Extracellular Vesicles in Viral Pathogenesis: A Case of Dr. Jekyll and Mr. Hyde



1
Institute of Biomedical Systems and Botechnologies, Peter the Great St. Petersburg Polytechnic University, 194021 St. Petersburg, Russia
2
Department of Molecular Biology of Viruses, Smorodintsev Research Institute of Influenza, Ministry of Health of the Russian Federation, 197376 St. Petersburg, Russia
*
Author to whom correspondence should be addressed.
Life 2021, 11(1), 45; https://0-doi-org.brum.beds.ac.uk/10.3390/life11010045
Submission received: 26 November 2020
/
Revised: 4 January 2021
/
Accepted: 11 January 2021
/
Published: 13 January 2021
(This article belongs to the Section Physiology and Pathology)
Abstract
:Secretion of extracellular vesicles (EVs) is a fundamental property of living cells. EVs are known to transfer biological signals between cells and thus regulate the functional state of recipient cells. Such vesicles mediate the intercellular transport of many biologically active molecules (proteins, nucleic acids, specific lipids) and participate in regulation of key physiological processes. In addition, EVs are involved in the pathogenesis of multiple diseases: infectious, neurodegenerative, and oncological. The current EV classification into microvesicles, apoptotic bodies, and exosomes is based on their size, pathways of cellular biogenesis, and molecular composition. This review is focused on analysis of the role of EVs (mainly exosomes) in the pathogenesis of viral infection. We briefly characterize the biogenesis and molecular composition of various EV types. Then, we consider EV-mediated pro- and anti-viral mechanisms. EV secretion by infected cells can be an important factor of virus spread in target cell populations, or a protective factor limiting viral invasion. The data discussed in this review, on the effect of EV secretion by infected cells on processes in neighboring cells and on immune cells, are of high significance in the search for new therapeutic approaches and for design of new generations of vaccines.
1. Introduction
Intense intercellular communication, providing functional integration of cells and tissues in the organism, is the main attribute of multicellular life forms. In recent decades, the rapid progress of high-performance –omics technologies in biology has led to a secretome concept, as a sum of biologically active substances, of various natures, secreted by the cell, including soluble molecules, various granules, and extracellular vesicles. Secretome components circulate in the local intercellular medium or they can be transported by biological fluids to recipient cells throughout the organism. The cellular secretome determines the functional tuning of other cells in the organism [1] and also plays a key role in the pathogenesis of many diseases, among them infectious diseases. Besides molecules, significant portions of the cell secretome are more complex components, including various nanoparticles and several types of extracellular vesicles [2]. The phenomenon of membrane vesicle secretion by cells was first discovered more than three decades ago [3]. Extracellular vesicles were found to be secreted by all types of cells, and they were detected in various biological fluids: urine [4], blood [5], saliva [6], breast milk [7], bronchoalveolar fluid [8], amniotic fluid [9], and others.
Different types of extracellular vesicles (EVs) have certain degrees of similarity in physical and morphological characteristics (size, shape, buoyant density). Therefore, for classification of EVs, a complex of attributes is used, including pathways of biogenesis, size, and molecular composition [10]. The molecular composition of EVs depends on the type and functional state of the cells secreting them, and there are mechanisms that allow certain cellular proteins, lipids, and nucleic acids to be selectively transported into various types of vesicles. Currently, there are three databases accumulating information on the molecular composition of extracellular vesicles: EVpedia [11]; Vesiculepedia [12]; and Exocarta [13].
Over the past two decades, the role of EVs has been shown in several important biological processes such as immunosuppression during trophoblast implantation and placenta formation [14]; antigen presentation [15]; and proinflammatory cytokine secretion [16]. EVs play a key role in the pathogenesis of many diseases, among them oncological diseases [17,18,19], amyloidosis, and prion infection [20,21]. Therefore, assessment of changes in molecular EV composition (particularly, exosomal microRNA composition) can be a biomarker for various pathological states of cells and potentially an important diagnostic and prognostic tool in medicine.
Extracellular vesicles (especially exosomes) have been confirmed to be actively involved in the pathogenesis of various viral diseases [22,23,24]. Studies of the composition of EVs secreted by virus-infected cells has revealed that EVs can specifically pack and transport not only cellular proteins and nucleic acids, but also viral components and even whole viral particles [25,26]. The transfer of specific functional proteins, mRNAs, microRNAs, and other biologically active molecules to uninfected cells can lead to significant changes in cellular processes. This can either promote further virus invasion or, conversely, impede infection by activating anti-viral defense mechanisms [27,28].
In this review, we analyze the role of extracellular vesicles in viral invasion and pathogenesis, and consider the data on EV-mediated regulation of antiviral immune response in a number of viral infections.
2. Biogenesis and Molecular Composition of Extracellular Vesicles
Initially, EVs were assigned the function of waste management: an effective and safe way for cells way to eliminate unwanted components, an alternative to their intracellular enzymatic degradation [29]. Subsequently, it was shown that EVs can perform more complex and specialized functions related to intercellular communication and signaling. The regulatory and communicative functions of EVs are realized through the intercellular transfer of numerous biologically active molecules: various types of nucleic acid, proteins, and lipids. The EV membrane acts as an effective protector from enzymatic degradation of these molecules in the extracellular medium. EV-transferred molecules impart new functional properties to cells by activating various signaling pathways and regulating gene expression.
Formation of different types of extracellular vesicles involves various cellular mechanisms. Biogenesis of exosomes (typical size 40–100 nm) is associated with the endosomal membrane compartment. Membranes of the late endosomes form secondary invaginations, and multivesicular bodies with multiple intraluminal vesicles are then generated [30]. Multivesicular bodies can subsequently merge with lysosomes, with degradation of their contents [31]. Alternatively, they can merge with the plasma membrane and secrete enclosed intraluminal vesicles as exosomes [32]. The most important molecular component of exosome formation is the endosomal sorting complex required for transport (ESCRT). It consists of four multi-protein complexes: ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III [33]. ESCRT-0 is responsible for the clustering of proteins, which are ubiquitinylated during sorting and enter the intraluminal vesicles for further secretion in exosomes [34]. Protein TSG101 (tumor susceptibility gene 101) is a component of ESCRT-I that forms a complex with ubiquitinylated proteins and participates in the ESCRT-II activation, causing the formation of secondary invaginations in late endosomes [35]. ESCRT-II mediates the movement of ubiquitin-labeled proteins into intraluminal vesicles, acting in cooperation with ubiquitin-removing enzymes. The final stage of multivisicular body formation is the budding off of intraluminal vesicles, with the participation of ESCRT-III [36,37,38]. It is assumed that there is also an ESCRT-independent pathway of exosome formation, in which ceramides [39,40], tetraspanins, and heat shock proteins [41,42] play a crucial role.
Exosomes contain multiple specific proteins, nucleic acids, and lipids. According to data of high-throughput proteomic and transcriptomic analysis, over 2600 proteins, more than 900 mRNA, and more than 270 microRNAs are found in exosomes [43].
Unlike exosomes, another type of EVs, microvesicles, are directly budded from the plasma membrane [44]. On average, they are larger than exosomes (about 100–1000 nm). Components of future microvesicles are locally accumulated on the inner membrane side, then the membrane curves, forming a vesicle. This process is accompanied by specific translocase-mediated transfer of phosphatidylserine to the outer plasma membrane layer and contraction of the submembrane actin cytoskeleton [45,46]. Components of the ESCRT complex (TSG101 protein) are apparently involved in the formation of microvesicles as well [47].
Apoptotic bodies have a diameter of 500–5000 nm and contain highly compacted contents from the nucleus and cytoplasm. They bud off from the membrane during programmed cell death [48,49,50] and subsequently are captured by phagocytic cells (mainly macrophages) [51]. Briefly, the main molecules of extracellular vesicles are summarized in the Table 1.
The scheme of EV biogenesis and the main molecular components of exosomes, as the most fully characterized type of EV, are shown in Figure 1.
Interaction between EVs and target cells can occur by two different ways. First, EV membrane proteins can bind to receptor molecules on the surface of target cells, which are coupled to their signaling cascades. EV membrane proteins also can be cleaved by proteases and convert into soluble fragments, which act as ligands for corresponding cell receptors. Second, EV can fuse with the membrane of the target cells, with transfer of their cargo (protein, RNA) to the recipient cell [76].
3. Pro-Viral Role of Extracellular Vesicles in the Infectious Process
An objective analysis of EV effects on progression of viral infection meets certain methodological difficulties. Extracellular vesicles secreted by infected cells have very similar physical characteristics (size, buoyant density) to the majority of viruses. This makes the separation of viral particles and extracellular vesicles a difficult goal and complicates subsequent analytical studies [77]. Use of known protein markers to separate EV and viral particles is not always effective, because in reality not all vesicular structures can carry these markers on their surface [12]. On the other hand, various molecules of viral origin are selectively included into EVs, and this brings an additional complexity into EV separation and molecular analysis [78,79]. EVs of different origins were shown to carry a range of genetic material such as DNA fragments [80,81], mitochondrial DNA [82,83], and a great variety of RNAs including mRNAs, miRNAs, and small non-coding RNAs [84]. This ability makes EVs an important part of intercellular communication during virus infection. Transfer of nucleic acids derived by EVs from infected cells to the recipient could leads to proviral and antiviral effects as well [85].
Moreover, many viruses use pathways similar to EV pathways of biogenesis and secretion. This allows viruses to assemble with maximal efficiency in cells, with potent spreading to neighboring cells. In particular, many viruses use ESCRT and Rab GTPases for their secretion, namely, rhabdoviruses, filoviruses, arenaviruses, paramyxoviruses, herpesviruses, and hepatitis viruses A and C [86,87,88,89,90].
Extracellular vesicles are able to interact with the recipient cell via both receptor-dependent and receptor-independent pathways. Therefore, some viruses use EVs for their own invasion to expand the range of cells available for infection, which in general contributes to more efficient spread [91]. Overall, the role of EVs in viral pathogenesis is similar to that in tumor metastasis. Exosomes and microvesicles prepare the microenvironment for the further viral invasion.
Today the role of exosomes and other extracellular vesicles in the intercellular communication during cell infection is the most studied in the context of herpes simplex virus (HSV), Epstein-Bar virus (EBV), hepatitis viruses, and human immunodeficiency virus (HIV). Each of these viruses has unique characteristics and enhancing approaches of replication and spread. Virus adaptive and protecting systems are unique and effective as well. The use of extracellular vesicles containing specific cellular and viral components is one of the approaches implemented by viruses to achieve effective spread and infection of cells. A brief description of the main components of extracellular vesicles during different infections of viruses are presented in Table 2.
Below, we consider recent data on the molecular mechanisms of pro-viral exploitation of extracellular vesicles by a number of viruses investigated in this regard.
3.1. Herpes Simplex Virus (HSV-1)
HSV-1-infected cells secrete vesicular structures known as L-particles, which contain multiple cellular factors and viral envelope proteins (ICP0, ICP4) [112,113]. L-particles are comparable to exosomes in size; they are formed from the inner membranes of cells and are able to deliver their contents to neighboring non-infected cells [114]. The interaction of secreted L-particles with surrounding cells contributes to further viral spread and suppresses the immune response [115]. Though L-particles are non-infectious, they prepare cells for subsequent infection by activating the transcription of certain cellular proteins beneficial for subsequent HSV reproduction [116,117]. Moreover, the interaction of L-particles with mature dendritic cells reduces the expression of CD83 in them and ultimately contributes to immunosuppression [118].
In addition, HSV-1-infected cells secrete exosomes carrying viral glycoprotein B, which targets the MHCII synthesis pathway by antigen-presenting cells (APCs) [119]. In the cytoplasm of APCs, glycoprotein B binds to the HLA-DR protein receptor. This binding hinders HLA-DR receptor export to the cell surface. Thus, glycoprotein B reduces the number of HLA-DR receptors presenting viral determinants on the surface of APCs [120]. This also helps HSV-1 to avoid the immune response.
HSV-1 virions can be entirely included in microvesicles secreted by the infected cells, and later virions are transferred to neighbor cells. This allows the virus to expand the range of cells available for infection, as well as to avoid a targeted immune response [121]. The process of incorporation of HSV-1 virions into microvesicles is presumably associated with autophagy, since the autophagosome marker LC3-II has been found in microvesicles with viral particles [25].
3.2. Epstein-Barr Virus (EBV)
The gamma herpes virus EBV was the first oncogenic virus to be characterized [122], and its main oncogene is the membrane protein LMP1 [123,124]. This protein is selectively incorporated into exosomes secreted by EBV-infected cells [125]. To enter the exosomes, LMP1 interacts with tetraspanin CD63 [95,96] or is associated with lipid rafts [126]. Within exosomes, LMP1 is transferred to uninfected cells. LMP1-enriched exosomes also contain the viral glycoprotein gp350, which interacts with the CD21 receptor of B cells for exosome targeting [94]. Exosomal delivery of LMP1 to B cells causes their transformation [127]. LMP1 also induces massive apoptotic death of CD4+ T cells and natural killer cells [128].
Exosomes of EBV-infected cells selectively accumulate certain cellular proteins, such as galestine 9, which has a suppressive effect on Th1 T lymphocyte function [94,95]. In addition, exosomes deliver to uninfected cells a set of activated signaling proteins: epidermal growth factor receptor (EGFR); phosphatidylinositol-3-kinase (PI3K); fibroblast growth factor (FGF2); and HIF1-alpha [96,97,98,99,100,101]. These proteins trigger pathways responsible for recipient cell proliferation and tumor angiogenesis.
Exosomes of EBV-infected cells selectively include a variety of functional viral miRNAs and transfer them to recipient cells. More than 300 viral miRNAs (this is 20–25% of the entire fraction of cellular miRNAs) were detected in EBV-infected cells using NGS [129]. Numerous viral miRNAs, from all three known clusters (BHRF1, BART1, and BART2), were also identified during exosome profiling of EBV-infected cells [102,130]. Viral miRNAs are known to perform post-transcriptional multifunctional control, they can regulate infection pathogenesis (switching from lytic to persistent), induce changes in cellular metabolism beneficial to the virus, modulate the cellular life cycle, and evade host immune system action [131].
In particular, BHRF1 represses the IFN-inducible T-cell attracting chemokine CXCL11 [132], and miR-BART15 regulates NLRP3 inflammasomes and IL-1β production [133]. In addition, these miRNAs promote apoptosis in recipient cells by silencing the mRNA of apoptosis inhibitors such as BRUCE and TAX1BP1 [134,135]. Suppression of cytokine production, and the induction of apoptosis in immune recipient cells, contributes to the suppression of antiviral reactions against EBV. Thus, the exosomes of infected cells mediate immunosuppression and carry several signaling molecules (proteins and miRNAs) that create a favorable microenvironment for the progression of EBV-associated tumors, including Hodgkin’s lymphoma [136] and nasopharyngeal carcinoma [137].
Moreover, EVs secreted from EBV-infected cells contain coding and non-coding RNAs. Protein coding latent phase mRNAs (LMP1, LMP2, EBNA1, and EBNA2 gene transcripts) were detected in the exosomes from EBV-infected cells [138]. However, it is still unclear if these gene transcripts have biological activity after vesicular delivery into uninfected cells. EBV-encoded small RNAs (EBER1 and EBER2) are shown to be released from EBV-infected cells. They are suspected of supporting the survival and carcinogenesis of infected cells by avoiding cell apoptosis via signaling from TLR3 [139].
3.3. Human Immunodeficiency Virus (HIV)
For HIV, the “Trojan horse hypothesis” was formulated, postulating a hijacking, by HIV, of the pathways of exosome biogenesis and secretion to assemble and to bind the target cell without Env viral protein participation [140]. High similarity in lipid composition (high cholesterol and glycosphingolipid content) and protein composition (tetraspanins, GPI proteins, some cytoplasmic proteins) of exosomes and retroviruses was observed [141]. HIV also uses ESCRT complex components for self-assembly and release. Inhibition of exosome secretion in infected cells leads to a partial decrease in the virion release of HIV progeny [142,143]. However, the prevailing route of HIV progeny release is budding from the plasma membrane (similar to microvesicles) in regions with a high Gag protein level [144,145,146]. Therefore, the detailed molecular mechanism of the contribution of the EV biogenesis pathway to HIV replication remains unclear.
The main factor of CD4+ T cell apoptosis during HIV infection is the viral protein Nef. There is clear evidence that it is exosomes that mediate intercellular Nef transport to uninfected T cells, with their subsequent apoptotic death [105,147]. The Nef uptake by exosomes is based on their interaction with mortalin (Hsp70 family member) [148]. Nef-containing vesicles can also be captured by B cells, contributing to decreased IgG2 and IgA production and suppression of humoral immune response [149]. In addition, Nef enhances the production of multivesicular bodies and EV secretion [149,150,151], thereby enhancing the exosomal spread of this virulence factor. EVs can also transfer the HIV coreceptors CCR5 and CXCR4 to uninfected cells, which do not yet have those proteins. This process significantly facilitates further spread of infection [106,107]. Other viral components transported into EVs from HIV-infected cells are Gag protein (its impact on infectious spread is still unclear) [103] and gp120 envelope protein, which seemed to increase significantly the viral infectivity in human lymphoid tissues [104].
Moreover, exosomes secreted from HIV-infected cells are capable of carrying RNA, in particular HIV transactivation response element (TAR), which also increases the susceptibility of cells to viral infection [108]. The hairpin (stem-loop structure) at the 5’-end of TAR RNA binds to the Tat protein (trans-activator of transcription) in infected cells, enhancing viral RNA transcription [152].
3.4. Hepatitis C Virus (HCV) and Hepatitis A (HAV)
Hepatitis C virions have a size about 50 nm and are assembled in the cytoplasm of infected cells with the participation of components of exosome biogenesis [153]. EVs of infected cells contain viral E1 and E2 proteins [109] and viral RNA. The exosome marker CD81 is associated with viral proteins, which indicates a possible CD81 role in the packaging of viral proteins into exosomes [154]. Vesicles carrying the E2-CD81 complex increase the infectivity of the hepatitis C virus (HCV). EVs from infected cells can also contain whole virions [155]. Delivery within extracellular vesicles protects the virus from attack by neutralizing antibodies, thus contributing to the evasion of the immune system [156]. It also allows HCV to enter an increasing variety of potential host cells (other than hepatocytes) upon receptor-independent fusion of EVs with the cell membrane. Exosomes from infected cells are also capable of transferring viral RNA in complex with miR-122, Ago2, and HSP90, ensuring more efficient viral replication [110].
In contrast to HCV, the hepatitis A virus (HAV) is non-enveloped, but it is able to be incorporated into EVs for secretion and spread as well due to small virion size. Virion packaging into the vesicles is a result of interaction between the structural viral protein pX and the Alix protein [111]. The use of the cellular ESCRT complex allows the viral particles to be released into the extracellular space without rupture of the cell. Thus, non-enveloped HAVs acquire an envelope made from the host cell membrane, with no transmembrane viral proteins [25,26]. Usually, during HAV infection, virions are detected in feces. However, viral particles in EVs can circulate in the blood and will not be attacked by neutralizing antibodies. In addition, packaging of viral particles inside EVs increases the likelihood of infecting cells other than hepatocytes, in particular the cells of spleen or lymph nodes, which filter blood and lymph.
3.5. Coronaviruses
Viruses of the family Coronaviridae are enveloped, 80–120 nm in size, with a viral genome consisting of large single-stranded positive RNA [157]. Some coronaviruses are known to be etiological agents of dangerous pandemic respiratory illnesses such as SARS [158], MERS [159], and COVID-19 [160]. MERS-CoV infection is associated with interaction between the viral S-protein and cell dipeptidyl peptidase-4 (DPP4) [161,162]. Other cellular cofactors for viral infection are tetraspanins. In particular, CD9 tetraspanin contributes to a more effective infectious process and leads to more severe infection in mice [163]. CD9 is a major protein of exosomes. Therefore, it can be assumed that exosome biogenesis pathways are involved in MERS-CoV infection.
Other CoV proteins are also involved in membrane modifications and can be found in EVs from infected cells. The S2-glycoprotein subunit possesses various membranotropic segments that induce membrane perturbation and could allow membrane negative curvature [164]. In addition, it was reported that M and E glycoproteins of CoV can mediate the formation and release of 100 nm “vesicles”, which are morphologically very similar to viral particles. For other viruses, the production of such vesicles greatly assists infection spread, masking real virions from immune response [165].
The SARS-CoV and SARS-CoV2 viruses use angiotensin-converting enzyme 2 (ACE2) as a receptor [166,167]. ACE2 is abundant on the surfaces of lung alveolar type II (AT2) cells, and it is also expressed in the heart, blood vessels, liver, digestive organs, and kidneys [168]. Exosomes are able to carry ACE2 [169] and deliver it to uninfected recipient cells, presumably making them susceptible to the infection. Thus, exosomes can contribute to coronavirus pathogenesis. However, clarification of exosomes’ roles in coronavirus infections requires further research.
Recent, electron microscopy-based research has demonstrated that virions of SARS-CoV and SARS-CoV2 are associated with double membrane-enveloped vesicles in infected cells [170,171]. This may be indicative of the exploitation of cellular systems for EV biogenesis by coronaviruses for their spread and evasion of the immune system, like that seen in a number of other viruses.
4. EVs in Antiviral Defense
Extracellular vesicles not only participate in viral pathogenesis, but also can help the host’s organism to resist the infection. There is a lot of evidence that secretion of exosomes and microvesicles activates antiviral immune responses due to various molecular signals contained in the vesicles [172,173,174]. Exosomal proteins are one of these signals. Epithelial cells infected with cytomegalovirus are known to secrete EVs containing viral antigens. These antigens are delivered by EVs to antigen-presenting cells, with subsequent immune response activation [175]. Similarly, with influenza A virus, exosomes transfer hemagglutinin epitopes in complex with MHCII molecules, increasing the efficiency of antigenic determinant presentation to immune cells [176]. Macrophages infected with influenza virus secrete exosomes containing a wide range of specific proteins: proteins binding fatty acids, proteins of copper metabolism, cytokines (IFN-α1, IFN-α14, IFN-β, IL-6, IL-18, TNF), and proteins involved in cellular autophagy [177]. Additionally, EVs derived from virus-infected dendrite cells are implicated in the stimulation of proliferation and differentiation of CD8+ T cells into effector cytotoxic T lymphocytes (CTLs). This occurs in induction of immune responses to tumors and in protecting neighbor cells from tumor growth [178].
RNAs contained in exosomes during viral infection can also be a signal for immune activation. The exosomes from infected cells include both viral mRNAs and microRNAs. Upon transfer to other cells, viral RNA is recognized by pattern recognition receptors that signal the organism about viral attack and trigger innate immunity reactions. Viral RNAs have been found within exosomes derived from HCV, HIV, EBV, and cytomegalovirus (CMV)-infected cells. For example, exosomes from HCV-infected cells transfer viral RNA to plasmacytoid dendritic cells, stimulating the production of IFN-α [179]. EVs secreted from HIV-infected cells carry viral RNAs, which stimulate Toll like receptor-8 (TLR8) signaling to promote TNFα release, contributing to chronic immune activation [180]. Another well-known example is EBV that exploits different types of RNAs engaged in vesicles to block the antiviral response. Viral proteins encoded with mRNAs gene transcripts that play key roles in the latent infectious phase were found in the exosomes from EBV-infected cells [138]. Several small non-coding RNAs (EBER1 and EBER2) have been found in EVs secreted from EBV-infected cells, they are suspected of supporting the survival of infected cells by avoiding cell apoptosis [139]. In addition miRNAs (miR-BART15-3p) presented in exosomal cargo from EBV-infected cells were shown to induces apoptosis in immune cells [134]. In addition, profiling of EBV-induced lymphoblastoid cell lines revealed that long non-coding RNAs H19 and H19 antisense exist in secreted exosomes, and they are suggested to play a role in the regulation of the microenvironment of cells [181]. In the case of cytomegalovirus (CMV)-infected cells, the herpes simplex virus 1 (HSV-1) secreted exosomes containing viral mRNAs and microRNAs, whose major function may be in silencing viral genes in latently infected neurons. Moreover, this pull of RNAs is presented in exosomes rather than in virions. It has been suggested that HSV-1 controls the spread of infection between cells using exosomal RNAs for curtailing the infection under certain conditions [182].
HIV-infected macrophages secrete extracellular vesicles containing viral miRNA, which stimulates TLR8 and NF-κB signaling pathways when passed to other macrophages. As a result, the production of proinflammatory cytokines (for example, TNF-α) is enhanced, and the immune response against HIV is activated [180]. Cells infected with HSV-1 secrete exosomes containing the viral miRNAs miR-H28 and miR-H29. Inhibition of viral replication and spread has been observed after exosomal transfer of these microRNAs to neighboring uninfected cells [183].
Numerous microRNAs of cellular origin are also contained in exosomes and transferred by them from infected to intact cells, triggering an antiviral response. For example, exosomes derived from the bronchoalveolar lavage of patients infected with the influenza virus accumulate miR-483-3p, mir-374-5p, and miR-446i-5p. These microRNAs promote the expression of IFN-β, proinflammatory cytokines, and interferon-stimulated genes, including IL-6, TNF-α, C-C motif chemokine ligand 2 (CCL2), and SP100 [184]. Another example is the high resistance to viral infection in trophoblast cells, which protects the fetus from infection. These cells secrete exosomes containing a specific set of cellular microRNAs (chromosome 19 miRNA cluster), and transfer of these microRNAs to recipient cells makes them less sensitive to a wide range of viruses [185].
Besides various molecular signals, exosomes from infected cells also contain effector molecules, directly participating in the antiviral host response. It is known that CD4+ T-cells secrete specific extracellular vesicles containing CD4 (CD4 is an HIV receptor). These EVs are able to bind HIV virions, ultimately decreasing the infection. HIV counteracts this defense mechanism by loading its Nef protein into the EVs secreted by infected cells. Nef, being delivered to T cells, prevents the incorporation of CD4 into EVs and suppresses the antiviral response described above [186].
Exosomes isolated from HIV-infected cells were also shown to carry cytidine deaminase, which inhibits viral replication in cells [187]. Cytidine deaminase APOBEC3G is incorporated into virions and causes deamination of cytidine in the synthesized single-stranded DNA during reverse transcription of viral RNA [188]. Structural remodeling of the synthesized DNA further leads to G-A point mutations. However, the APOBEC3G effect can be blocked by the viral protein Vif, which binds APOBEC3G and promotes its ubiquitinylation, with subsequent degradation in proteosomes [189].
Exosomes derived from respiratory tract epithelial cells during influenza A infection carry mucins (MUC1, MUC4, MUC16) [190], which are bound to sialic acids and are capable of virus neutralization. With Dengue virus, infected cells secrete exosomes with the effector antiviral protein IFITM3. IFITM3-containing exosomes are transferred to uninfected cells and equip them before they face the viral invasion [191].
One of the promising approaches to COVID-19 therapy is the use of multipotent mesenchymal stromal cells (MMSCs), which can reduce the cytokine storm in patients with severe disease [192]. After systemic injection, some of the MMSCs accumulate in the lungs, helping to restore their functionality. MMSCs are resistant to viral infection because they actively express IFN-stimulated genes (ISGs) and do not carry ACE2 (SARS-CoV-2 receptor) on their surfaces [193]. MMSCs affect innate and adaptive immune cells (including T-cells, B-cells, dendritic cells, macrophages, and natural killer cells) by secreting a wide pool of mediators of various types [194,195]. In addition, MMSCs secrete extracellular vesicles containing these mediators [196]. This gave rise to the idea of therapeutic usage of MMSC-derived EVs, instead of MMSCs themselves [197,198,199]. This is much safer and more practical, compared to MMSC-cell therapy; development of inhalational preparations is possible in this case.
A widespread complication of SARS-CoV-2 is myocardial injury, including ischemia and heart attack [200,201]. Treatment with MMSC exosomes has been experimentally proven to reduce infarction damage, to reduce oxidative stress, and to increase levels of NADH and ATP. Apparently, this is due to activation of the PI3K/Akt signaling pathway responsible for cell viability after myocardial injury [202,203]. Exosomes carrying the S-protein were also investigated as a candidate SARS-CoV vaccine in vivo. After injection, a significant titer of neutralizing antibodies against the S-protein in blood was detected in mice [204].
Thus, detailed studies of EV biology are necessary for understanding the mechanisms of exploitation of cellular systems of biogenesis and secretion of EVs by viruses and the deregulation of cellular processes under their influence. Multiple similarities between EVs and viruses in their organization, assembly, and molecular composition suggest approaches for development of new generation vaccines and biomimetic designs for targeted drug delivery [205]. In particular, COVID-19 treatment with MMSC-derived exosomes is a promising new approach. It requires, however, much more research to standardize isolation conditions, composition, storage, and dose selection depending on illness severity.
5. Conclusions
Analysis of the roles of extracellular vesicles during viral infection is extremely important for understanding the mechanisms of pathogenesis and possible ways to resist viral infection. Viruses can use the EV biogenesis system to activate their own secretion or to package viral material into vesicles. EV-mediated transfer of viral proteins and nucleic acids (including whole genomes and completely assembled viral particles) enhances viral spread and provides viruses with evasion of the immune response [205]. In addition, extracellular vesicles are capable of delivering surface receptors to uninfected cells for subsequent binding with viral particles, which leads to an increasing number of cells sensitive to viral infection. On the other hand, transfer and presentation of individual viral components to immune cells by EVs can trigger antiviral reactions. Exosomes could be good therapeutic agents, because they are non-immunogenic and can pass the cellular barriers, and the contents can be manipulated. EVs are capable of transporting both hydrophobic (inside) and hydrophilic (in the compound of the lipid membrane) molecules [206]. However, EVs are not optimized for the encapsulation of hydrophilic molecules. The loading capacity of the exosomes is rather low due to the presence of proteins and nucleic acids within. To improve cell targeting, exosomes should be generated by cells expressing ligands with a high binding affinity to the target cells [207,208]. However, there are some reported positive results in implementing exosomal delivery for therapeutics against neurotropic viruses [209]. On that basis, EVs are considered a promising component for use in new generation vaccine designs [210].
Author Contributions
Conceptualization, L.P., A.G. and A.V.; writing—original draft preparation, L.P. and A.G.; writing—review and editing, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 20-15-00228.
Acknowledgments
We are grateful to Edward S. Ramsay for the professional editing of the text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zullo, J.; Matsumoto, K.; Xavier, S.; Ratliff, B.; Goligorsky, M.S. The cell secretome, mediator of cell-to-cell communication. Prostaglandins Other Lipid Mediat. 2016, 120, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busatto, S.; Zendrini, A.; Radeghieri, A.; Paolini, L.; Romano, M.; Presta, M.; Bergese, P. The nanostructured secretome. Biomater. Sci. 2019, 8, 39–63. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocytematuration. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Conde-Vancells, J.; Rodriguez-Suarez, E.; Gonzalez, E.; Berisa, A.; Embade, N.; Gil, D.; Matthiesen, R.; Valle, M.; Elortza, F.; Wagner, C.; et al. Candidate biomarkers in exosome-like vesicles purified from rat and mouse urine samples. Proteom. Clin. 2010, 4, 416–425. [Google Scholar] [CrossRef] [Green Version]
- Caby, M.P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like vesicles are present in human blood plasma. Int. Immunol. 2005, 17, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Michael, A.; Bajracharya, S.D.; Yuen, P.S.T.; Zhou, H.; Star, R.A.; Illei, G.G.; Alevizos, I. Exosomes from human saliva as a source of microRNA biomarkers. Oral Dis. 2010, 16, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Admyre, C.; Johansson, S.M.; Rahman Qazi, K.; Filén, J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef]
- Prado, N.; Marazuela, E.G.; Segura, E.; Fernández-García, H.; Villalba, M.; Théry, C.; Rodríguez, R.; Batanero, E. Exosomes from bronchoalveolar fluid of tolerized mice prevent allergic reaction. J. Immunol. 2008, 181, 1519–1525. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Rupp, C.; Stoeck, A.; Runz, S.; Fogel, M.; Lugert, S.; Hager, H.D.; Abdel-Bakky, M.S.; Gutwein, P.; Altevogt, P. CD24 is a marker of exosomes secreted into urine and amniotic fluid. Kidney Int. 2007, 72, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, 968–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-K.; Lee, J.; Kim, S.R.; Choi, D.-S.; Yoon, Y.J.; Kim, J.H.; Jang, S.C. EVpedia: A community web portal for extracellular vesicles research. Bioinformatics 2014, 31, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucl. Acids Res. 2018, 47, 516–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Mathivanan, S. ExoCarta: A web-based compendium of exosomal cargo. J. Molec. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frängsmyr, L.; Baranov, V.; Nagaeva, O.; Stendahl, U.; Kjellberg, L.; Mincheva- Nilsson, L. Cytoplasmic microvesicular form of Fas ligand in human early placenta: Switching the tissue immune privilege hypothesis from cellular to vesicular level. Mol. Hum. Reprod. 2005, 11, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Niel, G.; Mallegol, J.; Bevilacqua, C.; Candalh, C.; Brugie’re, S.; Tomaskovic-Crook, E.; Heath, J.K.; Cerf-Bensussan, N.; Heyman, M. Intestinal epithelial exosomes carry MHC class II/peptides able to inform the immune system in mice. Gut 2003, 52, 1690–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKenzie, A.; Wilson, H.L.; Kiss-Toth, E.; Dower, S.K.; North, R.A.; Surprenant, A. Rapid secretion of interleukin-1beta by microvesicle shedding. Immunity 2001, 15, 825–835. [Google Scholar] [CrossRef]
- Barros, F.M.; Carneiro, F.; Machado, J.C.; Melo, S.A. Exosomes and immune response in cancer: Friends or foes? Front. Immunol. 2018, 9, 730. [Google Scholar] [CrossRef]
- Xu, R.; Rai, A.; Chen, M.; Suwakulsiri, W.; Greening, D.W.; Simpson, R.J. Extracellular vesicles in cancer—Implications for future improvements in cancer care. Nat. Rev. Clin. Oncol. 2018, 15, 617–638. [Google Scholar] [CrossRef]
- Huber, V.; Fais, S.; Iero, M.; Lugini, L.; Canese, P.; Squarcina, P. Human colorectal cancer cells induce T-cell death through release of proapoptotic microvesicles: Role in immune escape. Gastroenterology 2005, 128, 1796–1804. [Google Scholar] [CrossRef]
- Rajendran, L.; Honsho, M.; Zahn, T.R.; Keller, P.; Geiger, K.D.; Verkade, P.; Simons, K. Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc. Natl. Acad. Sci. USA 2006, 103, 11172–11177. [Google Scholar] [CrossRef] [Green Version]
- Vella, L.J.; Sharples, R.A.; Lawson, V.A.; Masters, C.L.; Cappai, R.; Hill, A.F. Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J. Pathol. 2007, 211, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.W.; Lichty, B.D.; Bowdish, D.M. Microvesicles: Ubiquitous contributors to infection and immunity. J. Leukoc. Biol. 2015, 97, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.R.; Kashanchi, F.; Jacobson, S. Exosomes in Viral Disease. Neurotherapeutics 2016, 13, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Meckes, D.G. Exosomal communication goes viral. J. Virol. 2015, 89, 5200–5203. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.-H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Hirai-Yuki, A.; Hensley, L.; Whitmire, J.K.; Lemon, S.M. Biliary secretion of quasi-enveloped human hepatitis A virus. MBio 2016, 7, e01998-16. [Google Scholar] [CrossRef] [Green Version]
- Silverman, J.M.; Reiner, N.E. Exosomes and other microvesicles in infection biology: Organelles with unanticipated phenotypes. Cell. Microbiol. 2010, 13, 1–9. [Google Scholar] [CrossRef]
- Meckes, D.G.; Raab-Traub, N. Microvesicles and viral infection. J. Virol. 2011, 85, 12844–12854. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.M.; Mathew, A.; Mason, A.B.; Teng, K. Exosome formation during maturation of mammalian and avian reticulocytes: Evidence that exosome release is a major route for externalization of obsolete membrane proteins. J. Cell Physiol. 1991, 147, 27–36. [Google Scholar] [CrossRef]
- Stoorvogel, W.; Kleijmeer, M.J.; Geuze, H.J.; Raposo, G. The biogenesis and functions of exosomes. Traffic 2002, 3, 321–330. [Google Scholar] [CrossRef]
- Van Meel, E.; Klumperman, J. Imaging and imagination: Understanding the endo-lysosomal system. Histochem. Cell Biol. 2008, 3, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.L.; Urbé, S. The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell Biol. 2007, 8, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Odorizzi, G.; Emr, S.D. Receptor downregulation and multivesicular-body sorting. Nat. Rev. Mol. Cell Biol. 2002, 12, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Babst, M.; Emr, S.D. Ubiquitin-dependent sorting into the multivesicular body pathway requires the function of a conserved endosomal protein sorting complex, ESCRT-I. Cell 2001, 106, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Février, B.; Raposo, G. Exosomes: Endosomal-derived vesicles shipping extracellular messages. Curr. Opin. Cell Biol. 2004, 16, 415–421. [Google Scholar] [CrossRef]
- Raiborg, C.; Rusten, T.E.; Stenmark, H. Protein sorting into multivesicular endosomes. Curr. Opin. Cell Biol. 2003, 15, 446–455. [Google Scholar] [CrossRef]
- Wollert, T.; Yang, D.; Ren, X.; Lee, H.H.; Im, Y.J.; Hurley, J.H. The ESCRT machinery at a glance. J Cell Sci. 2009, 122, 2163–2166. [Google Scholar] [CrossRef] [Green Version]
- Stuffers, S.; Sem Wegner, C.; Stenmark, H.; Brech, A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009, 10, 925–937. [Google Scholar] [CrossRef]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brügger, B.; Simons, M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.S.; Rubinstein, E.; Raposo, G. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev. Cell 2011, 21, 708–721. [Google Scholar]
- Edgar, J.R.; Eden, E.R.; Futter, C.E. Hrs- and CD63-dependent competing mechanisms make different sized endosomal intraluminal vesicles. Traffic 2014, 15, 197–211. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small GTPases 2017, 8, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, D.; Veitch, S.; Fish, J.E. Extracellular vesicles as protagonists of diabetic cardiovascular pathology. Front. Cardiovasc. Med. 2017, 4, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, L.S.; Faria, L.P.; Rody, W.J., Jr. Actin and actin-associated proteins in extracellular vesicles shed by osteoclasts. Int. J. Mol. Sci. 2019, 21, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Lu, Q. Plasma membrane-derived extracellular microvesicles mediate non-canonical intercellular NOTCH signaling. Nat. Commun. 2017, 8, 709. [Google Scholar] [CrossRef]
- Atkin-Smith, G.K.; Tixeira, R.; Paone, S.; Mathivanan, S.; Collins, C.; Liem, M.; Goodall, K.J.; Ravichandran, K.S.; Hulett, M.D.; Poon, I. A novel mechanism of generating extracellular vesicles during apoptosis via a beads-on-a-string membrane structure. Nat. Commun. 2015, 6, 7439. [Google Scholar] [CrossRef]
- Caruso, S.; Poon, I.K.H. Apoptotic cell-derived extracellular vesicles: More than just debris. Front. Immunol. 2018, 9, 1486. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Erwig, L.P.; Henson, P.M. Clearance of apoptotic cells by phagocytes. Cell Death Differ. 2008, 15, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Odorizzi, G. Get on the exosome bus with alix. Nat. Cell Biol. 2012, 14, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Savina, A.; Fader, C.M.; Damiani, M.T.; Colombo, M.I. Rab11 promotes docking and fusion of multivesicular bodies in a calcium-dependent manner. Traffic 2005, 6, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Maji, S.; Chaudhary, P.; Akopova, I.; Nguyen, P.M.; Hare, R.J.; Gryczynski, I.; Vishwanatha, J.K. Exosomal Annexin II Promotes Angiogenesis and Breast Cancer Metastasis. Mol. Cancer Res. 2017, 15, 93–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldolesi, J. Exosomes and ectosomes in intercellular communication. Curr. Biol. 2018, 28, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Shimaoka, M.; Kawamoto, E.; Gaowa, A.; Okamoto, T.; Jeong Park, E. Connexins and Integrins in Exosomes. Cancers 2019, 11, 106. [Google Scholar] [CrossRef] [Green Version]
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of Extracellular HSPs as Biomarkers in Immune Surveillance and Immune Evasion. Int. J. Mol. Sci. 2019, 20, 4588. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of exosome composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef] [Green Version]
- Llorente, A.; Skotland, T.; Sylvänne, T.; Kauhanen, D.; Róg, T.; Orłowski, A.; Vattulainen, I.; Ekroos, K.; Sandvig, K. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim. Biophys. Acta 2013, 1831, 1302–1309. [Google Scholar] [CrossRef]
- Sagini, K.; Costanzi, E.; Emiliani, C.; Buratta, S.; Urbanelli, L. Extracellular vesicles as conveyors of membrane-derived bioactive lipids in immune system. Int. J. Mol. Sci. 2018, 19, 1227. [Google Scholar] [CrossRef] [Green Version]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagini, K.; Urbanelli, L.; Costanzi, E.; Mitro, N.; Caruso, D.; Emiliani, C.; Buratta, S. Oncogenic H-Ras expression induces fatty acid profile changes in human fibroblasts and extracellular vesicles. Int. J. Mol. Sci. 2018, 19, 3515. [Google Scholar] [CrossRef] [Green Version]
- Bellingham, S.A.; Coleman, B.M.; Hill, A.F. Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucl. Acids Res. 2012, 21, 10937–10949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.M.; Abdelmohsen, K.; Mustapic, M.; Kapogiannis, D.; Gorospe, M. RNA in extracellular vesicles. Wiley Interdiscip. Rev. RNA 2017, 8, e1413. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Odenthal, M.; Fries, J.W. Exosomes as miRNA carriers: Formation-function-future. Int. J. Mol. Sci. 2016, 17, 2028. [Google Scholar] [CrossRef] [PubMed]
- Boorn, J.G.; Daßler, J.; Coch, C.; Schlee, M.; Hartmann, G. Exosomes as nucleic acid nanocarriers. Adv. Drug Deliv. Rev. 2013, 65, 331–333. [Google Scholar] [CrossRef]
- Guescini, M.; Genedani, S.; Stocchi, V.; Agnati, L.F. Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J. Neural. Transm. 2010, 117, 1–4. [Google Scholar] [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes end exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Muralidharan-Chari, V.; Clancy, J.; Plou, C.; Romao, M.; Chavrier, P.; Raposo, G.; D’Souza-Schorey, C. ARF6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr. Biol. 2009, 19, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Haraszti, R.A.; Didiot, M.C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J. Extracell. Vesicl. 2016, 5, 32570. [Google Scholar] [CrossRef] [PubMed]
- Crescitelli, R.; Lasser, C.; Szabo, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzas, E.I.; Lotvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicl. 2013, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Waldenström, A.; Gennebäck, N.; Hellman, U.; Ronquist, G. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS ONE 2012, 7, e34653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Hauser, P.; Wang, S.; Didenko, V.V. Apoptotic bodies: Selective detection in extracellular vesicles. Methods Mol Biol. 2017, 1554, 193–200. [Google Scholar] [CrossRef]
- Choi, D.S.; Kim, D.K.; Kim, Y.K.; Gho, Y.S. Proteomics of extracellular vesicles: Exosomes and ectosomes. Mass. Spectrom. Rev. 2015, 34, 474–490. [Google Scholar] [CrossRef]
- Cantin, R.; Diou, J.; Bélanger, D.; Tremblay, A.M.; Gilbert, C. Discrimination between exosomes and HIV-1: Purification of both vesicles from cell-free supernatants. J. Immunol. Methods 2008, 338, 21–30. [Google Scholar] [CrossRef]
- Chugh, P.E.; Sin, S.H.; Ozgur, S.; Henry, D.H.; Menezes, P.; Griffith, J.; Eron, J.J.; Damania, B.; Dittmer, D.P. Systemically circulating viral and tumor-derived microRNAs in KSHV-associated malignancies. PLoS Pathog. 2013, 9, e1003484. [Google Scholar] [CrossRef] [Green Version]
- Amen, M.A.; Griffiths, A. Packaging of non-coding RNAs into herpesvirus virions: Comparisons to coding RNAs. Front. Genet. 2011, 2, 81. [Google Scholar] [CrossRef] [Green Version]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2004, 24, 766–769. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, A.; Villar-Prados, A.; Oliphint, P.A.; Zhang, J.; Song, X.; De Hoff, P.; Morey, R.; Liu, J.; Roszik, J.; Clise-Dwyer, K.; et al. Mechanisms of nuclear content loading to exosomes. Sci. Adv. 2019, 5, eaax8849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torralba, D.; Baixauli, F.; Villarroya-Beltri, C.; Fernández-Delgado, I.; Latorre-Pellicer, A.; Acín-Pérez, R.; Martín-Cófreces, N.B.; Jaso-Tamame, A.L.; Iborra, S.; Jorge, I.; et al. Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat. Commun. 2018, 9, 1–17. [Google Scholar]
- Li, Y.; Guo, X.; Guo, S.; Wang, Y.; Chen, L.; Liu, Y.; Jia, M.; An, J.; Tao, K.; Xing, J. Next generation sequencing-based analysis of mitochondrial DNA characteristics in plasma extracellular vesicles of patients with hepatocellular carcinoma. Oncol. Lett. 2020, 20, 2820–2828. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host–pathogen interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.J.; Lamb, R.A. Mechanisms for enveloped virus budding: Can some viruses do without an ESCRT? Virology 2008, 372, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Harrison, M.S.; Sakaguchi, T.; Schmitt, A.P. Paramyxovirus assembly and budding: Building particles that transmit infections. Int. J. Biochem. Cell Biol. 2010, 42, 1416–1429. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Koike, M.; Moriishi, E.; Kawabata, A.; Tang, H.; Oyaizu, H.; Uchiyama, Y.; Yamanishi, K. Human herpesvirus-6 induces MVB formation, and virus egress occurs by an exosomal release pathway. Traffic 2008, 9, 1728–1742. [Google Scholar] [CrossRef] [Green Version]
- Kharkwal, H.; Smith, C.G.; Wilson, D.W. Herpes simplex virus capsid localization to ESCRT-VPS4 complexes in the presence and absence of the large tegument protein UL36p. J. Virol. 2016, 90, 7257–7267. [Google Scholar] [CrossRef] [Green Version]
- Votteler, J.; Sundquist, W.I. Virus budding and the ESCRT pathway. Cell Host Microbe 2013, 14, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Dongen, H.M.; Masoumi, N.; Witwer, K.W.; Pegtel, D.M. Extracellular vesicles exploit viral entry routes for cargo delivery. Microbiol. Mol. Biol. Rev. 2016, 80, 369–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, P.C.; Han, J.; Chadha, P.; Meckes, D.G.; Ward, M.D., Jr.; Semmes, O.J.; Wills, J.W. Direct and specific binding of the UL16 tegument protein of herpes simplex virus to the cytoplasmic tail of glycoprotein. J. Virol. 2011, 85, 9425–9436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meckes, D.G., Jr.; Gunawardena, H.P.; Dekroon, R.M.; Heaton, P.R.; Edwards, R.H.; Ozgur, S.; Griffith, J.D.; Damania, B.; Raab-Traub, N. Modulation of B-cell exosome proteins by gamma herpesvirus infection. Proc. Natl. Acad. Sci. USA 2013, 110, 2925–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallhov, H.; Gutzeit, C.; Johansson, S.M.; Nagy, N.; Paul, M.; Li, Q.; Friend, S.; George, T.C.; Klein, E.; Scheynius, A.; et al. Exosomes containing glycoprotein 350 released by EBV-transformed B cells selectively target B cells through CD21 and block EBV infection in vitro. J. Immunol. 2011, 186, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, J.; Middeldorp, J.; Sculley, T. Localization of the Epstein-Barr virus protein LMP 1 to exosomes. J. Gen. Virol. 2003, 84, 1871–1879. [Google Scholar] [CrossRef]
- Verweij, F.J.; van Eijndhoven, M.A.J.; Hopmans, E.S.; Vendrig, T.; Wurdinger, T.; Cahir-McFarland, E.; Kieff, E.; Geerts, D.; van der Kant, R.; Neefjes, J.; et al. LMP1 association with CD63 in endosomes and secretion via exosomes limits constitutive NF-kappaB activation. EMBO J. 2011, 30, 2115–2129. [Google Scholar] [CrossRef]
- Keryer-Bibens, C.; Pioche-Durieu, C.; Villemant, C.; Souquère, S.; Nishi, N.; Hirashima, M.; Middeldorp, J.; Busson, P. Exosomes released by EBV-infected nasopharyngeal carcinoma cells convey the viral latent membrane protein 1 and the immunomodulatory protein galectin 9. BMC Cancer 2006, 6, 283. [Google Scholar] [CrossRef] [Green Version]
- Klibi, J.; Niki, T.; Riedel, A.; Pioche-Durieu, C.; Souquere, S.; Rubinstein, E.; Le Moulec, S.; Guigay, J.; Hirashima, M.; Guemira, F.; et al. Blood diffusion and Th1-suppressive effects of galectin-9-containing exosomes released by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Blood 2009, 113, 1957–1966. [Google Scholar] [CrossRef] [Green Version]
- Meckes, D.G., Jr.; Shair, K.H.Y.; Marquitz, A.R.; Kung, C.P.; Edwards, R.H.; Raab-Traub, N. Human tumor virus utilizes exosomes for intercellular communication. Proc. Natl. Acad. Sci. USA 2010, 107, 20370–20375. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, S.; Visco, V.; Raffa, S.; Wakisaka, N.; Pagano, S.; Torrisi, M.R. Epstein-Barr virus latent membrane protein 1 promotes concentration in multivesicular bodies of fibroblast growth factor 2 and its release through exosomes. Int. J. Cancer 2007, 121, 1494–1506. [Google Scholar] [CrossRef]
- Aga, M.; Bentz, G.L.; Raffa, S.; Torrisi, M.R.; Kondo, S.; Wakisaka, N.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. Exosomal HIF1alpha supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene 2014, 33, 4613–4622. [Google Scholar] [CrossRef] [Green Version]
- Skalsky, R.L.; Corcoran, D.L.; Gottwein, E.; Frank, C.L.; Kang, D.; Hafner, M.; Nusbaum, J.D.; Feederle, R.; Delecluse, H.J.; Luftig, M.A.; et al. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS Pathog. 2012, 8, e1002484. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wu, N.; Gan, X.; Yan, W.; Morrell, J.C.; Gould, S.J. Higher-order oligomerization targets plasma membrane proteins and HIV gag to exosomes. PLoS Biol. 2007, 5, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vanpouille, C.; Margolis, L. Extracellular Vesicles Carry HIV Env and Facilitate Hiv Infection of Human Lymphoid Tissue. Sci. Rep. 2017, 7, 1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenassi, M.; Cagney, G.; Liao, M.; Vaupotic, T.; Bartholomeeusen, K.; Cheng, Y.; Krogan, N.J.; Plemenitas, A.; Peterlin, B.M. HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic 2010, 11, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Mack, M.; Kleinschmidt, A.; Brühl, H.; Klier, C.; Nelson, P.J.; Cihak, J.; Plachý, J.; Stangassinger, M.; Erfle, V.; Schlöndorff, D. Transfer of the chemokine receptor CCR5 between cells by membrane-derived microparticles: A mechanism for cellular human immunodeficiency virus 1 infection. Nat. Med. 2000, 6, 769–775. [Google Scholar] [CrossRef]
- Rozmyslowicz, T.; Majka, M.; Kijowski, J.; Murphy, S.L.; Conover, D.O.; Poncz, M.; Ratajczak, J.; Gaulton, G.N.; Ratajczak, M.Z. Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV. AIDS 2003, 17, 33–42. [Google Scholar] [CrossRef]
- Narayanan, A.; Iordanskiy, S.; Das, R.; Van Duyne, R.; Santos, S.; Jaworski, E.; Guendel, I.; Sampey, G.; Dalby, E.; Iglesias-Ussel, M.; et al. Exosomes derived from HIV-1-infected cells contain trans-activation response element RNA. J. Biol. Chem. 2013, 288, 20014–20033. [Google Scholar] [CrossRef] [Green Version]
- Masciopinto, F.; Giovani, C.; Campagnoli, S.; Galli-Stampino, L.; Colombatto, P.; Brunetto, M.; Yen, T.S.B.; Houghton, M.; Pileri, P.; Abrignani, S. Association of hepatitis C virus envelope proteins with exosomes. Eur. J. Immunol. 2004, 34, 2834–2842. [Google Scholar] [CrossRef]
- Bukong, T.N.; Momen-Heravi, F.; Kodys, K.; Bala, S.; Szabo, G. Exosomes from hepatitis C infected patients transmit HCV infection and contain replication competent viral RNA in complex with Ago2-miR122-HSP90. PLoS Pathog. 2014, 10, e1004424. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Ma, P.; Deng, L.; Liu, Z.; Wang, X.; Liu, X.; Long, G. Hepatitis A virus structural protein pX interacts with ALIX and promotes the secretion of virions and foreign proteins through exosome-like vesicles. J. Extracell. Vesicl. 2020, 9, 1716513. [Google Scholar] [CrossRef] [PubMed]
- Kalamvoki, M.; Deschamps, T. Extracellular vesicles during Herpes simplex virus type 1 infection: An inquire. Virol. J. 2016, 13, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLauchlan, J.; Rixon, F.J. Characterization of enveloped tegument structures (L particles) produced by alphaherpesviruses: Integrity of the tegument does not depend on the presence of capsid or envelope. J. Gen. Virol. 1992, 73, 269–276. [Google Scholar] [CrossRef] [PubMed]
- McLauchlan, J.; Addison, C.; Craigie, M.C.; Rixon, F.J. Noninfectious L-particles supply functions which can facilitate infection by HSV-1. Virology 1992, 190, 682–688. [Google Scholar] [CrossRef]
- Dargan, D.J.; Subak-Sharpe, J.H. The effect of herpes simplex virus type 1 L-particles on virus entry, replication, and the infectivity of naked herpesvirus DNA. Virology 1997, 239, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.J.; Fraefel, C.; Cunningham, A.L.; Diefenbach, R.J. Functional roles of the tegument proteins of herpes simplex virus type 1. Virus Res. 2009, 145, 173–186. [Google Scholar] [CrossRef]
- Rixon, F.J.; Addison, C.; McLauchlan, J. Assembly of enveloped tegument structures (L particles) can occur independently of virion maturation in herpes simplex virus type 1-infected cells. J. Gen. Virol. 1992, 73, 277–284. [Google Scholar] [CrossRef]
- Heilingloh, C.S.; Kummer, M.; Mühl-Zürbes, P.; Drassner, C.; Daniel, C.; Klewer, M.; Steinkasserer, A. L-Particles transmit viral proteins from herpes simplex virus 1-infected mature dendritic cells to uninfected bystander cells, inducing CD83 down modulation. J. Virol. 2015, 89, 11046–11055. [Google Scholar] [CrossRef] [Green Version]
- Temme, S.; Eis-Hübinger, A.M.; McLellan, A.D.; Koch, N. The herpes simplex virus-1 encoded glycoprotein B diverts HLA-DR into the exosome pathway. J. Immunol. 2010, 184, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Neumann, J.; Eis-Hübinger, A.M.; Koch, N. Herpes simplex virus type 1 targets the MHC class II processing pathway for immune evasion. J. Immunol. 2003, 171, 3075–3083. [Google Scholar] [CrossRef] [Green Version]
- Bello-Morales, R.; López-Guerrero, J.A. Extracellular vesicles in herpes viral spread and immune evasion. Front. Microbiol. 2018, 9, 2572. [Google Scholar] [CrossRef] [Green Version]
- Epstein, A. Why and how Epstein-Barr virus was discovered 50 years ago. Curr. Top. Microbiol. Immunol. 2015, 390, 3–15. [Google Scholar] [PubMed]
- Wang, L.W.; Jiang, S.; Gewurz, B.E. Epstein-Barr virus LMP1-mediated oncogenicity. J. Virol. 2017, 91, e01718-16. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Liebowitz, D.; Kieff, E. An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells. Cell 1985, 43, 831–840. [Google Scholar] [CrossRef]
- Hurwitz, S.N.; Nkosi, D.; Conlon, M.M.; York, S.B.; Liu, X.; Tremblay, D.C.; Meckes, D.G., Jr. CD63 regulates Epstein-Barr virus LMP1 exosomal packaging, enhancement of vesicle production, and noncanonical NF-kappaB signaling. J. Virol. 2017, 91, e02251-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardila-Osorio, H.; Pioche-Durieu, C.; Puvion-Dutilleul, F.; Clausse, B.; Wiels, J.; Miller, W.; Raab-Traub, N.; Busson, P. TRAF interactions with raft-like buoyant complexes, better than TRAF rates of degradation, differentiate signaling by CD40 and EBV latent membrane protein 1. Int. J. Cancer 2005, 113, 267–275. [Google Scholar] [CrossRef]
- Kaye, K.M.; Izumi, K.M.; Kieff, E. Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA 1993, 90, 9150–9154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukers, D.F.; Meij, P.; Vervoort, M.B.; Vos, W.; Scheper, R.J.; Meijer, C.J.; Bloemena, E.; Middeldorp, J.M. Direct immunosuppressive effects of EBV-encoded latent membrane protein 1. J. Immunol. 2000, 165, 663–670. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Würdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [Green Version]
- Verweij, F.J.; van Eijndhoven, M.A.; Middeldorp, J.; Pegtel, D.M. Analysis of viral microRNA exchange via exosomes in vitro and in vivo. Methods Mol. Biol. 2013, 1024, 53–68. [Google Scholar]
- Grundhoff, A.; Sullivan, C.S. Virus-encoded microRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, T.; O’Hara, A.; Araujo, I.; Barreto, J.; Carvalho, E.; Sapucaia, J.B.; Ramos, J.C.; Luz, E.; Pedroso, C.; Manrique, M.; et al. EBV microRNAs in primary lymphomas and targeting of CXCL-11 by ebv-mir-BHRF1-3. Cancer Res. 2008, 68, 1436–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O’Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1β production. J. Immunol. 2012, 189, 3795–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Lee, H.; Kim, S.R.; Gho, Y.S.; Lee, S.K. Epstein-Barr virus-encoded microRNA BART15-3p promotes cell apoptosis partially by targeting BRUCE. J. Virol. 2013, 87, 8135–8144. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Lee, S.K. TAX1BP1 downregulation by EBV-miR-BART15-3p enhances chemosensitivity of gastric cancer cells to 5-FU. Arch. Virol. 2017, 162, 369–377. [Google Scholar] [CrossRef]
- Gandhi, M.K.; Moll, G.; Smith, C.; Dua, U.; Lambley, E.; Ramuz, O.; Gill, D.; Marlton, P.; Seymour, J.F.; Khanna, R. Galectin-1 mediated suppression of Epstein-Barr virus specific T-cell immunity in classic Hodgkin lymphoma. Blood 2007, 110, 1326–1329. [Google Scholar] [CrossRef]
- Li, J.; Zeng, X.H.; Mo, H.Y.; Rolén, U.; Gao, Y.F.; Zhang, X.S.; Chen, Q.Y.; Zhang, L.; Zeng, M.S.; Li, M.Z.; et al. Functional inactivation of EBV-specific T-lymphocytes in nasopharyngeal carcinoma: Implications for tumor immunotherapy. PLoS ONE 2007, 2, e1122. [Google Scholar] [CrossRef] [Green Version]
- Canitano, A.; Venturi, G.; Borghi, M.; Ammendolia, M.G.; Fais, S. Exosomes released in vitro from Epstein-Barr virus (EBV)-infected cells contain EBV-encoded latent phase mRNAs. Cancer Lett. 2013, 337, 193–199. [Google Scholar] [CrossRef]
- Ahmed, W.; Philip, P.S.; Tariq, S.; Khan, G. Epstein-Barr virus-encoded small RNAs (EBERs) are present in fractions related to exosomes released by EBV-transformed cells. PLoS ONE 2014, 9, e99163. [Google Scholar] [CrossRef]
- Gould, S.J.; Booth, A.M.; Hildreth, J.E. The Trojan exosome hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 10592–10597. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Useros, N.; Puertas, M.C.; Borràs, F.E.; Blanco, J.; Martinez-Picado, J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol. 2011, 13, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Popov, S.; Popova, E.; Inoue, M.; Göttlinger, H.G. Human immunodeficiency virus type 1 Gag engages the Bro1 domain of ALIX/AIP1 through the nucleocapsid. J. Virol. 2008, 82, 1389–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouvenet, N.; Zhadina, M.; Bieniasz, P.D.; Simon, S.M. Dynamics of ESCRT protein recruitment during retroviral assembly. Nat. Cell Biol. 2011, 13, 394–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieniasz, P.D. The cell biology of HIV-1 virion genesis. Cell Host. Microbe 2009, 5, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Caballero, D.; Hatziioannou, T.; Martin-Serrano, J.; Bieniasz, P.D. Human immunodeficiency virus type 1 matrix inhibits and confers cooperativity on gag precursor-membrane interactions. J. Virol. 2004, 78, 9560–9563. [Google Scholar] [CrossRef] [Green Version]
- Arenaccio, C.; Chiozzini, C.; Columba-Cabezas, S.; Manfredi, F.; Affabris, E.; Baur, A.; Federico, M. Exosomes from human immunodeficiency virus type 1 (HIV-1)-infected cells license quiescent CD4+ T lymphocytes to replicate HIV-1 through a Nef- and ADAM17-dependent mechanism. J. Virol. 2014, 88, 11529–11539. [Google Scholar] [CrossRef] [Green Version]
- Shelton, M.N.; Huang, M.-B.; Ali, S.A.; Powell, M.D.; Bond, V.C. Secretion modification region-derived peptide disrupts HIV-1 Nef’s interaction with mortalin and blocks virus and Nef exosome release. J. Virol. 2012, 86, 406–419. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Santini, P.A.; Sullivan, J.S.; He, B.; Shan, M.; Ball, S.C.; Dyer, W.B.; Ketas, T.J.; Chadburn, A.; Cohen-Gould, L.; et al. HIV-1 evades virus-specific IgG2 and IgA responses by targeting systemic and intestinal B cells via long-range intercellular conduits. Nat. Immunol. 2009, 10, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Madrid, R.; Janvier, K.; Hitchin, D.; Day, J.; Coleman, S.; Noviello, C.; Bouchet, J.; Benmerah, A.; Guatelli, J.; Benichou, S. Nef-induced alteration of the early/recycling endosomal compartment correlates with enhancement of HIV-1 infectivity. J. Biol. Chem. 2005, 280, 5032–5044. [Google Scholar] [CrossRef] [Green Version]
- Stumptner-Cuvelette, P.; Jouve, M.; Helft, J.; Dugast, M.; Glouzman, A.S.; Jooss, K.; Raposo, G.; Benaroch, P. Human immunodeficiency virus-1 Nef expression induces intracellular accumulation of multivesicular bodies and major histocompatibility complex class II complexes: Potential role of phosphatidylinositol 3-kinase. Mol. Biol. Cell 2003, 14, 4857–4870. [Google Scholar] [CrossRef] [Green Version]
- Muratori, C.; Cavallin, L.E.; Krätzel, K.; Tinari, A.; De Milito, A.; Fais, S.; D’Aloja, P.; Federico, M.; Vullo, V.; Fomina, A.; et al. Massive secretion by T cells is caused by HIV Nef in infected cells and by Nef transfer to bystander cells. Cell Host Microbe 2009, 6, 218–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, N.; Liu, M.; Hsu, J.; Xue, Y.; Chou, S.; Burlingame, A.; Krogan, N.J.; Alber, T.; Zhou, Q. HIV-1 Tat and host AFF4 recruit two transcription elongation factors into a bifunctional complex for coordinated activation of HIV-1 transcription. Mol. Cell. 2010, 38, 428–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, K.; Shiina, M.; Tanaka, N.; Nakano, T.; Yamamoto, A.; Kondo, Y.; Kakazu, E.; Inoue, J.; Fukushima, K.; Sano, K.; et al. Regulation of hepatitis C virus secretion by the Hrs-dependent exosomal pathway. Virology 2012, 422, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Randall, G.; Higginbottom, A.; Monk, P.; Rice, C.M.; McKeating, J.A. CD81 is required for hepatitis C virus glycoprotein-mediated viral infection. J. Virol. 2004, 78, 1448–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnaiah, V.; Thumann, C.; Fofana, I.; Habersetzer, F.; Pan, Q.; de Ruiter, P.E.; Willemsen, R.; Demmers, J.A.A.; Raj, V.S.; Jenster, G.; et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc. Natl. Acad. Sci. USA 2013, 110, 13109–13113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brimacombe, C.L.; Grove, J.; Meredith, L.W.; Hu, K.; Syder, A.J.; Flores, M.V.; Timpe, J.M.; Krieger, S.E.; Baumert, T.F.; Tellinghuisen, T.L.; et al. Neutralizing antibody-resistant hepatitis C virus cell-to-cell transmission. J. Virol. 2011, 85, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Kuiken, T.; Fouchier, R.A.; Schutten, M.; Rimmelzwaan, G.F.; Van Amerongen, G.; van Riel, D.; Laman, J.D.; de Jong, T.; van Doornum, G.; Lim, W. Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet 2003, 362, 263–270. [Google Scholar] [CrossRef] [Green Version]
- De Groot, R.J.; Baker, S.C.; Baric, R.S.; Brown, C.S.; Drosten, C.; Enjuanes, L.; Fouchier, R.A.; Galiano, M.; Gorbalenya, A.E.; Memish, Z.A. Commentary: Middle East respiratory syndrome coronavirus (MERS-CoV): Announcement of the coronavirus study group. J. Virol. 2013, 87, 7790–7792. [Google Scholar] [CrossRef] [Green Version]
- Kannan, S.; Shaik Syed Ali, P.; Sheeza, A.; Hemalatha, K. COVID-19 (Novel Coronavirus 2019)—Recent trends. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2006–2011. [Google Scholar]
- Widagdo, W.; Sooksawasdi Na Ayudhya, S.; Hundie, G.B.; Haagmans, B.L. Host determinants of MERS-CoV transmission and pathogenesis. Viruses 2019, 11, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnest, J.T.; Hantak, M.P.; Li, K.; McCray, P.B.; Perlman, S.; Gallagher, T. The tetraspanin CD9 facilitates MERS-coronavirus entry by scaffolding host cell receptors and proteases. PLoS Pathog. 2017, 13, e1006546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, L.G.M.; Vicente, E.F.; Crusca, E.; Cilli, E.M.; Costa-Filho, A.J. SARS-CoV fusion peptides induce membrane surface ordering and curvature. Sci. Rep. 2016, 6, 37131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vennema, H.; Godeke, G.J.; Rossen, J.W.; Voorhout, W.F.; Horzinek, M.C.; Opstelten, D.J.; Rottier, P.J. Nucleocapsid-independent assembly of coronavirus-like particles by co-expression of viral envelope protein genes. EMBO J. 1996, 15, 2020–2028. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Wang, J.; Chen, S.; Bihl, J. Exosome-mediated transfer of ACE2 (angiotensin-converting enzyme 2) from endothelial progenitor cells promotes survival and function of endothelial cell. Oxid. Med. Cell. Longev. 2020, 2020, 4213541. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.J. Pathogenesis of COVID-19 from a cell biology perspective. Eur. Respir. J. 2020, 55, 2000607. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.; Travanty, E.A.; Oko, L.; Edeen, K.; Berglund, A.; Wang, J.; Ito, Y.; Holmes, K.V.; Mason, R.J. Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome-coronavirus. Amer. J. Respir. Cell Mol. Biol. 2013, 48, 742–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Leijendekker, R.; Harding, C.V.; Melief, C.J.M.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Bobrie, A.; Colombo, M.; Raposo, G.; Thery, C. Exosome secretion: Molecular mechanisms and roles in immune responses. Traffic 2011, 12, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Walker, J.D.; Maier, C.L.; Pober, J.S. Cytomegalovirus-infected human endothelial cells can stimulate allogeneic CD4+ memory T cells by releasing antigenic exosomes. J. Immunol. 2009, 182, 1548–1559. [Google Scholar] [CrossRef] [Green Version]
- Testa, J.S.; Apcher, G.S.; Comber, J.D.; Eisenlohr, L.C. Exosome-driven antigen transfer for MHC class II presentation facilitated by the receptor binding activity of influenza hemagglutinin. J. Immunol. 2010, 185, 6608–6616. [Google Scholar] [CrossRef] [Green Version]
- Cypryk, W.; Lorey, M.; Puustinen, A.; Nyman, T.A.; Matikainen, S. Proteomic and bioinformatic characterization of extracellular vesicles released from human macrophages upon influenza A virus infection. J. Proteome Res. 2017, 16, 217–227. [Google Scholar] [CrossRef]
- Hao, S.; Bai, O.; Li, F.; Yuan, J.; Laferte, S.; Xiang, J. Mature dendritic cells pulsed with exosomes stimulate efficient cytotoxic T-lymphocyte responses and antitumour immunity. Immunology 2007, 120, 90–102. [Google Scholar] [CrossRef]
- Dreux, M.; Garaigorta, U.; Boyd, B.; Décembre, E.; Chung, J.; Whitten-Bauer, C.; Wieland, S.; Chisari, F.V. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012, 12, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Bernard, M.A.; Zhao, H.; Yue, S.C.; Anandaiah, A.; Koziel, H.; Tachado, S.D. Novel HIV-1 miRNAs stimulate TNFα release in human macrophages via TLR8 signaling pathway. PLoS ONE 2014, 9, e106006. [Google Scholar] [CrossRef]
- Gallo, A.; Vella, S.; Miele, M.; Timoneri, F.; Di Bella, M.; Bosi, S.; Sciveres, M.; Conaldi, P.G. Global profiling of viral and cellular non-coding RNAs in Epstein-Barr virus-induced lymphoblastoid cell lines and released exosome cargos. Cancer Lett. 2017, 388, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Kalamvoki, M.; Du, T.; Roizman, B. Cells infected with herpes simplex virus 1 export to uninfected cells exosomes containing STING, viral mRNAs, and microRNAs. Proc. Natl. Acad. Sci. USA 2014, 111, E4991–E4996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Liu, X.; Chen, X.; Zhou, X.; Du, T.; Roizman, B.; Zhou, G. miR-H28 and miR-H29 expressed late in productive infection are exported and restrict HSV-1 replication and spread in recipient cells. Proc. Natl. Acad. Sci. USA 2016, 113, E894–E901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maemura, T.; Fukuyama, S.; Sugita, Y.; Lopes, T.J.S.; Nakao, T.; Noda, T.; Kawaoka, Y. Lung-derived exosomal miR-483-3p regulates the innate immune response to influenza virus infection. J. Infect. Dis. 2018, 217, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Delorme-Axford, E.; Donker, R.B.; Mouillet, J.-F.; Chu, T.; Bayer, A.; Ouyang, Y.; Wang, T.; Stolz, D.B.; Sarkar, S.N.; Morelli, A.E.; et al. Human placental trophoblasts confer viral resistance to recipient cells. Proc. Natl. Acad. Sci. USA 2013, 110, 12048–12053. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, J.V.; de Castro, R.O.; da Silva, E.Z.M.; Silveira, P.P.; da Silva-Januário, M.E.; Arruda, E.; Jamur, M.C.; Oliver, C.; Aguiar, R.S.; da Silva, L.L.P. Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLoS ONE 2014, 9, e113691. [Google Scholar] [CrossRef] [Green Version]
- Soros, V.B.; Yonemoto, W.; Greene, W.C. Newly synthesized APOBEC3G is incorporated into HIV virions, inhibited by HIV RNA, and subsequently activated by RNase, H. PLoS Pathog. 2007, 3, e15. [Google Scholar] [CrossRef] [Green Version]
- Khatua, A.K.; Taylor, H.E.; Hildreth, J.E.; Popik, W. Exosomes packaging APOBEC3G confer human immunodeficiency virus resistance to recipient cells. J. Virol. 2009, 83, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Donahue, J.P.; Vetter, M.L.; Mukhtar, N.A.; D’Aquila, R.T. The HIV-1 Vif PPLP motif is necessary for human APOBEC3G binding and degradation. Virology 2008, 377, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Kesimer, M.; Scull, M.; Brighton, B.; DeMaria, G.; Burns, K.; O’Neal, W.; Pickles, R.J.; Sheehan, J.K. Characterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: A possible role in innate defense. FASEB J. 2009, 23, 1858–1868. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; He, Z.; Yuan, J.; Wen, W.; Huang, X.; Hu, Y.; Lin, C.; Pan, J.; Li, R.; Deng, H.; et al. IFITM3-containing exosome as a novel mediator for anti-viral response in dengue virus infection. Cell Microbiol. 2015, 17, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Atluri, S.; Manchikanti, L.; Hirsch, J.A. Expanded umbilical cord mesenchymal stem cells (UC-MMSCs) as a therapeutic strategy in managing critically Ill COVID-19 patients: The case for compassionate use. Pain Phys. 2020, 23, 71–83. [Google Scholar]
- Wu, X.; Dao Thi, V.L.; Huang, Y.; Billerbeck, E.; Saha, D.; Hoffmann, H.H.; Wang, Y.; Silva, L.A.V.; Sarbanes, S.; Sun, T.; et al. Intrinsic immunity shapes viral resistance of stem cells. Cell 2018, 172, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Meisel, R.; Zibert, A.; Laryea, M.; Göbel, U.; Däubener, W.; Dilloo, D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [Green Version]
- Mijiritsky, E.; Gardin, C.; Ferroni, L.; Lacza, Z.; Zavan, B. Albumin-impregnated bone granules modulate the interactions between mesenchymal stem cells and monocytes under in vitro inflammatory conditions. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 110, 110678. [Google Scholar] [CrossRef]
- Ranganath, S.H.; Levy, O.; Inamdar, M.S.; Karp, J.M. Harnessing the mesenchymal stem cell secretome for the treatment of cardiovascular disease. Cell Stem Cell. 2012, 10, 244–258. [Google Scholar] [CrossRef] [Green Version]
- Phelps, J.; Sanati-Nezhad, A.; Ungrin, M.; Duncan, N.A.; Sen, A. Bioprocessing of mesenchymal stem cells and their derivatives: Toward cell-free therapeutics. Stem Cells Int. 2018, 2018, 9415367. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Zhang, Y.; Lan, B. MiRNA-sequence indicates that mesenchymal stem cells and exosomes have similar mechanism to enhance cardiac repair. BioMed Res. Int. 2017, 9. [Google Scholar] [CrossRef]
- Bari, E.; Perteghella, S.; Catenacci, L.; Sorlini, M.; Croce, S.; Mantelli, M.; Avanzini, M.A.; Sorrenti, M.; Torre, M.L. Freeze-dried and GMP-compliant pharmaceuticals containing exosomes for acellular mesenchymal stromal cell immunomodulant therapy. Nanomedicine 2019, 14, 753–765. [Google Scholar] [CrossRef]
- Akhmerov, A.; Marban, E. COVID-19 and the heart. Circ. Res. 2020, 126, 1443–1455. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Arslan, F.; Lai, R.C.; Smeets, M.B.; Akeroyd, L.; Choo, A.; Aguor, E.N.; Timmers, L.; van Rijen, H.V.; Doevendans, P.A.; Pasterkamp, G.; et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013, 10, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MMSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuate, S.; Cinatl, J.; Doerr, H.W.; Uberla, K. Exosomal vaccines containing the S protein of the SARS coronavirus induce high levels of neutralizing antibodies. Virology 2007, 362, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolte, E.; Cremer, T.; Gallo, R.C.; Margolis, L.B. Extracellular vesicles and viruses: Are they close relatives? Proc. Natl. Acad. Sci. USA 2016, 113, 9155–9161. [Google Scholar] [CrossRef] [Green Version]
- Agrahari, V.; Agrahari, V.; Mitra, A.K. Nanocarrier fabrication and macromolecule drug delivery: Challenges and opportunities. Ther. Deliv. 2016, 7, 257–278. [Google Scholar] [CrossRef] [Green Version]
- McMillan, J.; Batrakova, E.; Gendelman, H.E. Cell delivery of therapeutic nanoparticles. Prog. Mol. Biol. Transl. Sci. 2011, 104, 563–601. [Google Scholar]
- Wahid, B.; Ali, A.; Rafique, S.; Idrees, M. Current status of therapeutic and vaccine approaches against Zika virus. Eur. J. Intern. Med. 2017, 44, 12–18. [Google Scholar] [CrossRef]
- Ha, D.; Yang, N.N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome theranostics: Biology and translational medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef]
Figure 1.
Cellular routes of extracellular vesicle (EV) biogenesis and main molecular components of the exosome.
Figure 1.
Cellular routes of extracellular vesicle (EV) biogenesis and main molecular components of the exosome.


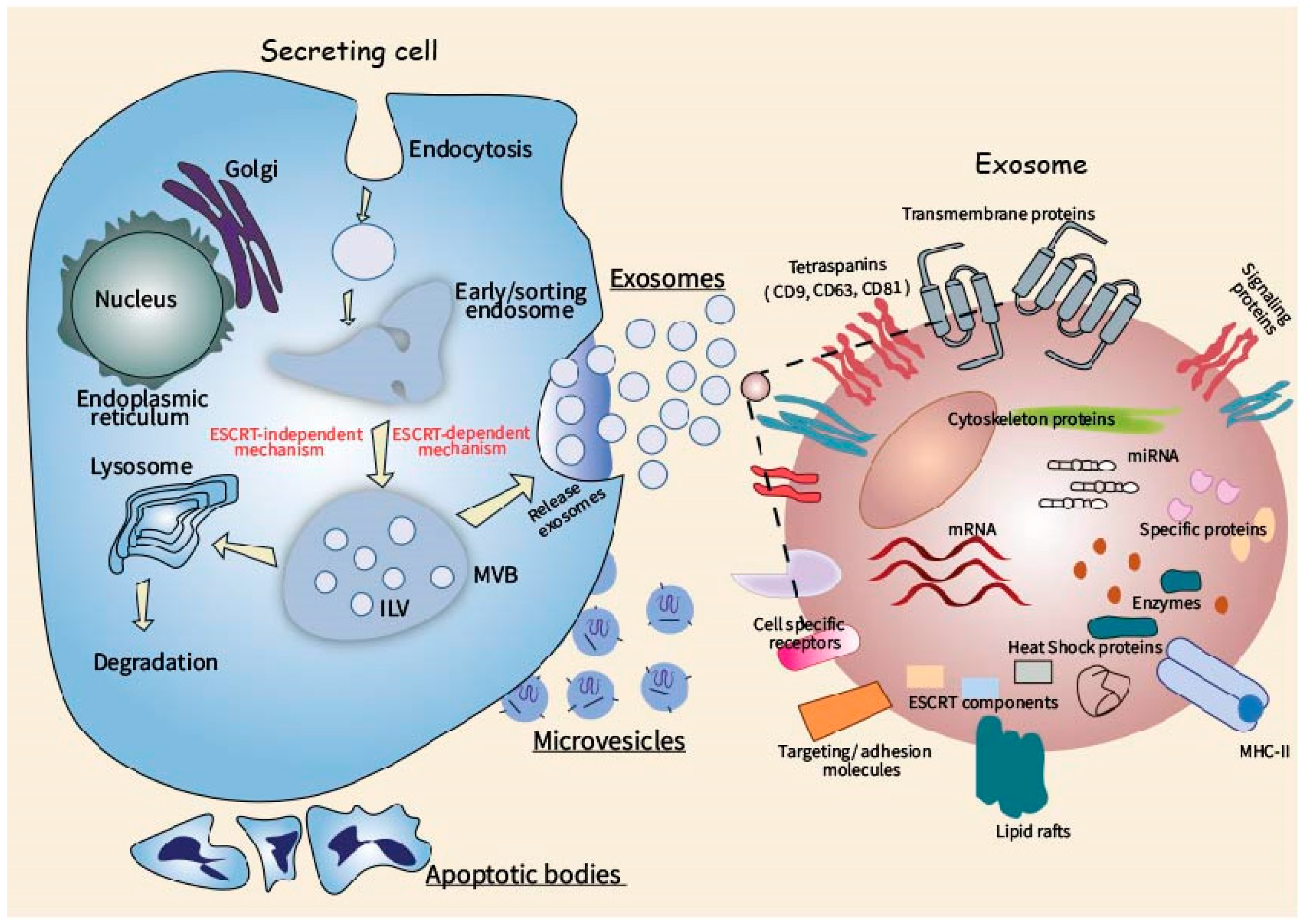{kind=link}
Table 1.
Molecular components of extracellular vesicles.
| Proteins | Lipids | Nucleic Acids | |
|---|---|---|---|
| Exosomes | Main ESCRT proteins TSG101 [35] Alix [52] Rab proteins [53] Annexin A2 [54] Target cell recognition and fusion proteins tetraspanins CD9, CD63, CD81 [55] integrins and connexins [56] Cytoskeletal proteins actin, tubulin, cofilin [46] Heat shock proteins HSP60, HSP70, HSP90 [57] Proteins regulating immune responses Fas ligand (FasL), tumor necrosis factor alpha (TNF-α), transforming growth factor beta (TGF-β) [58] Enzymes Phosphoglycerate kinase 1 (PGK1), glyceraldehyde3-phosphate dehydrogenase (GAPDH) [55] | Higher content then in cells Cholesterol Ceramides Sphingomyelin Glycosphingolipids Phosphatidylserine (PS) [59] Similar content Phosphatidylethanolamine (PE) Lower content then in cells Phosphatidylcholine (PC) Phosphatidylinositol (PI) Diacylglycerol [60,61] +Biologically active lipids Prostaglandins, lysophospholipids [62] | mRNAs non-coding RNAs microRNAs [63,64,65,66] Genomic and mitochondrial DNA [67] tRNA fragments [68] |
| Microvesicles | Membrane proteins Integrins and flotilin-1, matrix metalloproteinase MT1-MMP, glycoprotein receptors (GP1b and GPIIb/GPIIa), adhesion protein P-selectin [69] | Cholesterol, sphingomyelin, ceramide [70], Phosphatidylserine Polyunsaturated lipids [71] | miRNAs, mRNAs non-coding RNAs [72], genomic and mitochondrial DNA [73] |
| Apoptotic bodies | Marker proteins Annexin V [74] Thrombospondin C3b Cytoplasmic proteins | Limited by the plasma membrane | rRNAs [24] Genomic and mitochondrial DNA [75] |
Table 2.
The main components of extracellular vesicles, secreted from cells infected by different viruses.
Table 2.
The main components of extracellular vesicles, secreted from cells infected by different viruses.
| Herpes simplex virus (HSV-1) | Viral tegument proteins and other glycoproteins [92] Viral glycoprotein B [93] |
| Epstein-Barr virus (EBV) | LMP1 [94,95] Galestine 9 [96,97] Epidermal growth factor receptor (EGFR) [98] Phosphatidylinositol-3-kinase (PI3K) [99] Fibroblast growth factor (FGF2) [100] HIF1-alpha [101] Viral RNAs (BHRF1, BART1, and BART2) [102] |
| Human Immunodeficiency Virus (HIV) | Gag proteins [103,104] Nef protein [105] HIV coreceptors CCR5 and CXCR4 [106,107] Transactivation response element (TAR) [108] |
| Hepatitis C virus (HCV) | Viral E1 and E2 proteins [109] Viral RNA [110] |
| Hepatitis A (HAV) | Viral particles [111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Purvinsh, L.; Gorshkov, A.; Brodskaia, A.; Vasin, A. Extracellular Vesicles in Viral Pathogenesis: A Case of Dr. Jekyll and Mr. Hyde. Life 2021, 11, 45. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010045
AMA Style
Purvinsh L, Gorshkov A, Brodskaia A, Vasin A. Extracellular Vesicles in Viral Pathogenesis: A Case of Dr. Jekyll and Mr. Hyde. Life. 2021; 11(1):45. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010045
Chicago/Turabian StylePurvinsh, Lada, Andrey Gorshkov, Aleksandra Brodskaia, and Andrey Vasin. 2021. "Extracellular Vesicles in Viral Pathogenesis: A Case of Dr. Jekyll and Mr. Hyde" Life 11, no. 1: 45. https://0-doi-org.brum.beds.ac.uk/10.3390/life11010045
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.