
Transcriptome-Wide Analysis for Ginsenoside Rb3 Synthesis-Related Genes and Study on the Expression of Methyl Jasmonate Treatment in Panax ginseng

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Basic Analysis of Ginsenoside Rb3 Synthesis-Related Genes
2.2. Screening of Candidate Genes Related to Ginsenoside Rb3 Synthesis
2.3. GO (Gene Ontology) Annotation for Candidate Genes Related to Ginsenoside Rb3 Synthesis
2.4. Enrichment Analysis of Candidate Genes Related to Ginsenoside Rb3 Synthesis
2.5. Analysis of Spatiotemporal Expression Patterns of PgRb3 Genes
2.6. Correlation Analysis of PgRb3 Genes and Saponin Content
2.7. Correlation Analysis of PgRb3 Genes and Known Key Enzyme Genes for Ginsenoside Synthesis
2.8. Expressions of PgRb3 Genes in Ginseng Hairy Roots by MeJA Treatment
3. Results
3.1. Basic Analysis of Candidate Gene Population Related to Ginsenoside Rb3 Synthesis
3.2. Screening of Candidate Genes Related to Ginsenoside Rb3 Synthesis
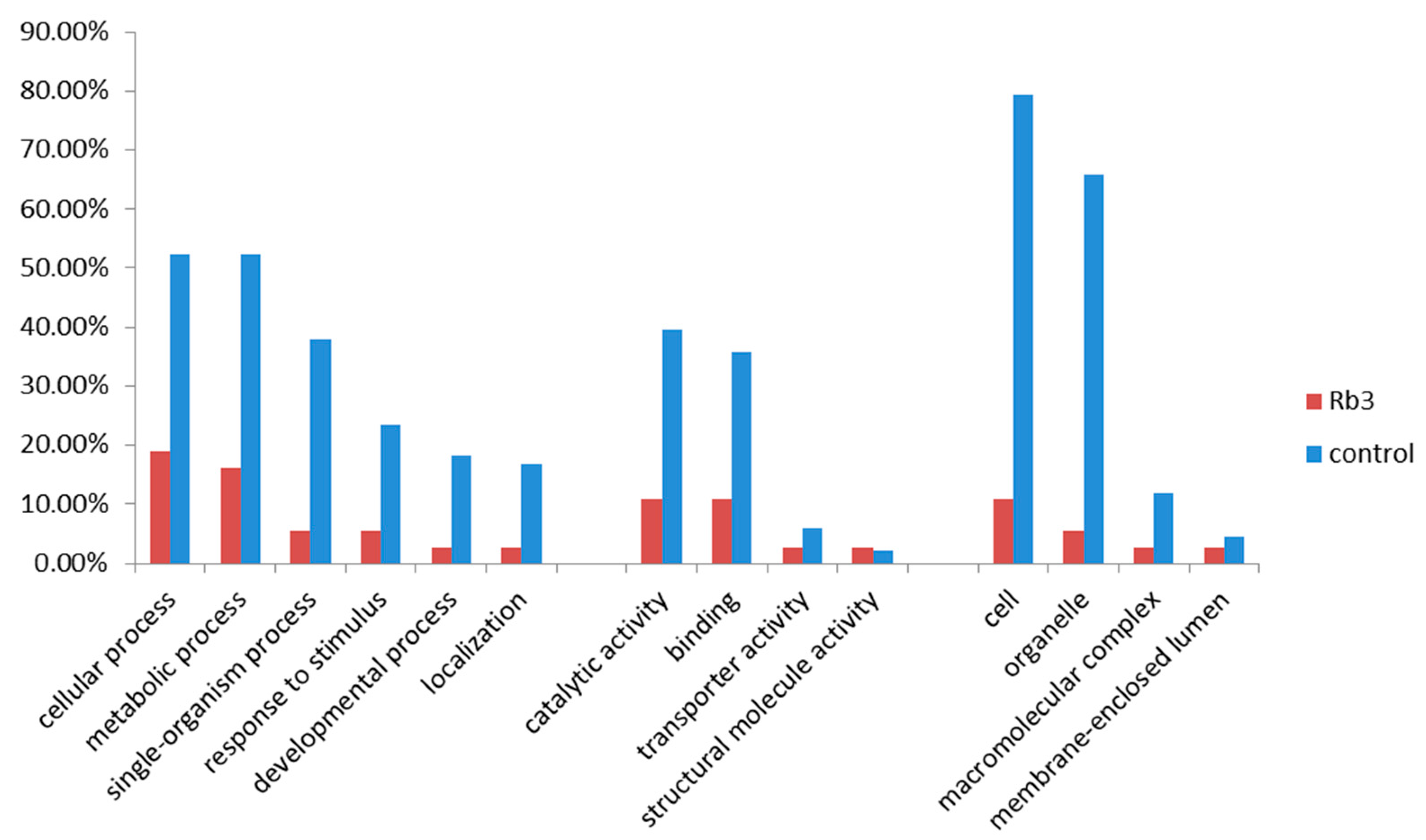
3.3. Annotation of GO Function for Candidate Genes Related to Ginsenoside Rb3 Synthesis
3.4. Enrichment Analysis of GO Annotation for PgRb3 Genes
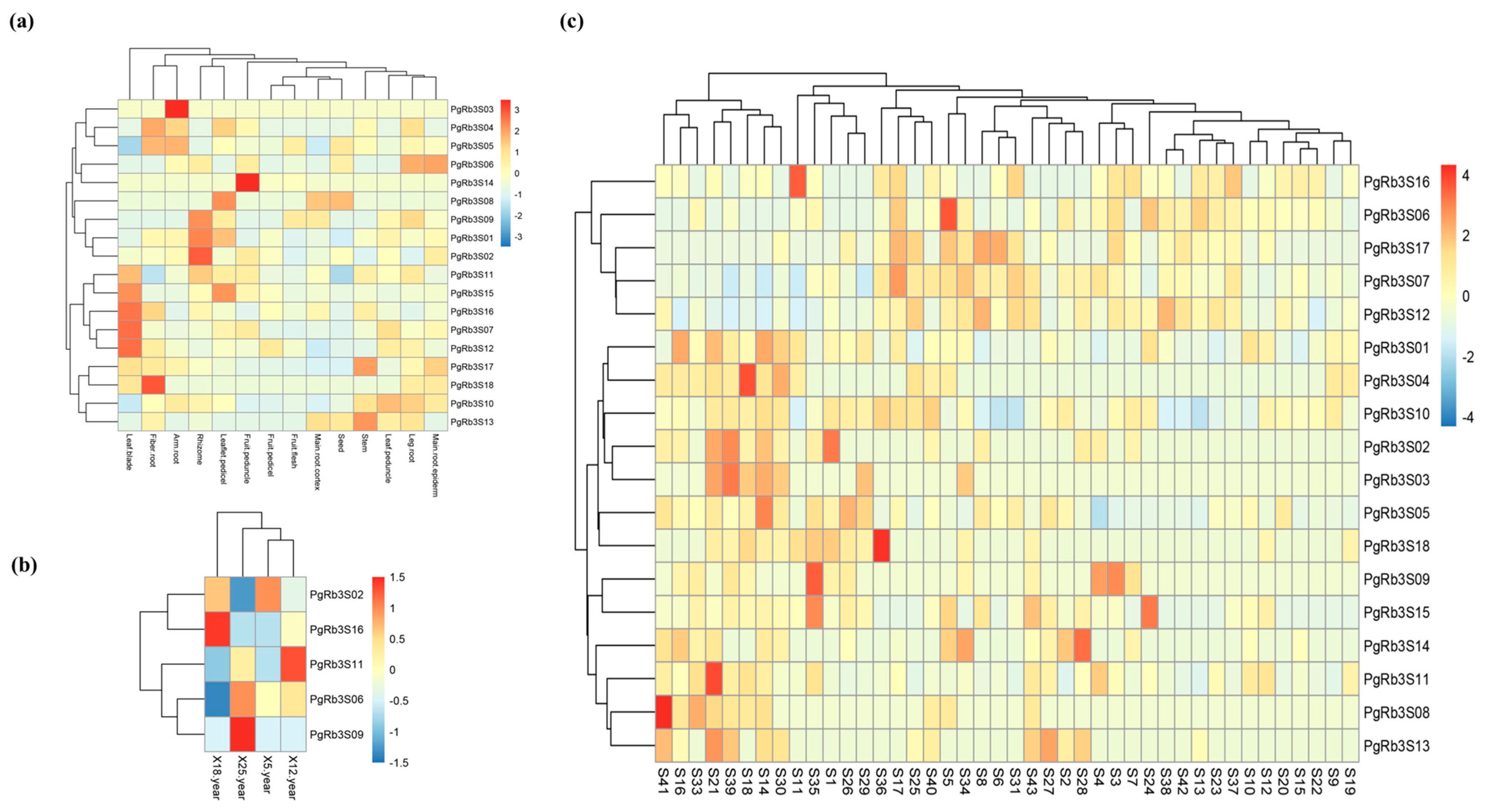
3.5. Heatmap Analysis for Spatiotemporal Expression Patterns of PgRb3 Genes
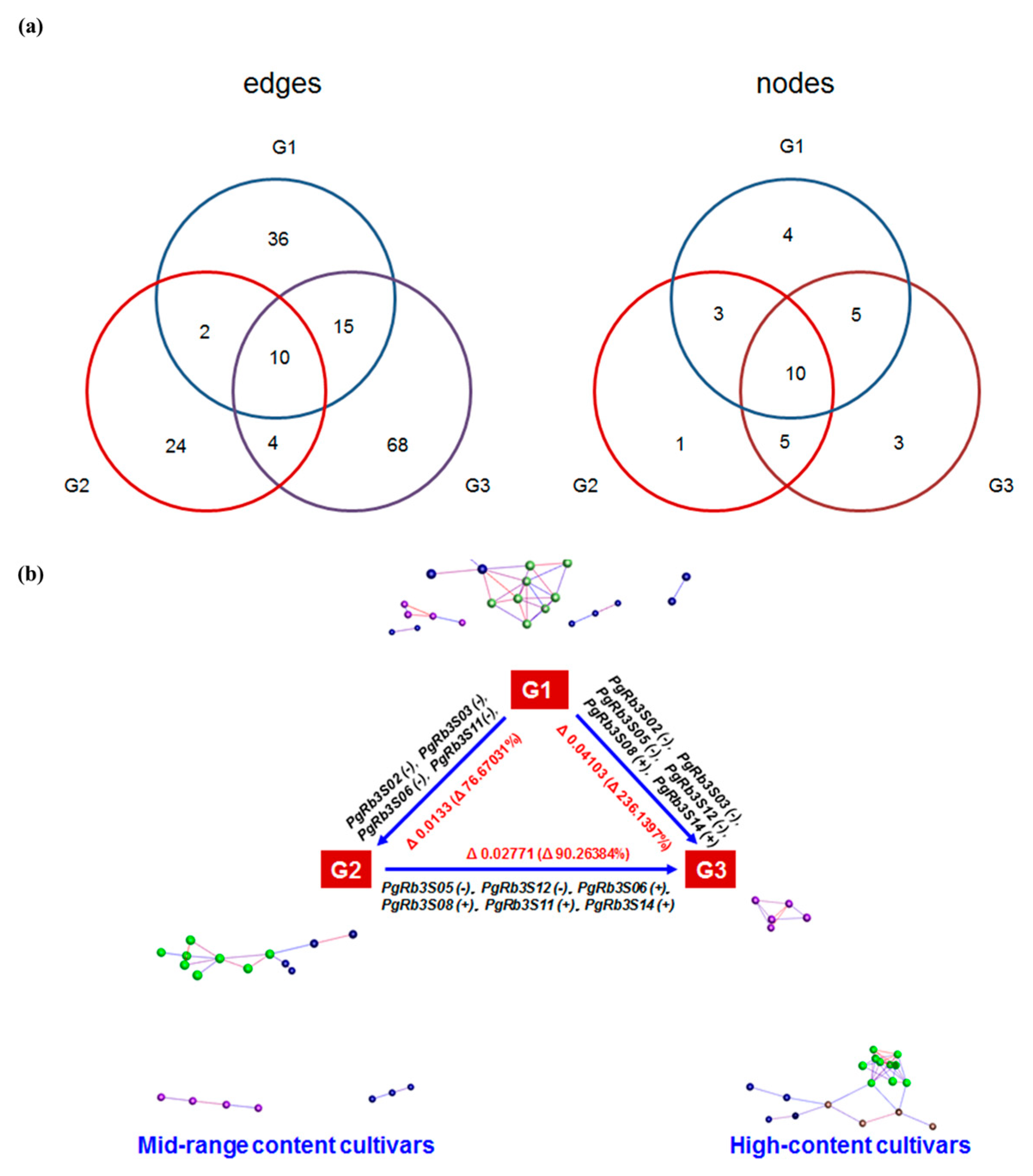
3.6. Network Analysis for Spatiotemporal Expression Patterns of PgRb3 Genes
3.7. Correlation Analysis of PgRb3 Genes Expression and Ginsenoside Content
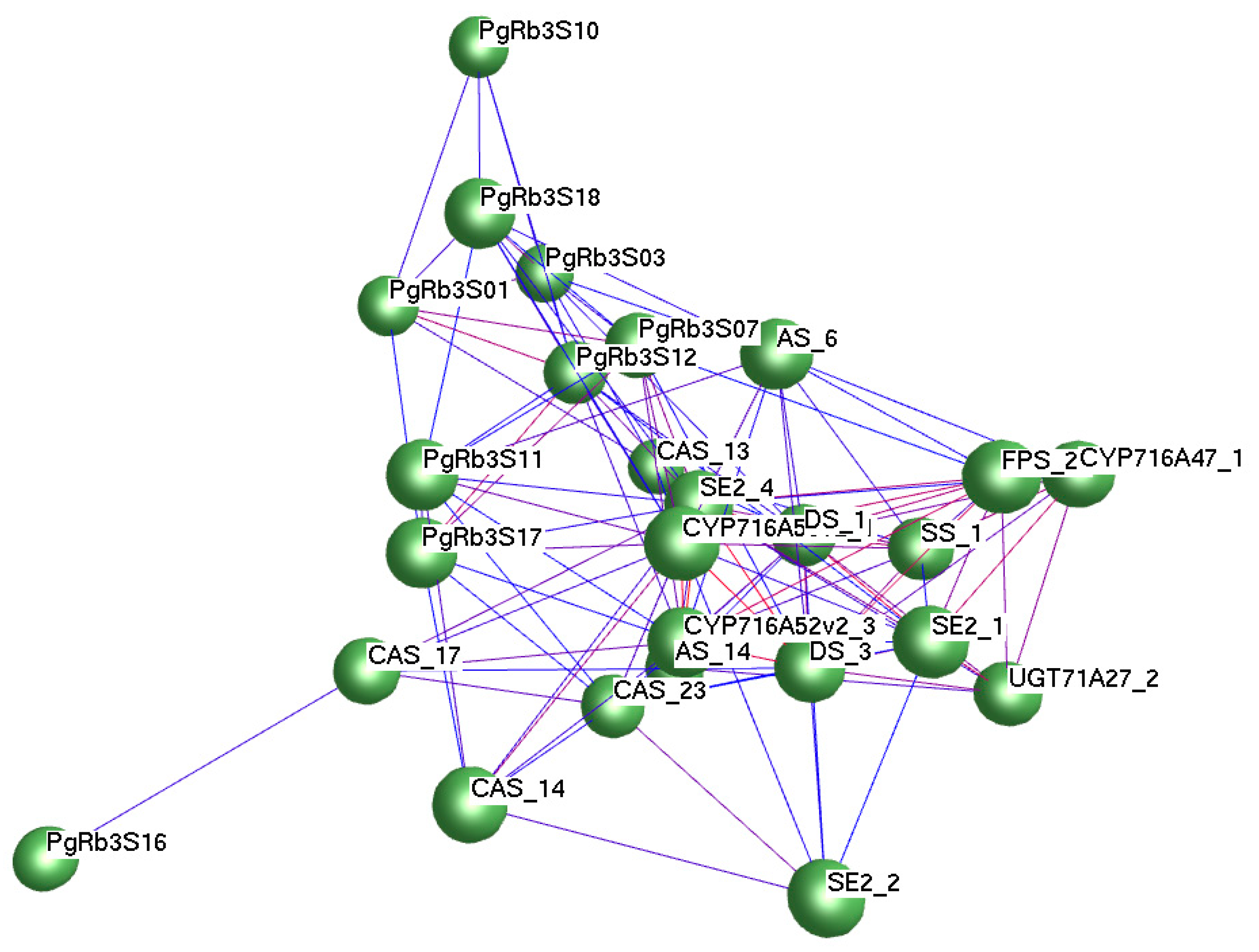
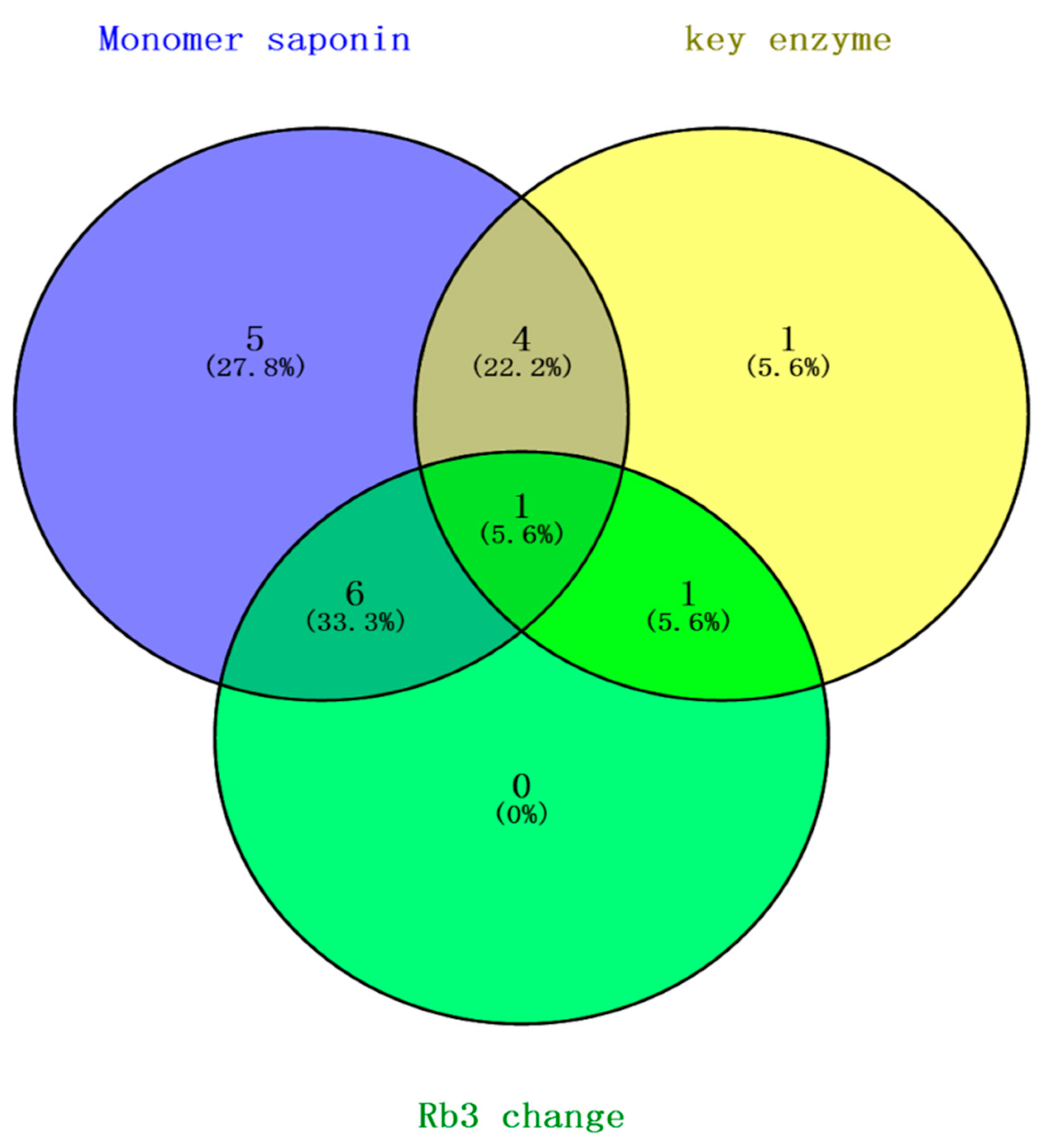
3.8. Correlation Analysis of PgRb3 Genes and Known Ginsenoside Synthesis Key Enzyme Genes
3.9. Identification of PgRb3 Genes Related to Ginsenoside Rb3 Synthesis in Panax ginseng
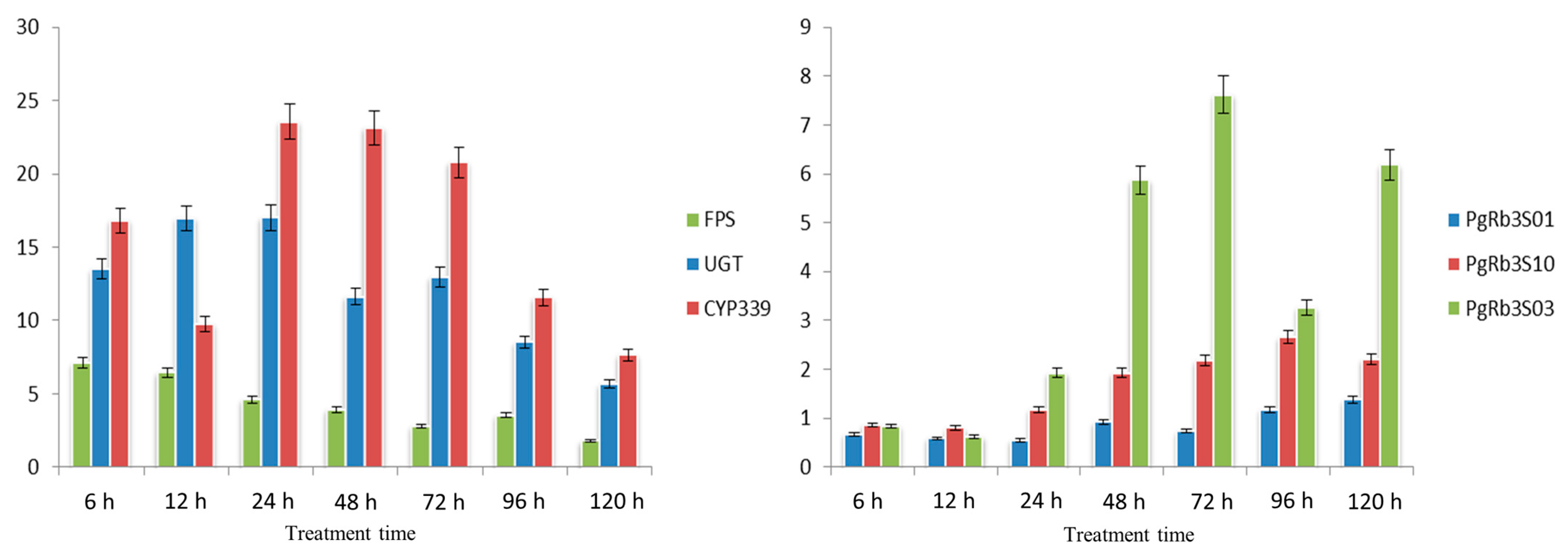
3.10. Study on PgRb3 Gene Expression Regularity under MeJA Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hemmerly, T.E. A ginseng farm in Lawrence County, Tennessee. Econ. Bot. 1997, 31, 160–162. [Google Scholar] [CrossRef]
- Leung, K.; Wong, A. Pharmacology of ginsenosides: A literature review. Chin. Med. 2010, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Paek, K.Y.; Murthy, H.N.; Hahn, E.J.; Zhong, J.J. Large scale culture of ginseng adventitious roots for production of ginsenosides. Adv. Biochem. Engin. Biotechnol. 2009, 113, 151–176. [Google Scholar]
- Zhang, Y.C.; Geng, L.; Chao, J.; Yang, B.; Yang, H.J.; Xu, H.Y.; Huang, L.Q. Tissue-specific distribution of ginsenosides in different aged ginseng and antioxidant activity of ginseng leaf. Molecules 2014, 19, 17381–17399. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.I.; Park, K.S.; Cho, I.H. Panax ginseng: A candidate herbal medicine for autoimmune disease. J. Ginseng Res. 2019, 43, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Zhang, D.B.; Yang, D.C. Biosynthesis and biotechnological production of ginsenosides. Biotechnol. Adv. 2015, 33, 717–735. [Google Scholar] [CrossRef]
- Eom, S.J.; Kim, K.T.; Paik, H.D. Microbial bioconversion of ginsenosides in Panax ginseng and their improved bioactivities. Food Rev. Int. 2018, 34, 1–15. [Google Scholar] [CrossRef]
- Kim, D.S.; Chang, Y.J.; Zedk, U.; Zhao, P.; Liu, Y.Q.; Yang, C.R. Dammarane saponins from Panax ginseng. Phytochemistry 1995, 40, 1493–1497. [Google Scholar] [PubMed]
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99. [Google Scholar]
- Tung, N.H.; Song, G.Y.; Park, Y.J.; Kim, Y.H. Two new dammaranetype saponins from the leaves of Panax ginseng. Chem. Pharm. Bull. 2009, 57, 1412–1414. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.Y.; Jiang, S.C.; Sun, C.Y.; Lin, Y.P.; Yin, R.; Wang, Y.; Zhang, M.P. The spatial and temporal transcriptomic landscapes of ginseng, Panax ginseng C. A. Meyer. Sci. Rep. 2015, 5, 18283. [Google Scholar] [CrossRef]
- Xu, J.; Chu, Y.; Liao, B.S.; Xiao, S.M.; Yin, Q.G.; Bai, R.; Su, H.; Dong, L.L.; Li, X.W.; Qian, J.; et al. Panax ginseng genome examination for ginsenoside biosynthesis. Gigasci. 2017, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.H.; Jayakodi, M.; Lee, S.C.; Choi, B.S.; Jang, W.; Lee, J.; Kim, H.H.; Waminal, N.E.; Lakshmanan, M.; Nguyen, B.V.; et al. Genome and evolution of the shade-requiring medicinal herb Panax ginseng. Plant Biotechnol. J. 2018, 16, 1904–1917. [Google Scholar] [CrossRef] [Green Version]
- Azimova, S.S. (Ed.) Ginsenoside-Rb3. In Natural Compounds; Springer: New York, NY, USA, 2013; p. 881. [Google Scholar]
- Xiang, Y.Z.; Shang, H.C.; Gao, X.M.; Zhang, B.L. A comparison of the ancient use of ginseng in traditional Chinese medicine with modern pharmacological experiments and clinical trials. Phytother. Res. 2008, 22, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Min, H. Ginseng, the immunity boost: The effects of Panax ginseng on immune system. J. Ginseng Res. 2012, 36, 354–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.S.; Bae, K.G.; Jung, I.S.; Kim, C.H.; Yun, Y.S.; Song, J.Y. Anti-septicaemic effect of polysaccharide from Panax ginseng by macrophage activation. J. Infect. 2002, 45, 32–38. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, D.; Lee, H.L.; Kim, C.E.; Jung, K.; Kang, K.S. Beneficial effects of Panax ginseng for the treatment and prevention of neurodegenerative diseases: Past findings and future directions. J. Ginseng Res. 2018, 42, 239–247. [Google Scholar] [CrossRef]
- Joo, S.S.; Lee, D.I. Potential effects of microglial activation induced by ginsenoside Rg3 in rat primary culture: Enhancement of type A Macrophage Scavenger Receptor expression. Arch. Pharm. Res. 2005, 28, 1164–1169. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Yu, K.W.; Gao, W.Y.; Son, S.H.; Paek, K.Y. Improvement of ginsenoside production by jasmonic acid and some other elicitors in hairy root culture of ginseng (Panax ginseng C.A. Meyer). Vitr. Cell. Dev. Biol. Plant 2000, 36, 424–428. [Google Scholar] [CrossRef]
- Palazón, J.; Cusido, R.M.; Bonfill, M.; Mallol, A.; Moyano, E.; Morales, C.; Piñol, M.T. Elicitation of different Panax ginseng transformed root phenotypes for an improved ginsenoside production. Plant Physiol. Biochem. 2003, 41, 1019–1025. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Javier, T.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Theocharidis, A.; Van Dongen, S.; Enright, A.J.; Freeman, T.C. Network visualization and analysis of gene expression data using BioLayout Express (3D). Nat. Protoc. 2009, 4, 1535–1550. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Jeong, J.H.; Seo, J.W.; Shin, C.G.; Kim, Y.S.; In, J.G.; Yang, D.C.; Yi, J.S.; Choi, Y.E. Enhanced triterpene and phytosterol biosynthesis in Panax ginseng overexpressing squalene synthase gene. Plant Cell Physiol. 2004, 45, 976–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.Y.; In, J.G.; Kwon, Y.S.; Choi, Y.E. Regulation of ginsenoside and phytosterol biosynthesis by RNA interferences of squalene epoxidase gene in Panax ginseng. Phytochemistry 2010, 71, 36–46. [Google Scholar] [CrossRef]
- Han, J.Y.; Kwon, Y.S.; Yang, D.C.; Jung, Y.R.; Choi, Y.E. Expression and RNA interference-induced silencing of the dammarenediol synthase gene in Panax ginseng. Plant Cell Physiol. 2006, 47, 1653–1662. [Google Scholar] [CrossRef]
- Kushiro, T.; Shibuya, M.; Ebizuka, Y. β-Amyrin synthase-Cloning of oxidosqualene cyclase that catalyzes the formation of the most popular triterpene among higher plants. Eur. J. Biochem. 1998, 256, 238–244. [Google Scholar] [CrossRef]
- Kim, O.T.; Bang, K.H.; Jung, S.J.; Kim, Y.C.; Hyun, D.Y.; Kim, S.H.; Cha, S.W. Molecular characterization of ginseng farnesyl diphosphate synthase gene and its up-regulation by methyl jasmonate. Biol. Plant. 2010, 54, 47–53. [Google Scholar] [CrossRef]
- Kim, O.T.; Bang, K.H.; Kim, Y.C.; Hyun, D.Y.; Kim, M.Y.; Cha, S.W. Upregulation of ginsenoside and gene expression related to triterpene biosynthesis in ginseng hairy root cultures elicited by methyl jasmonate. Plant Cell Tiss. Organ Cult. 2009, 98, 25–33. [Google Scholar] [CrossRef]
- Jung, S.C.; Kim, W.; Park, S.C.; Jeong, J.; Park, M.K.; Lim, S.; Lee, Y.; Im, W.T.; Lee, J.H.; Choi, G.; et al. Two ginseng UDP-glycosyltransferases synthesize ginsenoside Rg3 and Rd. Plant Cell Physiol. 2014, 55, 2177–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.Y.; Kim, H.J.; Kwon, Y.S.; Choi, Y.E. The Cyt P450 enzyme CYP716A47 catalyzes the formation of protopanaxadiol from dammarenediol-II during ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol. 2011, 52, 2062–2073. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Hwang, H.S.; Choi, S.W.; Kim, H.J.; Choi, Y.E. Cytochrome P450 CYP716A53v2 catalyzes the formation of protopanaxatriol from protopanaxadiol during ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol. 2012, 53, 1535–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.Y.; Kim, M.J.; Ban, Y.W.; Hwang, H.S.; Choi, Y.E. The involvement of β-amyrin 28-oxidase (CYP716A52v2) in oleanane-type ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol. 2013, 54, 2034–2046. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, K.Y.; Zhao, M.Z.; Li, S.K.; Jiang, Y.; Zhu, L.; Chen, J.; Wang, Y.F.; Sun, C.Y.; Chen, P.; et al. Selection and validation of reference genes desirable for gene expression analysis by qRT-PCR in MeJA-treated ginseng hairy roots. PLoS ONE 2019, 14, e0226168. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Zhao, M.Z.; Lin, Y.P.; Wang, Y.F.; Li, X.Y.; Han, Y.L.; Wang, K.Y.; Sun, C.Y.; Wang, Y.; Zhang, M.P. Transcriptome analysis identifies strong candidate genes for ginsenoside biosynthesis and reveals its underlying molecular mechanism in Panax ginseng C.A. Meyer. Sci. Rep. 2019, 9, 615. [Google Scholar] [CrossRef]
- Zhang, M.P.; Liu, Y.-H.; Xu, W.W.; Smith, C.W.; Murray, S.C.; Zhang, H.-B. Analysis of the genes controlling three quantitative traits in three diverse plant species reveals the molecular basis of quantitative traits. Sci. Rep. 2020, 10, 10074. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5’-3’) | Reverse Primer (5’-3’) |
|---|---|---|
| PgRb3S01 | CAATGAAAACCCTGAAACTCG | TAGGAGTATGAGGCAGAGGTAAAG |
| PgRb3S03 | TGGTCGGAGATCCACTTGCT | TTATTCATGCTGCTCATCTGATCTT |
| PgRb3S10 | GAAGATGGTCAAGGGACTGCT | TGTCGTCACCCACTTGCCT |
| UGT (UGT71A27_2) | TGCGTCCGTCTATCCCTAAAG | TGATGTCCTGTCCAAGAATCCTAC |
| CYP339 (CYP716A47_1) | GCAGAGGTTTACTTTGGCAC | TCACATTGATGGAGAGGACAC |
| FPS | GGATGATTATCTGGATTGCTTTGGCGAG | CAGTGCTTTTACTACCAACCAGGAG |
| GADPH | TTCCCACTGTGGATGTC | CTCCGACTCCTCCTTGATAGC |
| Number | Gene ID | Named ID |
|---|---|---|
| 1 | comp122476_c0_seq1 | PgRb3S01 |
| 2 | comp34816_c0_seq1 | PgRb3S02 |
| 3 | comp43937_c0_seq2 | PgRb3S03 |
| 4 | comp439496_c0_seq1 | PgRb3S04 |
| 5 | comp47140_c0_seq1 | PgRb3S05 |
| 6 | comp48914_c0_seq3 | PgRb3S06 |
| 7 | comp50009_c0_seq2 | PgRb3S07 |
| 8 | comp52220_c0_seq2 | PgRb3S08 |
| 9 | comp61172_c1_seq1 | PgRb3S09 |
| 10 | comp61669_c1_seq3 | PgRb3S10 |
| 11 | comp63877_c0_seq2 | PgRb3S11 |
| 12 | comp64020_c0_seq3 | PgRb3S12 |
| 13 | comp64616_c1_seq6 | PgRb3S13 |
| 14 | comp65415_c0_seq45 | PgRb3S14 |
| 15 | comp65868_c1_seq1 | PgRb3S15 |
| 16 | comp65985_c1_seq26 | PgRb3S16 |
| 17 | comp66742_c1_seq1 | PgRb3S17 |
| 18 | comp709833_c0_seq1 | PgRb3S18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Zhang, Z.; Li, S.; Hu, J.; Liu, T.; Jiang, Y.; Wu, J.; Lu, M.; Zhao, M.; Li, L.; et al. Transcriptome-Wide Analysis for Ginsenoside Rb3 Synthesis-Related Genes and Study on the Expression of Methyl Jasmonate Treatment in Panax ginseng. Life 2021, 11, 387. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050387
Wang K, Zhang Z, Li S, Hu J, Liu T, Jiang Y, Wu J, Lu M, Zhao M, Li L, et al. Transcriptome-Wide Analysis for Ginsenoside Rb3 Synthesis-Related Genes and Study on the Expression of Methyl Jasmonate Treatment in Panax ginseng. Life. 2021; 11(5):387. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050387
Chicago/Turabian StyleWang, Kangyu, Zixuan Zhang, Shaokun Li, Jian Hu, Tao Liu, Yang Jiang, Jun Wu, Minghai Lu, Mingzhu Zhao, Li Li, and et al. 2021. "Transcriptome-Wide Analysis for Ginsenoside Rb3 Synthesis-Related Genes and Study on the Expression of Methyl Jasmonate Treatment in Panax ginseng" Life 11, no. 5: 387. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050387