
Proteomic Insight into the Symbiotic Relationship of Pinus massoniana Lamb and Suillus luteus towards Developing Al-Stress Resistance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Ectomycorrhizal Fungus
2.2. Collection, Separation, and Purification of Suillus Luteus
2.3. Identification of Suillus Luteus Strain
2.4. Stress Treatments
2.5. Protein Extraction
2.6. LC-MS/MS Analysis
2.7. Functional Classification of Proteins
2.8. Protein Quantification and Differential Expressed Protein Analysis
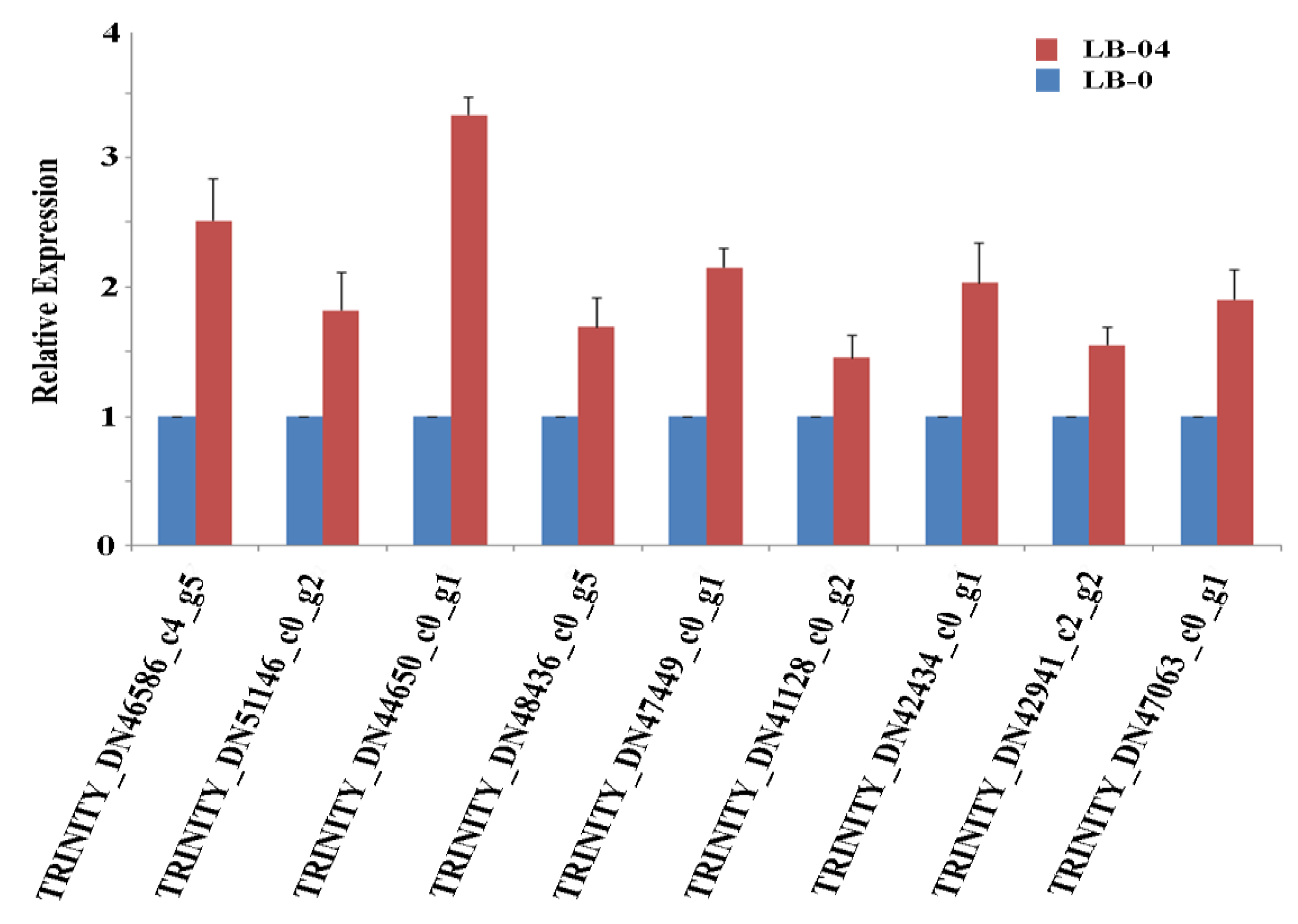
2.9. Expression Profile of Related Genes based on qRT-PCR
3. Results
3.1. Overview of the Proteome Profiling in Masson Pine Needle Samples
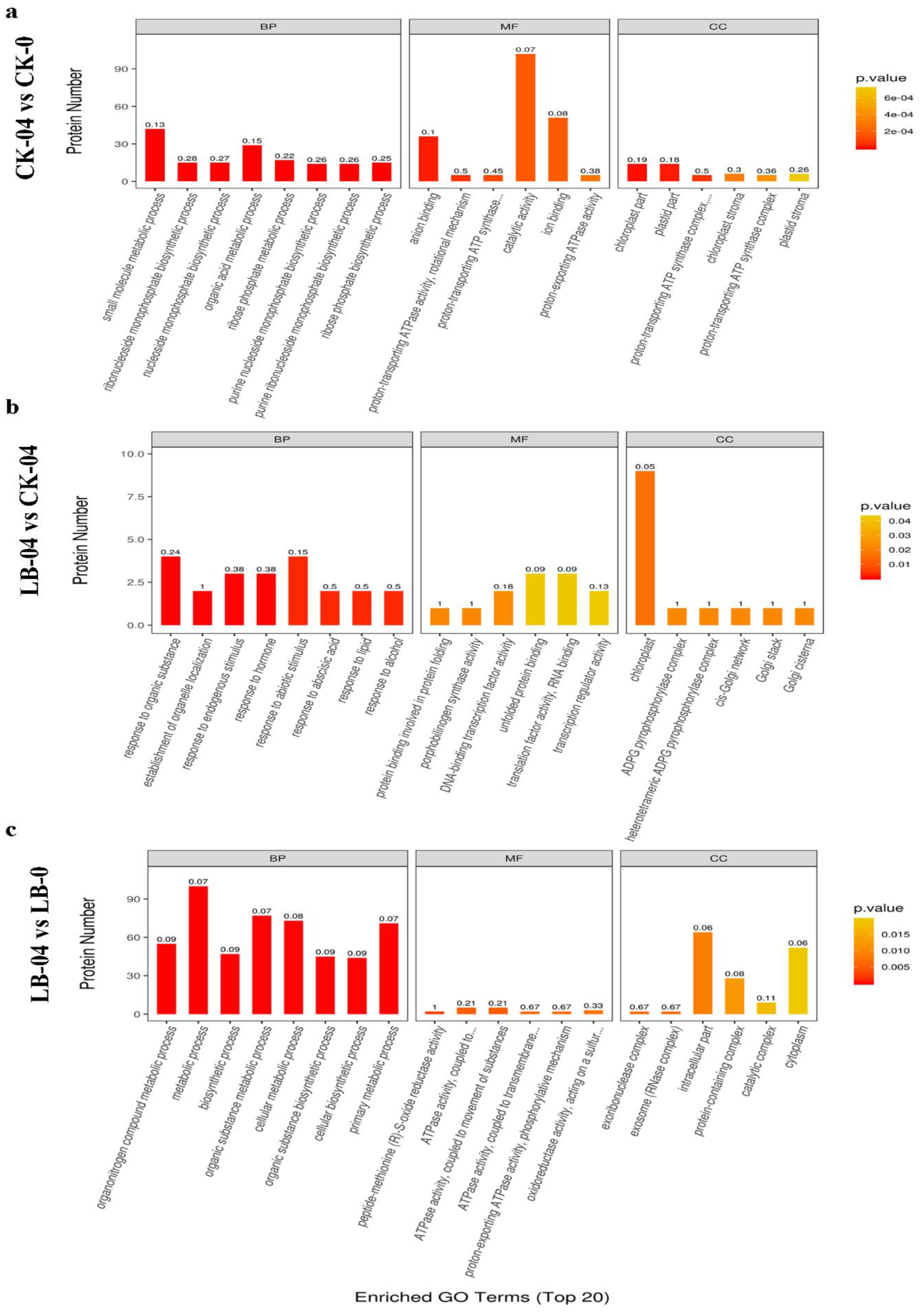
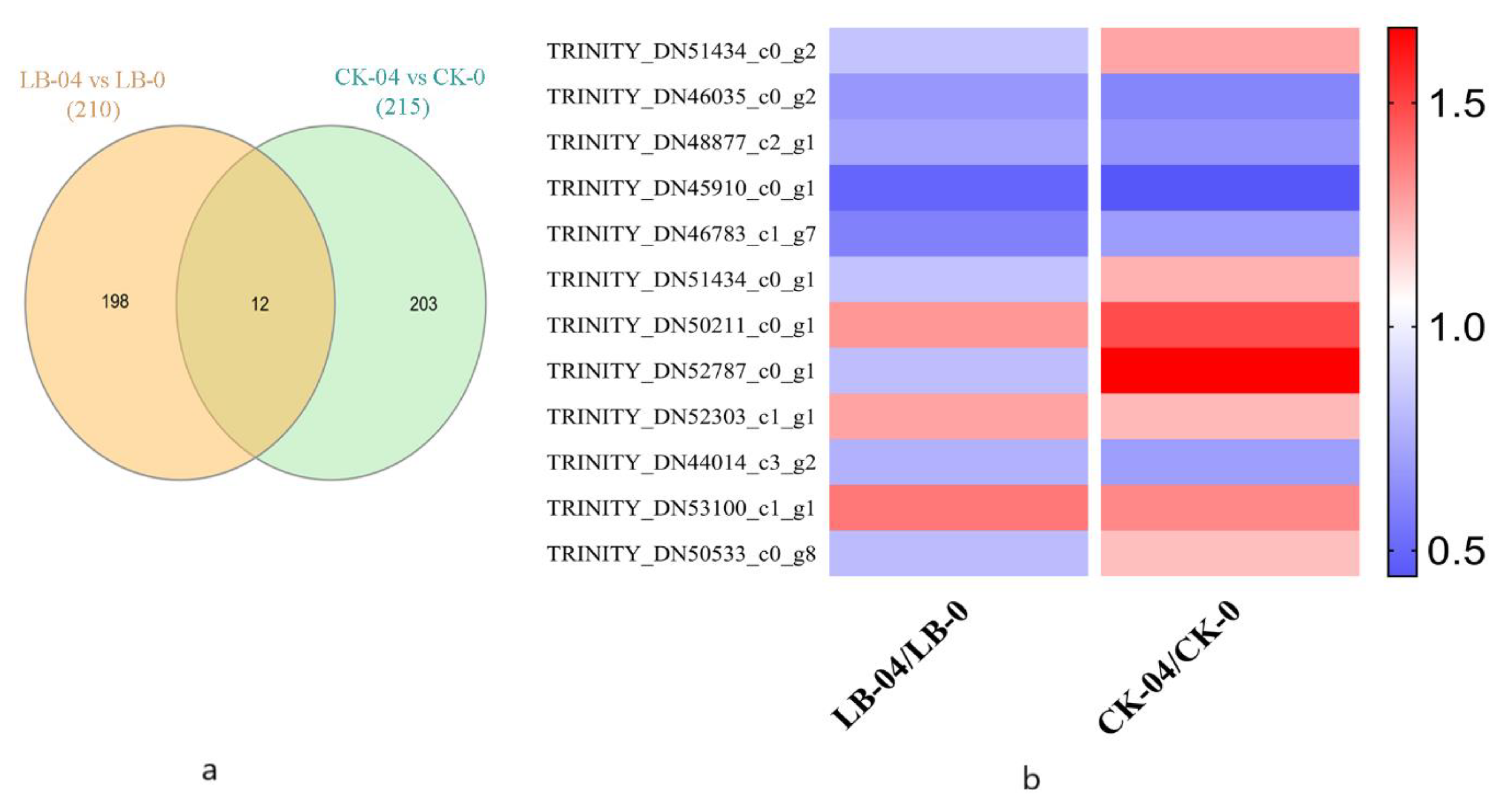
3.2. Differentially Expressed Proteins Analysis and Functional Annotations
3.3. Identification and Analysis of the Core Al Responsive Proteins
3.4. Identification and Analysis of Specific Proteins involved in Al Response under S. luteus Inoculation in Masson Pine Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.-C.; Liang, M.-J.; Wang, S.-L. Carbon stock density in planted versus natural Pinus massoniana forests in sub-tropical China. Ann. For. Sci. 2016, 73, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.; Deng, Q.; Li, M.; Wang, W.; Zhang, Q.; Cheng, X. Reforestation of Pinus massoniana alters soil organic carbon and nitrogen dynamics in eroded soil in south China. Ecol. Eng. 2013, 52, 154–160. [Google Scholar] [CrossRef]
- Yi, W.; Guijie, D. Physiological responses of mycorrhizal Pinus massoniana seedlings to drought stress and drought resistance evaluation. Yingyong Shengtai Xuebao 2013, 24, 639–645. [Google Scholar]
- Meng, J.; Lu, Y.; Zeng, J. Transformation of a degraded Pinus massoniana plantation into a mixed-species irregular forest: Impacts on stand structure and growth in southern China. Forests 2014, 5, 3199–3221. [Google Scholar] [CrossRef]
- Zhang, T.; Wen, X.-P.; Ding, G.-J. Ectomycorrhizal symbiosis enhances tolerance to low phosphorous through expression of phosphate transporter genes in masson pine (Pinus massoniana). Acta Physiol. Plant. 2017, 39, 101. [Google Scholar] [CrossRef]
- Laganiere, J.; Angers, D.A.; Pare, D. Carbon accumulation in agricultural soils after afforestation: A meta-analysis. Glob. Chang. Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Rincón, A.; Alvarez, I.F.; Pera, J. Inoculation of containerized Pinus pinea L. seedlings with seven ectomycorrhizal fungi. Mycorrhiza 2001, 11, 265–271. [Google Scholar] [CrossRef]
- Parladé, J.; Pera, J.; Alvarez, I.F. Inoculation of containerized Pseudotsuga menziesii and Pinus pinaster seedlings with spores of five species of ectomycorrhizal fungi. Mycorrhiza 1996, 6, 237–245. [Google Scholar] [CrossRef]
- Lu, N.; Yu, M.; Cui, M.; Luo, Z.; Feng, Y.; Cao, S.; Sun, Y.; Li, Y. Effects of different ectomycorrhizal fungal inoculates on the growth of Pinus tabulaeformis seedlings under greenhouse conditions. Forests 2016, 7, 316. [Google Scholar] [CrossRef] [Green Version]
- Rincón, A.; Álvarez, I.F.; Pera, J. Ectomycorrhizal fungi of Pinus pinea L. in northeastern Spain. Mycorrhiza 1999, 8, 271–276. [Google Scholar] [CrossRef]
- Luo, Z.-B.; Janz, D.; Jiang, X.; Goebel, C.; Wildhagen, H.; Tan, Y.; Rennenberg, H.; Feussner, I.; Polle, A. Upgrading root physiology for stress tolerance by ectomycorrhizas: Insights from metabolite and transcriptional profiling into reprogramming for stress anticipation. Plant Physiol. 2009, 151, 1902–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Z.; Xie, W.; Chen, B. Arbuscular mycorrhizal symbiosis affects plant immunity to viral infection and accumulation. Viruses 2019, 11, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2011, 21, 71–90. [Google Scholar] [CrossRef]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Magalhaes, J.V. Aluminum tolerance genes are conserved between monocots and dicots. Proc. Natl. Acad. Sci. USA 2006, 103, 9749–9750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, S.R.; Carrasco, J.; Pinochet, D.; Calderini, D.F. Grain yield, above-ground and root biomass of Al-tolerant and Al-sensitive wheat cultivars under different soil aluminum concentrations at field conditions. Plant Soil 2009, 318, 299–310. [Google Scholar] [CrossRef]
- Duressa, D.; Soliman, K.; Taylor, R.; Senwo, Z. Proteomic analysis of soybean roots under aluminum stress. Int. J. Plant Genom. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Rout, G.; Samantaray, S.; Das, P. Aluminium toxicity in plants: A review. Agronomie 2001, 21, 3–21. [Google Scholar] [CrossRef]
- Roy, A.K.; Sharma, A.; Talukder, G. Some aspects of aluminum toxicity in plants. Bot. Rev. 1988, 54, 145–178. [Google Scholar] [CrossRef]
- Godbold, D.; Fritz, E.; Hüttermann, A. Aluminum toxicity and forest decline. Proc. Natl. Acad. Sci. USA 1988, 85, 3888–3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, N.R.; Franco, A.R.; Oliveira, R.S.; Castro, P.M. Ectomycorrhizal fungi as an alternative to the use of chemical fertilisers in nursery production of Pinus pinaster. J. Environ. Manag. 2012, 95, S269–S274. [Google Scholar] [CrossRef]
- Dixon, R.; Buschena, C. Response of ectomycorrhizal Pinus banksiana and Picea glauca to heavy metals in soil. Plant Soil 1988, 105, 265–271. [Google Scholar] [CrossRef]
- Meharg, A.; Cairney, J.W. Co-evolution of mycorrhizal symbionts and their hosts to metal-contaminated environments. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1999; Volume 30, pp. 69–112. [Google Scholar]
- Jentschke, G.; Godbold, D. Metal toxicity and ectomycorrhizas. Physiol. Plant. 2000, 109, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Van Tichelen, K.K.; Vanstraelen, T.; Colpaert, J.V. Nutrient uptake by intact mycorrhizal Pinus sylvestris seedlings: A diagnostic tool to detect copper toxicity. Tree Physiol. 1999, 19, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adriaensen, K.; Van Der Lelie, D.; Van Laere, A.; Vangronsveld, J.; Colpaert, J.V. A zinc-adapted fungus protects pines from zinc stress. New Phytol. 2004, 161, 549–555. [Google Scholar] [CrossRef]
- Colpaert, J.V.; Vandenkoornhuyse, P.; Adriaensen, K.; Vangronsveld, J. Genetic variation and heavy metal tolerance in the ectomycorrhizal basidiomycete Suillus luteus. New Phytol. 2000, 147, 367–379. [Google Scholar] [CrossRef]
- Krznaric, E.; Verbruggen, N.; Wevers, J.H.; Carleer, R.; Vangronsveld, J.; Colpaert, J.V. Cd-tolerant Suillus luteus: A fungal insurance for pines exposed to Cd. Environ. Pollut. 2009, 157, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yuan, L.; Zhou, Z.; Yang, H.; Huang, J. Efflux of oxalate and uptake of nitrogen, phosphorus and potassium by ectomycorrhizal fungal isolates in vitro in response to aluminum stress. Sci. Silvae Sin. 2012, 48, 82–88. [Google Scholar]
- Rudawska, M.; Kieliszewska-Rokicka, B.; Leski, T. Effect of aluminium on Pinus sylvestris seedlings mycorrhizal with aluminium-tolerant and aluminium-sensitive strains of Suillus luteus. Dendrobiology 2000, 45, 89–96. [Google Scholar]
- Kong, F.; Liu, Y.; Hu, W.; Shen, P.; Zhou, C.; Wang, L. Biochemical responses of the mycorrhizae in Pinus massoniana to combined effects of Al, Ca and low pH. Chemosphere 2000, 40, 311–318. [Google Scholar] [CrossRef]
- Chen, L.-S.; Qi, Y.-P.; Liu, X.-H. Effects of aluminum on light energy utilization and photoprotective systems in citrus leaves. Ann. Bot. 2005, 96, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Basu, U.; Southron, J.; Stephens, J.; Taylor, G. Reverse genetic analysis of the glutathione metabolic pathway suggests a novel role of PHGPX and URE2 genes in aluminum resistance in Saccharomyces cerevisiae. Mol. Genet. Genom. 2004, 271, 627–637. [Google Scholar] [CrossRef]
- Shen, H.; He, L.F.; Sasaki, T.; Yamamoto, Y.; Zheng, S.J.; Ligaba, A.; Yan, X.L.; Ahn, S.J.; Yamaguchi, M.; Sasakawa, H. Citrate secretion coupled with the modulation of soybean root tip under aluminum stress. Up-regulation of transcription, translation, and threonine-oriented phosphorylation of plasma membrane H+-ATPase. Plant Physiol. 2005, 138, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, C.A.; Good, A.G.; Taylor, G.J. Induction of vacuolar ATPase and mitochondrial ATP synthase by aluminum in an aluminum-resistant cultivar of wheat. Plant Physiol. 2001, 125, 2068–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, P.B.; Geisler, M.J.; Jones, C.A.; Williams, K.M.; Cancel, J.D. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J. 2005, 41, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Anoop, V.M.; Basu, U.; McCammon, M.T.; McAlister-Henn, L.; Taylor, G.J. Modulation of citrate metabolism alters aluminum tolerance in yeast and transgenic canola overexpressing a mitochondrial citrate synthase. Plant Physiol. 2003, 132, 2205–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, C.; Yi, K.; Yang, L.; Zheng, B.; Wu, Y.; Liu, F.; Wu, P. Identification of aluminium-regulated genes by cDNA-AFLP in rice (Oryza sativa L.): Aluminium-regulated genes for the metabolism of cell wall components. J. Exp. Bot. 2004, 55, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosché, M.; Strid, Å. Cloning, expression, and molecular characterization of a small pea gene family regulated by low levels of ultraviolet B radiation and other stresses. Plant Physiol. 1999, 121, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Sivaguru, M.; Ezaki, B.; He, Z.-H.; Tong, H.; Osawa, H.; Baluška, F.; Volkmann, D.; Matsumoto, H. Aluminum-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003, 132, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Wang, Y.; Zhang, J.; Shi, W.; Qian, C.; Peng, X. Identification of aluminum-responsive proteins in rice roots by a proteomic approach: Cysteine synthase as a key player in Al response. Proteomics 2007, 7, 737–749. [Google Scholar] [CrossRef]
- Zhou, S.; Sauve, R.; Thannhauser, T.W. Aluminum induced proteome changes in tomato cotyledons. Plant Signal. Behav. 2009, 4, 769–772. [Google Scholar] [CrossRef] [Green Version]
- Zhen, Y.; Qi, J.L.; Wang, S.S.; Su, J.; Xu, G.H.; Zhang, M.S.; Miao, L.; Peng, X.X.; Tian, D.; Yang, Y.H. Comparative proteome analysis of differentially expressed proteins induced by Al toxicity in soybean. Physiol. Plant. 2007, 131, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Sun, Y.; Huang, Z.; Zhu, F.; Sun, Y.; Jiang, L. The effects of ectomycorrhizal fungi on heavy metals’ transport in Pinus massoniana and bacteria community in rhizosphere soil in mine tailing area. J. Hazard. Mater. 2020, 381, 121203. [Google Scholar] [CrossRef] [PubMed]
- Haiyan, L.; Houying, C.; Guijie, D.; Kuaifen, L.; Qifei, R. Identification of candidate genes conferring tolerance to aluminum stress in Pinus massoniana inoculated with ectomycorrhizal fungus. BMC Plant Biol. 2020, 20, 1–13. [Google Scholar]
- Fuqiang, Y.; Dagan, J.; Peigui, L. Studies on pure culture of the ectomycorrhizal fungi associated with Pinus yunnanensis. Zhiwu Yanjiu 2003, 23, 66–71. [Google Scholar]
- Gu, J.; Li, R.; ShiGui, J. Recent advance of fungi preservation technology. Mycosystema 2007, 24, 12–25. [Google Scholar]
- Du, P.; Tu, H.; Jiang, J.; Cui, B.; Dai, Y. Molecular Identification and Biological Characteristics of Wild Paxillus ammoniavirescens Strain. For. Sci. Res. 2020, 33, 146–155. [Google Scholar]
- Feng, J.; Wang, M.; YAN, W. Study on Molecular Identification and Genetic Relationships of Ectomycorrhizal Strains of Suillus Gene. For. Res. 2008, 6, 825. [Google Scholar]
- Hothem, S.D.; Marley, K.A.; Larson, R.A. Photochemistry in Hoagland’s nutrient solution. J. Plant Nutr. 2003, 26, 845–854. [Google Scholar] [CrossRef]
- Ghanati, F.; Morita, A.; Yokota, H. Effects of aluminum on the growth of tea plant and activation of antioxidant system. Plant Soil 2005, 276, 133–141. [Google Scholar] [CrossRef]
- Guo, P.; Qi, Y.-P.; Yang, L.-T.; Lai, N.-W.; Ye, X.; Yang, Y.; Chen, L.-S. Root adaptive responses to aluminum-treatment revealed by RNA-Seq in two Citrus species with different aluminum-tolerance. Front. Plant Sci. 2017, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Shen, R.; Zhao, X. Study of different parameters for evaluating Al tolerance in plants. Plant Nutr. Fertil. Sci. 2008, 5, 5. [Google Scholar]
- Xiao, S.; Liu, L.; Zhang, Y.; Sun, H.; Zhang, K.; Bai, Z.; Dong, H.; Liu, Y.; Li, C. Tandem mass tag-based (TMT) quantitative proteomics analysis reveals the response of fine roots to drought stress in cotton (Gossypium hirsutum L.). BMC Plant Biol. 2020, 20, 1–18. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Delhaize, E.; Zhou, M.; Ryan, P.R. Biotechnological solutions for enhancing the aluminium resistance of crop plants. Abiotic Stress Plants—Mech. Adapt. Brisb. Intech 2011, 1, 119–142. [Google Scholar]
- Zhu, J.-J.; Li, F.-Q.; Xu, M.-L.; Kang, H.-Z.; Wu, X.-Y. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China: An experimental approach. Ann. For. Sci. 2008, 65, 1. [Google Scholar] [CrossRef] [Green Version]
- Kipfer, T.; Wohlgemuth, T.; van der Heijden, M.G.; Ghazoul, J.; Egli, S. Growth response of drought-stressed Pinus sylvestris seedlings to single-and multi-species inoculation with ectomycorrhizal fungi. PLoS ONE 2012, 7, e35275. [Google Scholar] [CrossRef]
- Jie, W.; Huang, Y.; JIANG, X.-Y. Influence of ectomycorrhizal fungi on absorption and balance of essential elements of Pinus tabulaeformis seedlings in saline soil. Pedosphere 2011, 21, 400–406. [Google Scholar]
- Yin, D.; Halifu, S.; Song, R.; Qi, J.; Deng, X.; Deng, J. Effects of an ectomycorrhizal fungus on the growth and physiology of Pinus sylvestris var. mongolica seedlings subjected to saline–alkali stress. J. For. Res. 2020, 31, 781–788. [Google Scholar] [CrossRef]
- Kasuya, M.; Muchovej, R.; Muchovej, J. Influence of aluminum on in vitro formation of Pinus caribaea mycorrhizae. Plant Soil 1990, 124, 73–77. [Google Scholar] [CrossRef]
- Ahonen-Jonnarth, U.; Göransson, A.; Finlay, R.D. Growth and nutrient uptake of ectomycorrhizal Pinus sylvestris seedlings in a natural substrate treated with elevated Al concentrations. Tree Physiol. 2003, 23, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Han, Q.; Li, J. Soil propagule bank of ectomycorrhizal fungi associated with Masson pine (Pinus massoniana) grown in a manganese mine wasteland. PLoS ONE 2018, 13, e0198628. [Google Scholar] [CrossRef]
- Huang, J.; Nara, K.; Lian, C.; Zong, K.; Peng, K.; Xue, S.; Shen, Z. Ectomycorrhizal fungal communities associated with Masson pine (Pinus massoniana Lamb.) in Pb–Zn mine sites of central south China. Mycorrhiza 2012, 22, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Nara, K.; Zong, K.; Wang, J.; Xue, S.; Peng, K.; Shen, Z.; Lian, C. Ectomycorrhizal fungal communities associated with Masson pine (Pinus massoniana) and white oak (Quercus fabri) in a manganese mining region in Hunan Province, China. Fungal Ecol. 2014, 9, 1–10. [Google Scholar] [CrossRef]
- Thompson, G.W.; Medve, R.J. Effects of aluminum and manganese on the growth of ectomycorrhizal fungi. Appl. Environ. Microbiol. 1984, 48, 556–560. [Google Scholar] [CrossRef] [Green Version]
- Leski, T.; Rudawska, M.; Kieliszewska-Rokicka, B. Intraspecific aluminium response in Suillus luteus (L.) sf gray., an ectomycorrhizal symbiont of scots pine. Acta Soc. Bot. Pol. 1995, 64, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Johnson, C.E. Relationships among pH, aluminum solubility and aluminum complexation with organic matter in acid forest soils of the Northeastern United States. Geoderma 2016, 271, 234–242. [Google Scholar] [CrossRef]
- Li, H.; Huang, J.-G.; Yuan, L. Influence of aluminum and manganese on the growth, nutrient uptake and the efflux by ectomycorrhizal fungi. Environ. Sci. 2013, 34, 315–320. [Google Scholar]
- Wang, M.; Yuan, L.; Huang, J.; Zhou, Z. Al3+ Absorption and Assimilation by Four Ectomycorrhizal Fungi. Environ. Sci. 2015, 36, 3479–3485. [Google Scholar]
- Yang, X.; Xia, X.; Zhang, Z.; Nong, B.; Zeng, Y.; Wu, Y.; Xiong, F.; Zhang, Y.; Liang, H.; Pan, Y. Identification of anthocyanin biosynthesis genes in rice pericarp using PCAMP. Plant Biotechnol. J. 2019, 17, 1700. [Google Scholar] [CrossRef] [Green Version]
- Glatz, A.; Horváth, I.; Varvasovszki, V.; Kovács, E.; Török, Z.; Vigh, L. Chaperonin Genes of theSynechocystisPCC 6803 Are Differentially Regulated under Light–Dark Transition during Heat Stress. Biochem. Biophys. Res. Commun. 1997, 239, 291–297. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Sharma, A.; Rustad, T.; Mahajan, G.; Kumar, A.; Rao, K.V.; Banerjee, S.; Sherman, D.R.; Mande, S.C. Towards understanding the biological function of the unusual chaperonin Cpn60. 1 (GroEL1) of Mycobacterium tuberculosis. Tuberculosis 2016, 97, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Ortbauer, M.; Vahdati, K.; Leslie, C. Abiotic stress adaptation: Protein folding stability and dynamics. Abiotic Stress-Plant Responses Appl. Agric. 2013, 1, 3–25. [Google Scholar]
- Aremu, D.A.; Ezomo, O.F.; Meshitsuka, S. Gene expression in primary cultured astrocytes affected by aluminum: Alteration of chaperons involved in protein folding. Environ. Health Prev. Med. 2011, 16, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Ectopic expression of Pokkali phosphoglycerate kinase-2 (OsPGK2-P) improves yield in tobacco plants under salinity stress. Plant Cell Rep. 2016, 35, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Kapoor, A.; Zhu, J.-K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.C.; Qu, G.Z.; Li, H.Y.; Wu, Y.J.; Wang, C.; Liu, G.F.; Yang, C.P. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 2010, 37, 1119. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef]
- Ezaki, B.; Gardner, R.C.; Ezaki, Y.; Matsumoto, H. Expression of aluminum-induced genes in transgenic Arabidopsis plants can ameliorate aluminum stress and/or oxidative stress. Plant Physiol. 2000, 122, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Watt, D.A. Aluminium-responsive genes in sugarcane: Identification and analysis of expression under oxidative stress. J. Exp. Bot. 2003, 54, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Boscolo, P.R.; Menossi, M.; Jorge, R.A. Aluminum-induced oxidative stress in maize. Phytochemistry 2003, 62, 181–189. [Google Scholar] [CrossRef]
- Hossain, Z.; Nouri, M.-Z.; Komatsu, S. Plant cell organelle proteomics in response to abiotic stress. J. Proteome Res. 2012, 11, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Uchida, A.; Nakayama, K.; Okada, M. Water-soluble chlorophyll protein in Brassicaceae plants is a stress-induced chlorophyll-binding protein. Plant Cell Physiol. 2001, 42, 906–911. [Google Scholar] [CrossRef]
- La Roche, J.; Van der Staay, G.; Partensky, F.; Ducret, A.; Aebersold, R.; Li, R.; Golden, S.; Hiller, R.; Wrench, P.; Larkum, A. Independent evolution of the prochlorophyte and green plant chlorophyll a/b light-harvesting proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15244–15248. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-W.; Zhu, Y.-L.; Chen, C.-Y.; Geng, Z.-J.; Li, X.-Y.; Ye, T.-T.; Mao, X.-N.; Du, F. Cloning and characterization of two chlorophyll A/B binding protein genes and analysis of their gene family in Camellia sinensis. Sci. Rep. 2020, 10, 4602. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Zhou, Y.; Sun, P.; Cao, M.; Li, H.; Mao, Y. Identification of light-harvesting chlorophyll a/b-binding protein genes of Zostera marina L. and their expression under different environmental conditions. J. Ocean Univ. China 2016, 15, 152–162. [Google Scholar] [CrossRef]
- Park, Y.B.; Cosgrove, D.J. A revised architecture of primary cell walls based on biomechanical changes induced by substrate-specific endoglucanases. Plant Physiol. 2012, 158, 1933–1943. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.A.; Ahmad, J.; Bagheri, R.; Ali, A.A.; Al-Huqail, A.A.; Ibrahim, M.M.; Qureshi, M.I. Proteomic and ecophysiological responses of soybean (Glycine max L.) root nodules to Pb and hg stress. BMC Plant Biol. 2018, 18, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Houston, K.; Tucker, M.R.; Chowdhury, J.; Shirley, N.; Little, A. The plant cell wall: A complex and dynamic structure as revealed by the responses of genes under stress conditions. Front. Plant Sci. 2016, 7, 984. [Google Scholar] [CrossRef] [Green Version]
- Matern, U.; Grimmig, B.; Kneusel, R.E. Plant cell wall reinforcement in the disease-resistance response: Molecular composition and regulation. Can. J. Bot. 1995, 73, 511–517. [Google Scholar] [CrossRef]
- Chebli, Y.; Geitmann, A. Cellular growth in plants requires regulation of cell wall biochemistry. Curr. Opin. Cell Biol. 2017, 44, 28–35. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 4, 61. [Google Scholar] [CrossRef]
- Tucker, M.R.; Koltunow, A.M. Traffic monitors at the cell periphery: The role of cell walls during early female reproductive cell differentiation in plants. Curr. Opin. Plant Biol. 2014, 17, 137–145. [Google Scholar] [CrossRef]
- Sang, Q.; Shan, X.; An, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic analysis reveals the positive effect of exogenous spermidine in tomato seedlings’ response to high-temperature stress. Front. Plant Sci. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Liu, Y.; Wu, G.; Zhang, N.; Shen, Q.; Zhang, R. Beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 induces plant salt tolerance through spermidine production. Mol. Plant-Microbe Interact. 2017, 30, 423–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.; Shikanai, T. Supercomplex formation with photosystem I is required for the stabilization of the chloroplast NADH dehydrogenase-like complex in Arabidopsis. Plant Physiol. 2011, 155, 1629–1639. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Kuniyoshi, T.; Yamamoto, H.; Sugimoto, K.; Ishizaki, K.; Kohchi, T.; Nishimura, Y.; Shikanai, T. Composition and physiological function of the chloroplast NADH dehydrogenase-like complex in Marchantia polymorpha. Plant J. 2012, 72, 683–693. [Google Scholar] [CrossRef]
- Townsend, D.M.; Manevich, Y.; He, L.; Hutchens, S.; Pazoles, C.J.; Tew, K.D. Novel role for glutathione S-transferase π regulator of protein S-glutathionylation following oxidative and nitrosative stress. J. Biol. Chem. 2009, 284, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Roxas, V.P.; Smith, R.K.; Allen, E.R.; Allen, R.D. Overexpression of glutathione S-transferase/glutathioneperoxidase enhances the growth of transgenic tobacco seedlings during stress. Nat. Biotechnol. 1997, 15, 988–991. [Google Scholar] [CrossRef]
- Park, S.; Moon, J.-C.; Park, Y.C.; Kim, J.-H.; Kim, D.S.; Jang, C.S. Molecular dissection of the response of a rice leucine-rich repeat receptor-like kinase (LRR-RLK) gene to abiotic stresses. J. Plant Physiol. 2014, 171, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Timotijević, G.S.; Milisavljević, M.D.; Radović, S.R.; Konstantinović, M.M.; Maksimović, V.R. Ubiquitous aspartic proteinase as an actor in the stress response in buckwheat. J. Plant Physiol. 2010, 167, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liu, L.; Chen, C.; Chen, L. Regulations of granule-bound starch synthase I gene expression in rice leaves by temperature and drought stress. Biol. Plant. 2006, 50, 537–541. [Google Scholar] [CrossRef]
- Chen, H.-J.; Chen, J.-Y.; Wang, S.-J. Molecular regulation of starch accumulation in rice seedling leaves in response to salt stress. Acta Physiol. Plant. 2008, 30, 135–142. [Google Scholar] [CrossRef]
- Hurley, B.A.; Tran, H.T.; Marty, N.J.; Park, J.; Snedden, W.A.; Mullen, R.T.; Plaxton, W.C. The dual-targeted purple acid phosphatase isozyme AtPAP26 is essential for efficient acclimation of Arabidopsis to nutritional phosphate deprivation. Plant Physiol. 2010, 153, 1112–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, Z.; Qian, W.; Guo, W.; Gao, X.; Huang, L.; Wang, H.; Zhu, H.; Wu, J.-W.; Wang, D. The Arabidopsis purple acid phosphatase AtPAP10 is predominantly associated with the root surface and plays an important role in plant tolerance to phosphate limitation. Plant Physiol. 2011, 157, 1283–1299. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Peang, H.; Li, X.; Liu, C.; Lv, X.; Wei, X.; Zou, A.; Zhang, J.; Fan, G.; Ma, G. Genome-wide analysis of NDR1/HIN1-like genes in pepper (Capsicum annuum L.) and functional characterization of CaNHL4 under biotic and abiotic stresses. Hortic. Res. 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Grant, C.M.; Quinn, K.A.; Dawes, I.W. Differential protein S-thiolation of glyceraldehyde-3-phosphate dehydrogenase isoenzymes influences sensitivity to oxidative stress. Mol. Cell. Biol. 1999, 19, 2650–2656. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Kwon, H.-B.; Peng, H.-P.; Shih, M.-C. Stress responses and metabolic regulation of glyceraldehyde-3-phosphate dehydrogenase genes in Arabidopsis. Plant Physiol. 1993, 101, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Baudouin-Cornu, P.; Labarre, J. Regulation of the cadmium stress response through SCF-like ubiquitin ligases: Comparison between Saccharomyces cerevisiae, Schizosaccharomyces pombe and mammalian cells. Biochimie 2006, 88, 1673–1685. [Google Scholar] [CrossRef]
- Becerra, C.; Jahrmann, T.; Puigdomènech, P.; Vicient, C.M. Ankyrin repeat-containing proteins in Arabidopsis: Characterization of a novel and abundant group of genes coding ankyrin-transmembrane proteins. Gene 2004, 340, 111–121. [Google Scholar] [CrossRef]
- Pyo, Y.; Kim, G.M.; Choi, S.W.; Song, C.Y.; Yang, S.W.; Jung, I.L. Strontium stress disrupts miRNA biogenesis by reducing HYL1 protein levels in Arabidopsis. Ecotoxicol. Environ. Saf. 2020, 204, 111056. [Google Scholar] [CrossRef]
- Bertini, L.; Palazzi, L.; Proietti, S.; Pollastri, S.; Arrigoni, G.; Polverino de Laureto, P.; Caruso, C. Proteomic analysis of MeJa-induced defense responses in rice against wounding. Int. J. Mol. Sci. 2019, 20, 2525. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, L.; Su, H.; Guo, L.; Zhang, J.; Li, Y.; Xu, J.; Zhang, X.; Guo, Y.-D.; Zhang, N. Jasmonate and aluminum crosstalk in tomato: Identification and expression analysis of WRKYs and ALMTs during JA/Al-regulated root growth. Plant Physiol. Biochem. 2020, 154, 409–418. [Google Scholar] [CrossRef]
- Le Roy, J.; Blervacq, A.-S.; Créach, A.; Huss, B.; Hawkins, S.; Neutelings, G. Spatial regulation of monolignol biosynthesis and laccase genes control developmental and stress-related lignin in flax. BMC Plant Biol. 2017, 17, 1–20. [Google Scholar] [CrossRef]
- Hu, Q.; Min, L.; Yang, X.; Jin, S.; Zhang, L.; Li, Y.; Ma, Y.; Qi, X.; Li, D.; Liu, H. Laccase GhLac1 modulates broad-spectrum biotic stress tolerance via manipulating phenylpropanoid pathway and jasmonic acid synthesis. Plant Physiol. 2018, 176, 1808–1823. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Haroldsen, V.; Cai, X.; Wu, Y. Expression of a putative laccase gene, ZmLAC1, in maize primary roots under stress. Plant Cell Environ. 2006, 29, 746–753. [Google Scholar] [CrossRef]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef]
- Li, J.; Hu, L.; Zhang, L.; Pan, X.; Hu, X. Exogenous spermidine is enhancing tomato tolerance to salinity–alkalinity stress by regulating chloroplast antioxidant system and chlorophyll metabolism. BMC Plant Biol. 2015, 15, 1–17. [Google Scholar] [CrossRef]
- Kasukabe, Y.; He, L.; Nada, K.; Misawa, S.; Ihara, I.; Tachibana, S. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.-P.; Pang, X.-M.; Matsuda, N.; Kita, M.; Inoue, H.; Hao, Y.-J.; Honda, C.; Moriguchi, T. Over-expression of the apple spermidine synthase gene in pear confers multiple abiotic stress tolerance by altering polyamine titers. Transgenic Res. 2008, 17, 251–263. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhao, T.; Kermode, A.R. A conifer ABI3-interacting protein plays important roles during key transitions of the plant life cycle. Plant Physiol. 2013, 161, 179–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Garreton, V.; Chua, N.-H. The AIP2 E3 ligase acts as a novel negative regulator of ABA signaling by promoting ABI3 degradation. Genes Dev. 2005, 19, 1532–1543. [Google Scholar] [CrossRef] [Green Version]
- Carrie, C.; Small, I. A reevaluation of dual-targeting of proteins to mitochondria and chloroplasts. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Lecoy, J.; García-Gil, M. Evolutionary analysis of LP3 gene family in conifers: An ASR homolog. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Lee, S.Y.; Kim, W.Y.; Jung, Y.J.; Chae, H.B.; Jung, H.S.; Kang, C.H.; Shin, M.R.; Kim, S.Y.; Su’udi, M. Heat-induced chaperone activity of serine/threonine protein phosphatase 5 enhances thermotolerance in Arabidopsis thaliana. New Phytol. 2011, 191, 692–705. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer Sequence | Reverse Primer Sequence | E Value (%) | R2 |
|---|---|---|---|---|
| TRINITY_DN51949_c0_g2 | CCGTCATCGCTCCAGT | CACAGTTCGCCCTTCA | 93.2 | 0.94 |
| TRINITY_DN44650_c0_g1 | GGAAAGTGGGTGGTCT | AGGAGTTCGTGGGATT | 91.5 | 0.86 |
| TRINITY_DN50464_c0_g3 | ATTGATAGGAGGCTGA | TGAGGGAACTACGAGA | 89.6 | 0.95 |
| TRINITY_DN39799_c0_g2 | GAGACAATGTGGTGGC | TTTGGCAGTGTAAGCA | 94 | 0.91 |
| TRINITY_DN39005_c0_g1 | GCTACACCCTCGCAGT | AGCACGACCAGGAAAC | 87.6 | 0.88 |
| TRINITY_DN43045_c0_g9 | CCTTGAACCCAAATACA | ACGGGCTTACCAGTCT | 91.3 | 0.92 |
| TRINITY_DN43345_c2_g1 | AACAAGCCGTTGGACT | GGGAACAAAGGATGGG | 92.4 | 0.95 |
| TRINITY_DN47656_c0_g2 | CCTGTATTGCCTGATG | GACGAGATGGTGGAGT | 88.5 | 0.93 |
| TRINITY_DN47910_c0_g1 | TCACCTGCCATACAAA | TCCAGCATCAAAGAAA | 91 | 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Chen, H.; Ding, G.; Li, K.; Wang, Y. Proteomic Insight into the Symbiotic Relationship of Pinus massoniana Lamb and Suillus luteus towards Developing Al-Stress Resistance. Life 2021, 11, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020177
Liu H, Chen H, Ding G, Li K, Wang Y. Proteomic Insight into the Symbiotic Relationship of Pinus massoniana Lamb and Suillus luteus towards Developing Al-Stress Resistance. Life. 2021; 11(2):177. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020177
Chicago/Turabian StyleLiu, Haiyan, Houying Chen, Guijie Ding, Kuaifen Li, and Yao Wang. 2021. "Proteomic Insight into the Symbiotic Relationship of Pinus massoniana Lamb and Suillus luteus towards Developing Al-Stress Resistance" Life 11, no. 2: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020177