
Multidrug-Resistant Acinetobacter baumannii Genetic Characterization and Spread in Lithuania in 2014, 2016, and 2018

Abstract
:1. Introduction
2. Materials and Methods
2.1. Acinetobacter spp. Isolates Collection, Identification, and Genomic DNA Extraction
2.2. Characterization of Acinetobacter spp. Antibiotic Resistance Genes
2.3. Genotyping of Acinetobacter spp. Isolates
2.4. Data Analysis
3. Results
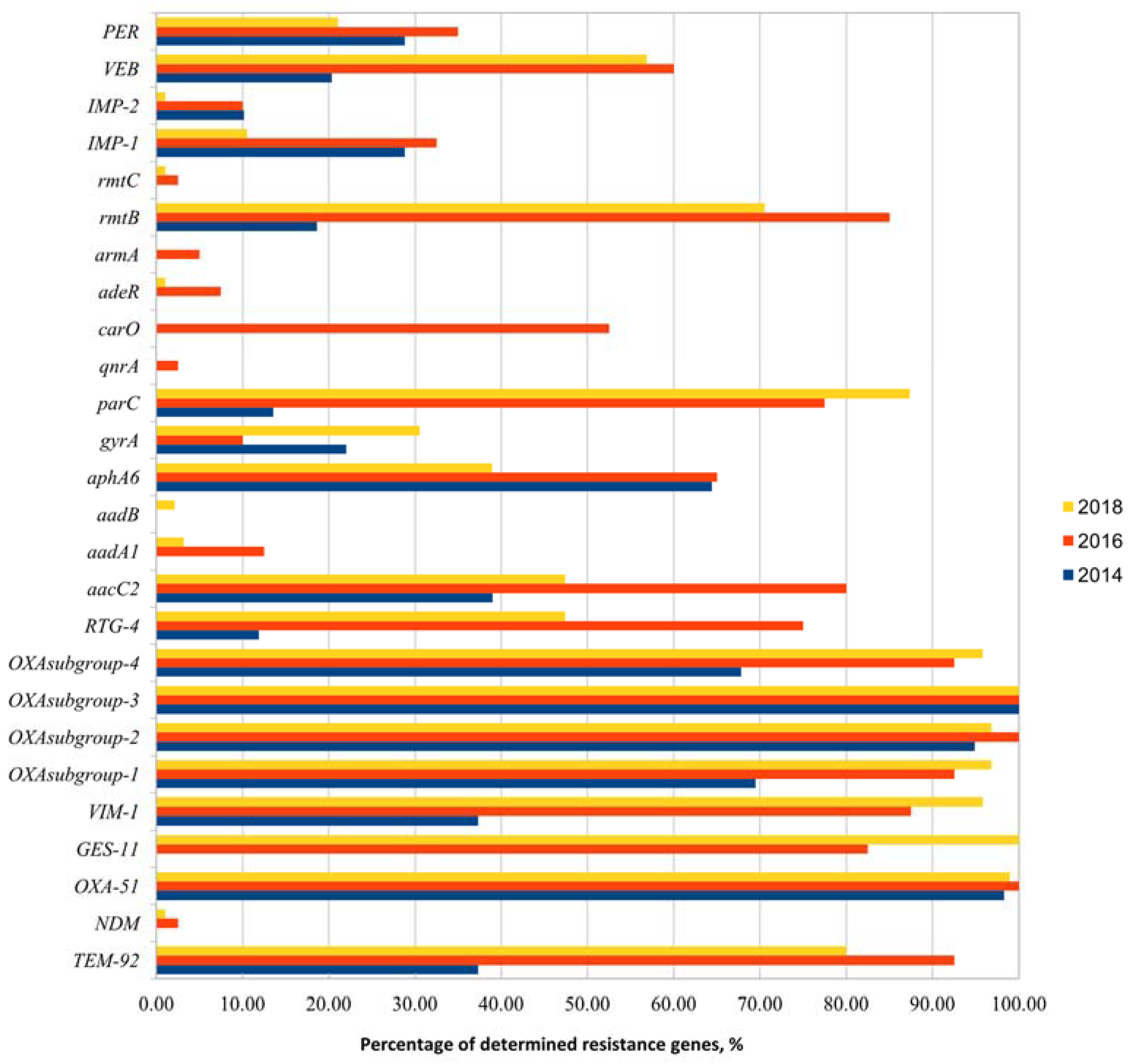
3.1. Acinetobacter spp. Identification, Phenotypic Characterization, and Detection of Antibiotic Resistance Genes
3.2. Rep-PCR and MLVA Genotyping Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Prior, K.; Harmsen, D.; Seifert, H. Development and evaluation of a core genome multilocus typing scheme for whole-genome sequence-based typing of Acinetobacter baumannii. PLoS ONE 2017, 12, e0179228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, Y.; Soler, C.; Jault, P.; Mérens, A.; Gérome, P.; Mac Nab, C.; Trueba, F.; Bargues, L.; Thien, H.V.; Vergnaud, G.; et al. Diversity of Acinetobacter baumannii in Four French Military Hospitals, as Assessed by Multiple Locus Variable Number of Tandem Repeats Analysis. PLoS ONE 2012, 7, e44597. [Google Scholar] [CrossRef]
- Hujer, K.M.; Hujer, A.M.; Hulten, E.A.; Bajaksouzian, S.; Adams, J.M.; Donskey, C.J.; Thomson, J.M. Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian pa-tients treated at the Walter Reed Army Medical Center. Antimicrob. Agents Chemother. 2006, 50, 4114–4123. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Arakawa, Y. 16S ribosomal RNA methylation: Emerging resistance mechanism against aminoglyco-sides. Clin. Infect. Dis. 2007, 45, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Rodriguez-Martinez, J.-M.; Mammeri, H.; Liard, A.; Nordmann, P. Origin of Plasmid-Mediated Quinolone Resistance Determinant QnrA. Antimicrob. Agents Chemother. 2005, 49, 3523–3525. [Google Scholar] [CrossRef] [Green Version]
- Pasterán, F.; Vázquez, M.; Tokumoto, M.; Rapoport, M.; Petroni, A.; Faccone, D.; Corso, A.; Galas, M.; Procopio, A.; Cagnoni, V. Emergence of PER-2 and VEB-1a in Acinetobacter baumannii Strains in the Americas. Antimicrob. Agents Chemother. 2006, 50, 3222–3224. [Google Scholar] [CrossRef] [Green Version]
- Moubareck, C.A.; Brémont, S.; Conroy, M.-C.; Courvalin, P.; Lambert, T. GES-11, a Novel Integron-Associated GES Variant in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3579–3581. [Google Scholar] [CrossRef] [Green Version]
- Endimiani, A.; Luzzaro, F.; Migliavacca, R.; Mantengoli, E.; Hujer, A.M.; Hujer, K.M.; Pagani, L.; Bonomo, R.A.; Rossolini, G.M.; Toniolo, A. Spread in an Italian Hospital of a Clonal Acinetobacter baumannii Strain Producing the TEM-92 Extended-Spectrum β-Lactamase. Antimicrob. Agents Chemother. 2007, 51, 2211–2214. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Poirel, L.; Croizé, J.; Chanteperdrix, V.; Nordmann, P. Genetic and Biochemical Characterization of the First Extended-Spectrum CARB-Type ß-Lactamase, RTG-4, from Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3010–3016. [Google Scholar] [CrossRef] [Green Version]
- Gales, A.C.; Menezes, L.C.; Silbert, S.; Sader, H.S. Dissemination in distinct Brazilian regions of an epidemic carbapenem-resistant Pseudomonas aeruginosa producing SPM metallo-β-lactamase. J. Antimicrob. Chemother. 2003, 52, 699–702. [Google Scholar] [CrossRef]
- Tsakris, A.; Pournaras, S.; Woodford, N.; Palepou, M.F.I.; Babini, G.S.; Douboyas, J.; Livermore, D.M. Out-break of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J. Clin. Microbiol. 2000, 38, 1290–1292. [Google Scholar] [CrossRef] [Green Version]
- Shokri, D.; Khorasgani, M.R.; Fatemi, S.M.; Soleimani-Delfan, A. Resistotyping, phenotyping and genotyping of New Delhi metallo-β-lactamase (NDM) among Gram-negative bacilli from Iranian patients. J. Med. Microbiol. 2017, 66, 402–411. [Google Scholar] [CrossRef]
- Turton, J.F.; Ward, M.E.; Woodford, N.; Kaufmann, M.E.; Pike, R.; Livermore, D.M.; Pitt, T.L. The role of IS Aba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett. 2006, 258, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dombek, P.E.; Johnson, L.K.; Zimmerley, S.T.; Sadowsky, M.J. Use of Repetitive DNA Sequences and the PCR To Differentiate Escherichia coli Isolates from Human and Animal Sources. Appl. Environ. Microbiol. 2000, 66, 2572–2577. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Minandri, F.; Hauck, Y.; d’Arezzo, S.; Imperi, F.; Vergnaud, G.; Visca, P. Identification of varia-ble-number tandem-repeat (VNTR) sequences in Acinetobacter baumannii and interlaboratory validation of an opti-mized multiple-locus VNTR analysis typing scheme. J. Clin. Microbiol. 2011, 49, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafei, R.; Osman, M.; Dabboussi, F.; Hamze, M. Update on the epidemiological typing methods for Acinetobacter baumannii. Future Microbiol. 2019, 14, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Bergogne-Berezin, E.; Towner, K.J. Acinetobacter spp. as nosocomial pathogens: Microbiological, clinical, and epidemiological features. Clin. Microbiol. Rev. 1996, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Galimand, M.; Courvalin, P.; Lambert, T. Plasmid-mediated high-level resistance to aminoglycosides in Enterobacteriaceae due to 16S rRNA methylation. Antimicrob. Agents Chemother. 2003, 47, 2565–2571. [Google Scholar] [CrossRef] [Green Version]
- Yamane, K.; Wachino, J.-I.; Suzuki, S.; Shibata, N.; Kato, H.; Shibayama, K.; Kimura, K.; Kai, K.; Ishikawa, S.; Ozawa, Y.; et al. 16S rRNA Methylase–producing, Gram-Negative Pathogens, Japan. Emerg. Infect. Dis. 2007, 13, 642–646. [Google Scholar] [CrossRef]
- Wachino, J.I.; Arakawa, Y. Exogenously acquired 16S rRNA methyltransferases found in aminoglyco-side-resistant pathogenic Gram-negative bacteria: An update. Drug Resist. Updates 2012, 15, 133–148. [Google Scholar] [CrossRef]
- Aliakbarzade, K.; Farajnia, S.; Nik, A.K.; Zarei, F.; Tanomand, A. Prevalence of Aminoglycoside Resistance Genes in Acinetobacter baumannii Isolates. Jundishapur J. Microbiol. 2014, 7, e11924. [Google Scholar] [CrossRef] [Green Version]
- Moniri, R.; Farahani, R.K.; Shajari, G.; Shirazi, M.N.; Ghasemi, A. Molecular epidemiology of aminoglyco-sides resistance in Acinetobacter spp. With emergence of multidrug-resistant strains. Iran. J. Public Health 2010, 39, 63. [Google Scholar] [PubMed]
- Nigro, S.J.; Hall, R.M. Loss and gain of aminoglycoside resistance in global clone 2 Acinetobacter baumannii in Australia via modification of genomic resistance islands and acquisition of plasmids. J. Antimicrob. Chemother. 2016, 71, 2432–2440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, T.K.M.; Chu, Y.W.; Chu, M.Y.; Ma, C.H.; Yung, R.W.H.; Kam, K.M. Plasmid-mediated resistance to ciprofloxacin and cefotaxime in clinical isolates of Salmonella enterica serotype Enteritidis in Hong Kong. J. Antimicrob. Chemother. 2005, 56, 586–589. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Cuenca, F.; Martínez-Martínez, L.; Conejo, M.C.; Ayala, J.A.; Perea, E.J.; Pascual, A. Relationship between beta-lactamase production, outer membrane protein and penicillin-binding protein profiles on the activity of carbapenems against clinical isolates of Acinetobacter baumannii. J. Antimicrob. Chemother. 2003, 51, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touati, A.; Brasme, L.; Benallaoua, S.; Gharout, A.; Madoux, J.; De Champs, C. First report of qnrB-producing Enterobacter cloacae and qnrA-producing Acinetobacter baumannii recovered from Algerian hospitals. Diagn. Microbiol. Infect. Dis. 2008, 60, 287–290. [Google Scholar] [CrossRef]
- Povilonis, J.; Šeputienė, V.; Krasauskas, R.; Juškaitė, R.; Miškinytė, M.; Sužiedėlis, K.; Sužiedėlienė, E. Spread of carbapenem-resistant Acinetobacter baumannii carrying a plasmid with two genes encoding OXA-72 carbapenemase in Lithuanian hospitals. J. Antimicrob. Chemother. 2013, 68, 1000–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héritier, C.; Poirel, L.; Lambert, T.; Nordmann, P. Contribution of acquired carbapenem-hydrolyzing oxacil-linases to carbapenem resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 3198–3202. [Google Scholar] [CrossRef] [Green Version]
- Chmielarczyk, A.; Pilarczyk-Zurek, M.; Kamińska, W.; Pobiega, M.; Romaniszyn, D.; Ziółkowski, G.; Wojkowska-Mach, J.; Bulanda, M. Molecular Epidemiology and Drug Resistance of Acinetobacter baumannii Isolated from Hospitals in Southern Poland: ICU as a Risk Factor for XDR Strains. Microb. Drug Resist. 2016, 22, 328–335. [Google Scholar] [CrossRef]
- Noori, M.; Karimi, A.; Fallah, F.; Hashemi, A.; Alimehr, S.; Goudarzi, H.; Aghamohammad, S. High Prevalence of Metallo-beta-lactamase Producing Acinetobacter. Arch. Pediatr. 2014, 2, e15439. [Google Scholar]
- Amin, M.; Navidifar, T.; Shooshtari, F.S.; Goodarzi, H. Association of the genes encoding Metallo-β-Lactamase with the presence of integrons among multidrug-resistant clinical isolates of Acinetobacter baumannii. Infect. Drug Resist. 2019, 12, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Bilya, S.; Xu, W. adeABC efflux gene in Acinetobacter baumannii. New Microbes New Infect. 2019, 30, 100549. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-Type Efflux Pump AdeABC in Acinetobacter baumannii Is Regulated by the AdeRS Two-Component System. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanin, P.; Palermo, R.L.; Cavalini, J.F.; Fávaro, L.D.S.; De Paula-Petroli, S.B.; Fernandes, E.V.; Yamauchi, L.M. Multidrug-and extensively drug-resistant Acinetobacter baumannii in a tertiary hospital from Brazil: The importance of carbapenemase encoding genes and epidemic clonal complexes in a 10-year study. Microb. Drug Resist. 2019, 25, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Koczura, R.; Przyszlakowska, B.; Mokracka, J.; Kaznowski, A. Class 1 Integrons and Antibiotic Resistance of Clinical Acinetobacter calcoaceticus–baumannii Complex in Poznań, Poland. Curr. Microbiol. 2014, 69, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, S.; Kim, M.-J.; Yun, S.-J.; Moon, J.Y.; Kim, Y.-S. Controlling endemic multidrug-resistant Acinetobacter baumannii in Intensive Care Units using antimicrobial stewardship and infection control. Korean J. Intern. Med. 2016, 31, 367–374. [Google Scholar] [CrossRef]
- Kirtikliene, T.; Naugzemys, D.; Steponkiene, A.; Bogdevic, R.; Vizuje, G.; Zvingila, D.; Kuisiene, N. Evaluation of the Inter- and Intrahospital Spread of Multidrug Resistant Gram-Negative Bacteria in Lithuanian Hospitals. Microb. Drug Resist. 2019, 25, 326–335. [Google Scholar] [CrossRef]
- Johnson, J.K.; Robinson, G.L.; Zhao, L.; Harris, A.D.; Stine, O.C.; Thom, K.A. Comparison of molecular typing methods for the analyses of Acinetobacter baumannii from ICU patients. Diagn. Microbiol. Infect. Dis. 2016, 86, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, B.; Jin, D.; Cui, Z.; Tao, X.; Zhang, B.; Zhang, J. Comparison of multiple-locus variable-number tan-dem-repeat analysis with pulsed-field gel electrophoresis typing of Acinetobacter baumannii in China. J. Clin. Microbiol. 2013, 51, 1263–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkshoorn, L. Acinetobacter baumannii. In Molecular Typing in Bacterial Infections; Humana Press: Totowa, NJ, USA, 2013; pp. 433–456. [Google Scholar]
- Saffari, F.; Monsen, T.; Karmostaji, A.; Azimabad, F.B.; Widerström, M. Significant spread of extensively drug-resistant Acinetobacter baumannii genotypes of clonal complex 92 among intensive care unit patients in a university hospital in southern Iran. J. Med. Microbiol. 2017, 66, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Najar-Peerayeh, S.; Karmostaji, A. Evaluation of Multilocus Variable-Number Tandem-Repeat (MLVA-8 Or-say) for Typing of Carbapenem-Resistant Acinetobacter baumannii Isolated from Patients in Tehran, Iran. Arch. Clin. Infect. Dis. 2019, 14, e64402. [Google Scholar]


{kind=link}
| Resistance Gene | Resistance Mechanism | Reference |
|---|---|---|
| aacC1 | Amynoglycoside-modifying enzymes (AMEs) | [5] |
| aacC2 | ||
| aadB | ||
| aadA1 | ||
| aphA6 | ||
| rmtB | [6] | |
| rmtC | ||
| armA | ||
| gyrA | Fluoroquinolone resistance-determining regions (QRDR) | [5] |
| parC | ||
| qnrA | ||
| adeR | Efflux pumps regulation genes | [5] |
| adeE | ||
| blaPER-1 | β-lactams resistance genes | [7] |
| blaVEB-1 | [8] | |
| blaGES-11 | [9] | |
| blaTEM-92 | [10] | |
| blaRTG-4 | [11] | |
| blaIMP-1 | [12] | |
| blaIMP-2 | ||
| blaVIM-1 | [13] | |
| blaNDM | [14] | |
| blaOXA subgroup-1 (blaOXA-23, blaOXA-27, blaOXA-49) | [8] | |
| blaOXA subgroup-2 (blaOXA-24, blaOXA-25, blaOXA-26, blaOXA-40, blaOXA-72) | ||
| blaOXA subgroup-3 (blaOXA-51, blaOXA-64, blaOXA-65, blaOXA-66, blaOXA-68, blaOXA-69, blaOXA-70, blaOXA-71, blaOXA-75, blaOXA-76, blaOXA-77, blaOXA-78 ) | ||
| blaOXA subgroup-4 (blaOXA-58) | ||
| blaOXA-51 | [15] | |
| blacarO | [5] |
| Cluster | 2014 Year: Number of Profiles | 2016 Year: Number of Profiles | 2018 Year: Number of Profiles | Number of Healthcare Institutions | Numbers of Profiles in Healthcare Institutions (Number of Profiles) |
|---|---|---|---|---|---|
| Cluster 1 | 14 | 1 | 1 | 9 | A (11), D (1), E (1), F (1), G (1), H (8), K (1), M (1), N (1) |
| Cluster 2 | 5 | 1 | 27 | 8 | A (12), C (2), D (3), F (1), G (2), H (8), I (1), K (2), one A/H profile, one A/G profile |
| Cluster 3 | - | - | 20 | 7 | A (9), C (2), D (1), E (1), F (1), H (4), I (2) |
| Cluster 4 | 11 | 22 | 2 | 10 | A (16), B (1), D (1), E (2), F (1), H (7), I (2), K (1), M (2), N (1) |
| Cluster 5 | 8 | 9 | 13 | 8 | A (15), D (2), F (1), J (1), H (4), K (3), L (1), M (3) |
| Cluster 6 | 20 | 6 | 20 | 9 | A (19), B (1), C (1), D (1), F (1), G (1), H (11), I (2), M (7), one A/C profile |
| Profile Number | Isolate | Healthcare Institution | Number of Resistance Genes Detected | Genes in Both Isolates | Different Genes in Isolates |
|---|---|---|---|---|---|
| 1 | 201429 | A | 7 | blaOXA-51, blaVIM-1, blaOXA subgroup-2, blaOXA subgroup-3 | blaaacC2, blaaphA6, blaIMP-1 |
| 2014212 | G | 8 | blaTEM-92, blaOXA subgroup-1, blaaacC1, blaarmA | ||
| 2 | 2018364 | A | 12 | blaTEM-92, blaOXA-51, blaGES-11, blaVIM-1, blaOXA subgroups 1-4, blaparC, blaVEB | blagyrA, blaPER |
| 2018444 | H | 11 | blaaacC2 | ||
| 3 | 2018391 | C | 12 | blaTEM-92, blaOXA-51, blaGES-11, blaVIM-1, blaOXA subgroups 1-4, blaRTG-4, blaaacC2, blaparC, blarmtB | - |
| 2018419 | A | 14 | blaaphA6, blaPER |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirtikliene, T.; Mierauskaitė, A.; Razmienė, I.; Kuisiene, N. Multidrug-Resistant Acinetobacter baumannii Genetic Characterization and Spread in Lithuania in 2014, 2016, and 2018. Life 2021, 11, 151. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020151
Kirtikliene T, Mierauskaitė A, Razmienė I, Kuisiene N. Multidrug-Resistant Acinetobacter baumannii Genetic Characterization and Spread in Lithuania in 2014, 2016, and 2018. Life. 2021; 11(2):151. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020151
Chicago/Turabian StyleKirtikliene, Tatjana, Aistė Mierauskaitė, Ilona Razmienė, and Nomeda Kuisiene. 2021. "Multidrug-Resistant Acinetobacter baumannii Genetic Characterization and Spread in Lithuania in 2014, 2016, and 2018" Life 11, no. 2: 151. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020151