk, M.; Liebeke, M.; Schweder, T.; Neubauer, P. Fed-batch process for the psychrotolerant marine bacterium Pseudoalteromonas haloplanktis. Microb. Cell Fact. 2010, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mykytczuk, N.C.S.; Wilhelm, R.C.; Whyte, L.G. Planococcus halocryophilus sp. nov., an extreme sub-zero species from high Arctic permafrost. Int. J. Syst. Evol. Microbiol. 2012, 62, 1937–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mykytczuk, N.C.S.; Foote, S.J.; Omelon, C.R.; Southam, G.; Greer, C.W.; Whyte, L.G. Bacterial growth at −15 °C; molecular insights from the permafrost bacterium Planococcus halocryophilus Or1. ISME J. 2013, 7, 1211–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauga, E.; DiLuzio, W.R.; Whitesides, G.M.; Stone, H.A. Swimming in circles: Motion of bacteria near solid boundaries. Biophys. J. 2006, 90, 400–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; With Applications in R; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7137-0. [Google Scholar]
- Einstein, A. Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen. Ann. Phys. 1905, 322, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Lindensmith, C.A.; Rider, S.; Bedrossian, M.; Wallace, J.K.; Serabyn, E.; Showalter, G.M.; Deming, J.W.; Nadeau, J.L. A submersible, off-axis holographic microscope for detection of microbial motility and morphology in aqueous and icy environments. PLoS ONE 2016, 11, e0147700. [Google Scholar] [CrossRef] [PubMed]
- Taute, K.M.; Gude, S.; Tans, S.J.; Shimizu, T.S. High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat. Commun. 2015, 6, 8776. [Google Scholar] [CrossRef] [PubMed]
- Europa Lander Study 2016 Report: Europa Lander Mission. JPL D-97667. Available online: https://solarsystem.nasa.gov/docs/Europa_Lander_SDT_Report_2016.pdf (accessed on 5 January 2021).
- Schulze-Makuch, D.; Irwin, L.N. Life in the Universe: Expectations and Constraints, 3rd ed.; Springer International Publishing: Cham, Switzeland, 2018; ISBN 978-3-319-97658-7. [Google Scholar]
Figure 1.
Setup of temperature control chamber: left side shows a schematic view, while the upper right image shows an example pathway of an Escherichia coli bacterium during a trial run at 25 °C. The lower right indicates the stability of the temperature for the same run over a time duration of 64 s (red box).
Figure 1.
Setup of temperature control chamber: left side shows a schematic view, while the upper right image shows an example pathway of an Escherichia coli bacterium during a trial run at 25 °C. The lower right indicates the stability of the temperature for the same run over a time duration of 64 s (red box).

Figure 2.
Sketch of angles of a microbe from three consecutive coordinates during motility behavior.
Figure 2.
Sketch of angles of a microbe from three consecutive coordinates during motility behavior.

Figure 3.
Mean microbial speed. (a) (top): Variations in mean microbial speed and particle mean speed due to Brownian motion as function of temperature with tracking accuracy. Note: The movement of the Brownian motion was simulated. (b) (bottom): Mean speed of each microbial pathway at 25 °C (sorted from fastest to slowest).
Figure 3.
Mean microbial speed. (a) (top): Variations in mean microbial speed and particle mean speed due to Brownian motion as function of temperature with tracking accuracy. Note: The movement of the Brownian motion was simulated. (b) (bottom): Mean speed of each microbial pathway at 25 °C (sorted from fastest to slowest).

Figure 4.
Motility data. (a) (left): Mean distance of microbial pathways and Brownian motion after ten seconds with standard deviation. (b) (right): Relative amount of low speed with standard deviation. Note: Brownian motion was simulated.
Figure 4.
Motility data. (a) (left): Mean distance of microbial pathways and Brownian motion after ten seconds with standard deviation. (b) (right): Relative amount of low speed with standard deviation. Note: Brownian motion was simulated.

Figure 5.
Distances of microbial pathways from their respective starting point after ten seconds at 25 °C (sorted from largest to smallest) in comparison to the mean distance due to Brownian motion. (a) (top left): Distances of the pathways of E. coli. (b) (top right), B. subtilis. (c) (bottom left): P. halocryophilus. (d) (bottom right): P. haloplanktis. Note: The movement of the Brownian motion was simulated. Its mean distance after ten seconds is shown in red.
Figure 5.
Distances of microbial pathways from their respective starting point after ten seconds at 25 °C (sorted from largest to smallest) in comparison to the mean distance due to Brownian motion. (a) (top left): Distances of the pathways of E. coli. (b) (top right), B. subtilis. (c) (bottom left): P. halocryophilus. (d) (bottom right): P. haloplanktis. Note: The movement of the Brownian motion was simulated. Its mean distance after ten seconds is shown in red.

Figure 6.
Relative amount of low speed at 25 °C with mean values (sorted from highest to lowest). (a) (top left): Relative amount of low speed of E. coli; (b) (top right): B. subtilis; (c) (bottom left): P. halocryophilus; and (d) (bottom right): P. haloplanktis. Note: The movement of Brownian motion was simulated. Its mean relative amount of low speed is shown in red. The feature low speed was here defined as speeds < 1.4 µm/s.
Figure 6.
Relative amount of low speed at 25 °C with mean values (sorted from highest to lowest). (a) (top left): Relative amount of low speed of E. coli; (b) (top right): B. subtilis; (c) (bottom left): P. halocryophilus; and (d) (bottom right): P. haloplanktis. Note: The movement of Brownian motion was simulated. Its mean relative amount of low speed is shown in red. The feature low speed was here defined as speeds < 1.4 µm/s.

Figure 7.
Classification results. (a) (left): Accuracy of the classification of biotic and abiotic particles with standard error. (b) (right): Accuracy of species identification with standard error. SVM is Support Vector Machines Classifier, NB is Naïve Bayes Classifier, LRC is Logistic Regression Classifier, LDA is Linear Discriminant Analysis Classifier, KNN is K-Nearest Neighbor Classifier, CART is Classification and Regression Tree Classifier.
Figure 7.
Classification results. (a) (left): Accuracy of the classification of biotic and abiotic particles with standard error. (b) (right): Accuracy of species identification with standard error. SVM is Support Vector Machines Classifier, NB is Naïve Bayes Classifier, LRC is Logistic Regression Classifier, LDA is Linear Discriminant Analysis Classifier, KNN is K-Nearest Neighbor Classifier, CART is Classification and Regression Tree Classifier.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Algorithm performance vs. features used. Best performing feature selection for each algorithm shown in dark grey background. The KNN and CART Classifiers were found to be superior to the other algorithms, even with a non-optimal feature selection. SVM is Support Vector Machines Classifier, NB is Naïve Bayes Classifier, LRC is Logistic Regression Classifier, LDA is Linear Discriminant Analysis Classifier, KNN is K-Nearest Neighbor Classifier, CART is Classification and Regression Tree Classifier.
Table 1.
Algorithm performance vs. features used. Best performing feature selection for each algorithm shown in dark grey background. The KNN and CART Classifiers were found to be superior to the other algorithms, even with a non-optimal feature selection. SVM is Support Vector Machines Classifier, NB is Naïve Bayes Classifier, LRC is Logistic Regression Classifier, LDA is Linear Discriminant Analysis Classifier, KNN is K-Nearest Neighbor Classifier, CART is Classification and Regression Tree Classifier.
| Classifier | Mean Speed of the Particle | Standard Deviation of Mean Particle Speed | Relative Amount of Clockwise Directional Change | Relative Amount of Counterclockwise Directional Change | Relative Amount of Low Directional Change | Average Directional Angle | Standard Deviation of Directional Changing Angles | Relative Amount of Low Speed | Mean Distance of Particles after Ten Seconds | Accuracy |
|---|---|---|---|---|---|---|---|---|---|---|
| KNN | ✓ | |||||||||
| CART | ✓ | |||||||||
| CART | ✓ | |||||||||
| CART | ✓ | ✓ | ||||||||
| CART | ✓ | ✓ | ||||||||
| KNN | ✓ | |||||||||
| KNN | ✓ | |||||||||
| KNN | ✓ | |||||||||
| NB | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| LDA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||
| SVM | ✓ | ✓ | ||||||||
| LRC | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Riekeles, M.; Schirmack, J.; Schulze-Makuch, D. Machine Learning Algorithms Applied to Identify Microbial Species by Their Motility. Life 2021, 11, 44. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010044
AMA Style
Riekeles M, Schirmack J, Schulze-Makuch D. Machine Learning Algorithms Applied to Identify Microbial Species by Their Motility. Life. 2021; 11(1):44. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010044
Chicago/Turabian StyleRiekeles, Max, Janosch Schirmack, and Dirk Schulze-Makuch. 2021. "Machine Learning Algorithms Applied to Identify Microbial Species by Their Motility" Life 11, no. 1: 44. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010044
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.