
Are Juglans neotropica Plantations Useful as a Refuge of Bryophytes Diversity in Tropical Areas?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Design and Data Collection
2.3. Data Analysis
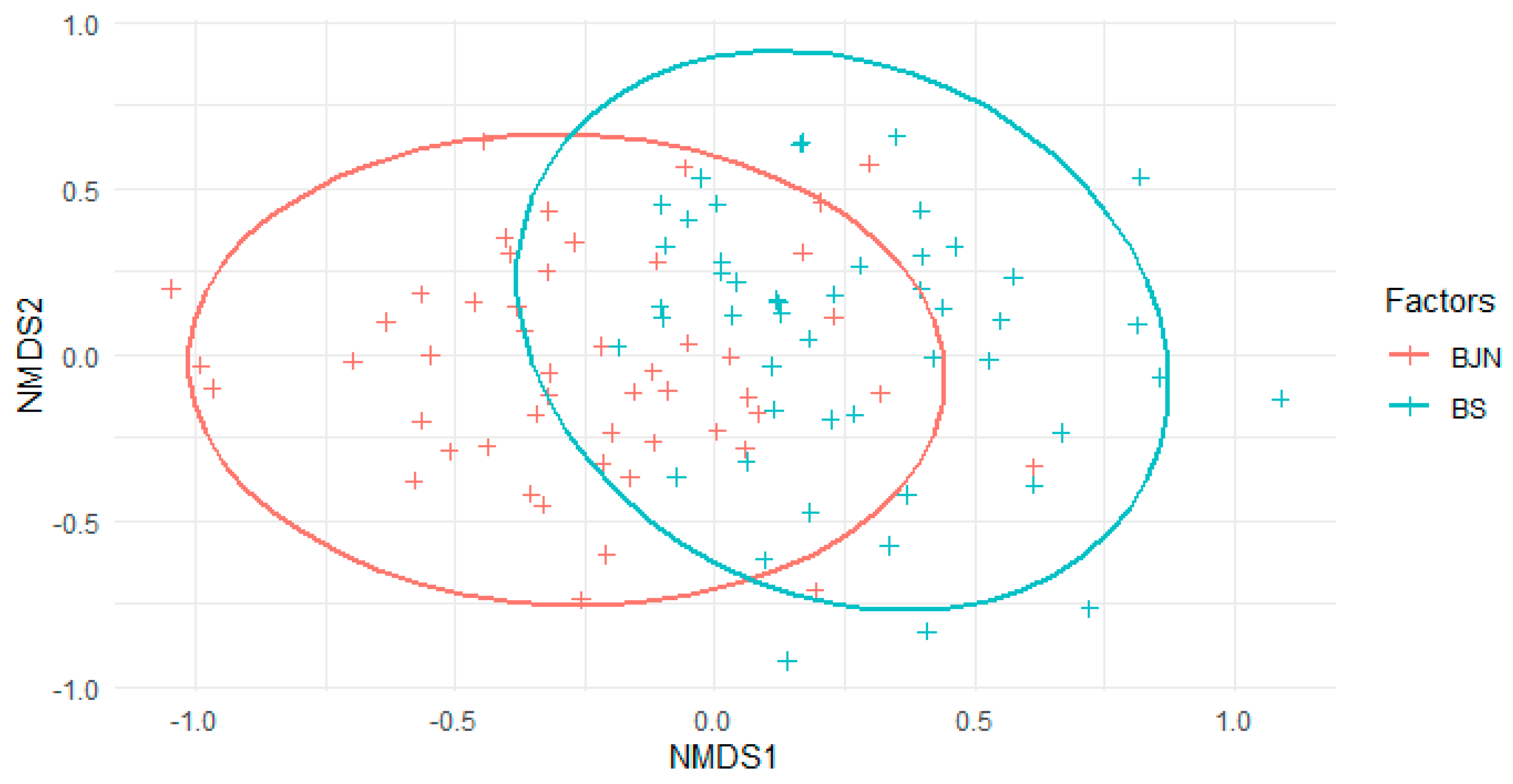
3. Results
3.1. Richness
3.2. Species Composition
3.3. Indicator Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Taxa | Forest of Juglans neotropica (BJN) | Secondary Forests (BS) |
|---|---|---|
| Liverworts | ||
| Aneuraceae | ||
| Riccardia digitiloba (Spruce) Pagan | 1 | |
| Adelanthaceae | ||
| Pseudomarsupidium decipiens (Hook.) Grolle | 1 | 8 |
| Frullanieaceae | ||
| Frullania brasiliensis Raddi | 6 | 1 |
| Frullania caulisequa (Nees) Mont. | 2 | 1 |
| Frullania ericoides (Nees) Mont. | 16 | |
| Frullania riojaneirensis (Raddi) Ångstr. | 15 | |
| Frullania subtilissima (Nees ex Mont.) Lindenb. | 1 | |
| Lejeuneaceae | ||
| Bryopteris filicina (Sw.) Nees | 1 | |
| Cheilolejeunea filiformis (Sw.) W. Ye, R.L. Zhu & Gradst. | 1 | |
| Dicranolejeunea axillaris (Nees & Mont.) Schiffn. | 1 | |
| Drepanolejeunea cutervoensis (Loitl.) Grolle | 1 | |
| Frullanoides densifolia Raddi subsp. densifolia. | 1 | |
| Lejeunea cerina (Lehm. & Lindenb.) Lehm. & Lindenb. | 9 | 15 |
| Lejeunea deplanata Nees | 9 | |
| Lejeunea flava (Sw.) Nees | 1 | 1 |
| Lejeunea laetevirens Nees & Mont. | 17 | 12 |
| Lejeunea ramulosa Spruce | 3 | |
| Marchesinia brachiata (Sw.) Schiffner | 1 | |
| Microlejeunea acutifolia Steph. | 1 | |
| Microlejeunea bullata (Taylor) Steph. | 1 | |
| Lophocoleaceae | ||
| * Leptoscyphus autoicus (J.J. Engel & Gradst.) Vanderp. and Gradst. | 1 | |
| Lophocolea bidentata (L.) Dumort. | 1 | 7 |
| Lophocolea muricata (Lehm.) Nees | 4 | |
| Metzgeriaceae | ||
| Metzgeria dorsipara (Herzog) Kuwah. | 2 | 1 |
| Metzgeria leptoneura Spruce | 1 | 3 |
| Plagiochilaceae | ||
| Plagiochila aerea Taylor | 1 | |
| Plagiochila bifaria (Sw.) Lindenb. | 1 | |
| Plagiochila cristata (Sw.) Lindenb. | 9 | 4 |
| Plagiochila diversifolia Lindenb. & Gottsche | 6 | 3 |
| Plagiochila gymnocalycina (Lehm. & Lindenb.) Mont. and Nees | 2 | 1 |
| Plagiochila raddiana Lindenb. | 29 | 36 |
| Porellaceae | ||
| Porella brachiata (Taylor) Spruce | 2 | 2 |
| Porella crispata (Hook.) Trevis. | 3 | 3 |
| Radulaceae | ||
| Radula episcia Spruce | 3 | 7 |
| Radula gottscheana Taylor | 7 | 8 |
| Radula tectiloba Steph. | 34 | 7 |
| Mosses | ||
| Bryaceae | ||
| Bryum apiculatum Schwägr | 1 | |
| Calymperaceae | ||
| Syrrhopodon incompletus Schwägr. | 7 | 5 |
| Cryphaeaceae | ||
| Cryphaea jamesonii Taylor | 3 | |
| Daltoniaceae | ||
| Adelothecium bogotense (Hampe) Mitt. | 1 | |
| Dicranaceae | ||
| Campylopus flexuosus (Hedw.) Brid. | 1 | |
| Fabroniaceae | ||
| Fabronia ciliaris (Brid.) Brid. | 1 | |
| Meteoriaceae | ||
| Meteoridium remotifolium (Müll. Hal.) Manuel | 6 | 5 |
| Squamidium leucotrichum (Taylor) Broth. | 3 | 5 |
| Squamidium nigricans (Hook.) Broth. | 2 | 3 |
| Zelometeorium recurvifolium (Hornsch.) Manuel | 13 | |
| Mniaceae | ||
| Plagiomnium rhynchophorum (Hook.) T.J. Kop. | 1 | |
| Neckeraceae | ||
| Neckeropsis undulata (Hedw.) Reichardt | 12 | 11 |
| Porotrichum filiferum Mitt. | 4 | |
| Porotrichum longirostre (Hook.) Mitt. | 15 | 26 |
| Orthotrichaceae | ||
| Macromitrium richardii Schwägr. | 2 | |
| Sematophyllaceae | ||
| Acroporium pungens (Hedw.) Broth. | 1 | 2 |
| Sematophyllum subsimplex (Hedw.) Mitt. | 14 | 7 |
| Thuidiaceae | ||
| Thuidium peruvianum Mitt. | 35 | 25 |
| Thuidium tomentosum Schimp. | 2 | 2 |
References
- Kapos, V.; Rhind, J.; Edwards, M.; Price, M.F.; Ravilious, C. Developing a map of the world’s mountain forests. In Forests in Sustainable Mountain Development: A State-of Knowledge Report for 2000; Price, M.F., Butt, N., Eds.; CAB International: Wallingford, UK, 2000; pp. 4–12. [Google Scholar]
- Antonelli, A.; Sanmartín, I. Why are there so many plant species in the Neotropics? Taxon 2011, 60, 403–414. [Google Scholar] [CrossRef]
- Lagomarsino, L.P.; Condamine, F.L.; Antonelli, A.; Mulch, A.; Davis, C.C. The abiotic and biotic drivers of rapid diversification in Andean bellflowers (Campanulaceae). New Phytol. 2016, 210, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.D.; Kappelle, M. (Eds.) Introducción a los bosques nublados del neotrópico: Una síntesis regional. In Bosques Nublados del Neotrópico; INBio: San Jose, Costa Rica, 2001; pp. 25–40. [Google Scholar]
- Churchill, S.P.; Balslev, H.; Forero, E.; Luteyn, J.L. Biodiversity and conservation of neotropical montane forests. In Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium, Bronx, NY, USA, 21–26 June 1993. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, F.; Peralvo, M.; Valarezo, N. Los Bosques Montanos de los Andes Tropicales: Una Evaluación Regional de su Estado de Conservación y de su Vulnerabilidad a Efecto del Cambio Climático; Serie Investigación y Sistematización; Imprenta Mariscal: Quito, Ecuador, 2009. [Google Scholar]
- Tejedor, G.N.; Álvarez, E.; Arango, C.S.; Araujo, M.A.; Blundo, C.; Boza, E.T.E.; La Torre, C.M.A.; Gaviria, J.; Gutiérrez, N.; Jørgensen, P.M.; et al. Evaluación delo estado de conservación de los bosques montanos en los Andes tropicales. Rev. Ecos. 2012, 21, 148–166. [Google Scholar]
- Food and Agriculture Organization of the United Nations. State of the Word’s Forest; Food and Agriculture Organization of the United Nations: Roma, Italy, 2011; p. 117. [Google Scholar]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; De la Cruz, M. Deforestation and forest fragmentation in South Ecuador since the 1970s—Losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurance, W.F. Reflections on the tropical deforestation crisis. Biol. Conserv. 1999, 91, 109–117. [Google Scholar] [CrossRef]
- Whitmore, T.; Sayer, J. Deforestation and species extinction in Tropical Forest. In Tropical Deforestation and Species Extinction; IUCN: Gland, Switzerland; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Whitmore, T. Tropical forest disturbance, disappearance, and species loss. In Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; University of Chicago Press: Chicago, IL, USA, 1997; pp. 3–12. [Google Scholar]
- Cayuela, L. Deforestación y fragmentación de bosques tropicales montanos en los Altos de Chiapas, México. Efectos sobre la diversidad de árboles. Ecosistemas 2006, 15, 192–198. [Google Scholar]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Bilogical consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Debinski, D.M.; Holt, R.D. A Survey and Overview of Habitat Fragmentation Experiments. Conserv. Biol. 2000, 14, 342–355. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Rylands, A.B.; Konstant, W.R.; Flick, P.; Pilgrim, J.; Oldfield, S.; Magin, G.; et al. Habitat Loss and Extinction in the Hotspots of Biodiversity. Conserv. Biol. 2002, 16, 909–923. [Google Scholar] [CrossRef] [Green Version]
- Carle, J.; Vuorinen, P.; Del Lungo, A. Status and trends in global forest plantation development. For. Prod. J. 2002, 52, 12–23. [Google Scholar]
- Knoke, T.; Bendix, J.; Pohle, P.; Hamer, U.; Hildebrandt, P.; Roos, K.; Gerique, A.; Sandoval, M.L.; Breuer, L.; Tischer, A.; et al. Afforestation or intense pasturing improve the ecological and economic value of abandoned tropical farmlands. Nat. Commun. 2014, 5, 5612. [Google Scholar] [CrossRef] [Green Version]
- Quichimbo, P.; Jiménez, L.; Veintimilla, D.; Tischer, A.; Günter, S.; Mosandl, R.; Hamer, U. Forest Site Classification in the Southern Andean Region of Ecuador: A Case Study of Pine Plantations to Collect a Base of Soil Attributes. Forests 2017, 8, 473. [Google Scholar] [CrossRef] [Green Version]
- Gradstein, S.R. Epiphytes of tropical montane forests-impact of deforestation and climate change. In The Tropical Mountain Forest, Patterns and Processes in a Biodiversity Hotspot; Gradstein, S.R., Homeier, J., Gansert, D., Eds.; University Press: Göttingen, Germany, 2008; pp. 51–65. [Google Scholar]
- Nöske, N.M.; Hilt, N.; Werner, F.A.; Brehm, G.; Fiedler, K.; Sipman, H.J.; Gradstein, S.R. Disturbance effects on diversity of epiphytes and moths in a montane forest in Ecuador. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Barthlott, W.; Schmit-Neuerburg, V.; Nieder, J.; Engwald, S. Diversity and abundance of vascular epiphytes: A comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecol. 2001, 152, 145–156. [Google Scholar] [CrossRef]
- Krömer, T.; García-Franco, J.G.; Toledo-Aceves, T. Epífitas vasculares como bioindicadores de la calidad forestal: Impacto antrópico sobre su diversidad y composición. In Bioindicadores: Guardianes de Nuestro Futuro Ambiental; González, C.A., Va-llarino, A., Pérez, J.C., Low, A., Eds.; Instituto Nacional de Ecología y Cambio Climático (INECC): Chiapas, Mexico, 2014; pp. 605–623. [Google Scholar]
- Holz, I.; Gradstein, R.S. Cryptogamic epiphytes in primary and recovering upper montane oak forest of Costa Rica-Species richness, community composition and ecology. Plant Ecol. 2005, 178, 89–109. [Google Scholar] [CrossRef]
- Benítez, A.; Prieto, M.; Aragón, G. Large trees and dense canopies: Key factors for maintaining high epiphytic diversity on trunk bases (bryophytes and lichens) in tropical montane forests. Forestry 2015, 88, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, L.D.P.; Pôrto, K.C. Patch size and isolation effects on epiphytic and epiphyllous bryophytes in the fragmented Brazilian Atlantic forest. Biol. Conserv. 2007, 134, 415–427. [Google Scholar] [CrossRef]
- Calviño-Cancela, M.; de Silanes, M.E.L.; Rubido-Bará, M.; Uribarri, J. The potential role of tree plantations in providing habitat for lichen epiphytes. For. Ecol. Manag. 2013, 291, 386–395. [Google Scholar] [CrossRef]
- Ríos, A.I.A.; Moncada, B.; Lücking, R. Epiphyte homogenization and de-diversification on alien Eucalyptus versus native Quercus forest in the Colombian Andes: A case study using lirellate Graphidaceae lichens. Biodivers. Conserv. 2015, 24, 1239–1252. [Google Scholar] [CrossRef]
- González-Montelongo, C.; Pérez-Vargas, I. Looking for a home: Exploring the potential of epiphytic lichens to colonize tree plantations in a Macaronesian laurel forest. For. Ecol. Manag. 2019, 453, 117541. [Google Scholar] [CrossRef]
- Goded, S.; Ekroos, J.; Domínguez, J.; Azcárate, J.G.; Guitián, J.A.; Smith, H.G. Effects of eucalyptus plantations on avian and herb species richness and composition in North-West Spain. Glob. Ecol. Conserv. 2019, 19, e00690. [Google Scholar] [CrossRef]
- Tavares, A.; Beiroz, W.; Fialho, A.; Frazão, F.; Macedo, R.; Louzada, J.; Audino, L. Eucapyptus plantations as hybrid ecosystems: Implication for species conservation in the Brazilian Atlantic. For. Ecol. Manag. 2019, 433, 131–139. [Google Scholar] [CrossRef]
- González-Montelongo, C.; Pérez-Vargas, I. Is an invasive alien tree able to sustain a similar lichen diversity as the native forest? The case of the sweet chestnut (Castanea sativa Mill.) and the laurel forest in Macaronesia. For. Ecol. Manag. 2021, 488, 119009. [Google Scholar] [CrossRef]
- Stephens, S.S.; Wagner, M.R. Forest plantations and biodiversity: A fresh perspective. J. For. 2007, 105, 307–313. [Google Scholar]
- Andersson, M.S.; Gradstein, S.R. Impact of management intensity on non-vascular epiphyte diversity in cacao plantations in western Ecuador. Biodivers. Conserv. 2005, 14, 1101–1120. [Google Scholar] [CrossRef]
- Ariyanti, N.S.; Bos, M.M.; Kartawinata, K.; Tjitrosoedirdjo, S.S.; Guhardja, E.; Gradstein, S.R. Bryophytes on tree trunks in natural forests, selectively logged forests and cacao agroforests in Central Sulawesi, Indonesia. Biol. Conserv. 2008, 141, 2516–2527. [Google Scholar] [CrossRef]
- Sporn, S.G.; Bos, M.M.; Hoffstätter-Müncheberg, M.; Kessler, M.; Gradstein, S.R. Microclimate determines community composition but not richness of epiphytic understory bryophytes of rainforest and cacao agroforests in Indonesia. Funct. Plant Biol. 2009, 36, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Gradstein, S.R.; Sporn, S.G. Land-use change and epiphytic bryophyte diversity in the Tropics. Nova Hedwig. 2010, 138, 311–323. [Google Scholar]
- Ezukanma, I.O.; Tessler, M.; Salaam, A.M.; Chukwuka, K.S.; Ogunniran, A.J. Epiphytic bryophytes of urban agroforests in Ibadan, Southwest Nigeria. J. Bryol. 2019, 41, 341–349. [Google Scholar] [CrossRef]
- Gradstein, S.R.; León-Yánez, S. Liverwort diversity in Polylepis pauta forests of Ecuador under different climatic conditions. Neotrop. Biodivers. 2020, 6, 138–146. [Google Scholar] [CrossRef]
- Gradstein, S.R.; Pócs, R. Diversity of liverworts and hornworts in Polylepis sericea forests in the Andes of Venezuela. Nova Hedwig. 2021, 112, 49–68. [Google Scholar] [CrossRef]
- Gil, J.E.; Morales, M.E. Estratificación vertical de briófitos epífitos encontrados en Quercus humboldtii (Fagaceae) de Boyacá, Colombia. Rev. Biol. Trop. 2014, 62, 719–727. [Google Scholar]
- Lencinas, M.V.; Martínez Pastur, G.; Solán, R.; Gallo, E.; Cellini, J.M. Forest management with variable retention impact over bryophyte communities of Nothofagus pumilio understory. Forstarchiv 2008, 79, 77–82. [Google Scholar]
- Manríquez, M.D.R.T.; Ardiles, V.; Promis, Á.; Herrera, A.H.; Soler, R.; Lencinas, M.V.; Pastur, G.M. Forest canopy-cover composition and landscape influence on bryophyte communities in Nothofagus forests of southern Patagonia. PLoS ONE 2020, 15, e0232922. [Google Scholar] [CrossRef]
- Palacios, B.; Aguirre, Z.; Pucha, D.; Mozha, J.; Armijos, C.; Feijoo, C.; Jumbo, J.; Rojas, A. Factor de forma y productividad de una plantación de Juglans neotropica Diels, establecida en los predios de la Universidad Nacional de Loja. In Memorias I Congreso Internacional de Bosques y Agroforestería; ESPOCH: Riobamba, Ecuador, 2017. [Google Scholar]
- Nieto, V.; Rodríguez, J. Juglans neotropica Diels. In Libro Rojo de Plantas de Colombia. Tomo 4; Corporación Nacional de Investigación Forestal: Bogotá, Colombia, 2002; pp. 258–259. [Google Scholar]
- Ramírez, F.; Kallarackal, J. The phenology of the endangered Nogal (Juglans neotropica Diels) in Bogota and its conservation implications in the urban forest. Urban Ecosyst. 2021, 1–16. [Google Scholar] [CrossRef]
- Aguirre Mendoza, Z.; Reyes Jiménez, B.; Quizhpe Coronel, W.; Cabrera, A. Composición florística, estructura y endemismo del componente leñoso de un bosque montano en el sur del Ecuador. Arnaldoa 2017, 24, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Gradstein, S.R.; Churchill, S.P.; Salazar-Allen, N. Guide to the Bryophytes of Tropical America; New York Botanical Garden: Bronx, NY, USA, 2001. [Google Scholar]
- Gradstein, S.R.; da Costa, D.P. Hepaticae and Anthocerotae of Brazil; New York Botanical Garden Press: Bronx, NY, USA, 2003. [Google Scholar]
- Gradstein, S.R. The Liverworts and Hornworts of Colombia and Ecuador; New York Botanical Garden: Bronx, NY, USA, 2021. [Google Scholar]
- Churchill, S.P.; Linares, E.L. Prodomus Bryologie Novo Granatensis. Introducción a la flora de musgos de Colombia; Tomos I y II. Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia: Bogotá, Colombia, 1995. [Google Scholar]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0. Technological tools. Bull. Ecol. Soc. Am. 2000, 81, 191–197. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Statistics for Biology and Health; Springer: New York, NY, USA, 2007. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan”. In Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 17 January 2019).
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Roberts, D.W. Package ‘labdsv’: Ordination and Multivariate Analysis for Ecology. Version 1.6-1. 2013. Available online: https://cran.r-project.org/web/packages/labdsv/ (accessed on 17 January 2019).
- R Team Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Acebey, A.; Gradstein, S.R.; Krömer, T. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia. J. Trop. Ecol. 2003, 19, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Guerra, G.; Arrocha, C.; Rodríguez, G.; Déleg, J.; Benítez, Á. Briófitos en los troncos de los árboles como indicadores de la alteración en bosques montanos de Panamá. Rev. Biol. Trop. 2020, 68, 492–502. [Google Scholar] [CrossRef]
- Gradstein, S.R.; Culmsee, H. Bryophyte diversity on tree trunks in montane forests of Central Salawesi, Indonesia. Trop. Bryol. 2010, 31, 95–105. [Google Scholar]
- León-Yánez, S.; Valencia, R.; Pitman, N.; Endara, L.; Ulloa Ulloa, C.; Navarrete, H. Libro Rojo de las Plantas Endémicas del Ecuador, 2nd ed.; Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2011. [Google Scholar]



| Plot | Forests | Canopy Openess (%) | MTD (cm) |
|---|---|---|---|
| 1 | BJN | 41.76 | 16.92 |
| 2 | BJN | 38.02 | 24.08 |
| 3 | BJN | 37.96 | 26.47 |
| 4 | BJN | 32.11 | 28.40 |
| 5 | BJN | 28.61 | 25.32 |
| 1 | BS | 21.86 | 17.37 |
| 2 | BS | 27.67 | 19.02 |
| 3 | BS | 26.56 | 24.17 |
| 4 | BS | 26.72 | 19.23 |
| 5 | BS | 24.09 | 14.55 |
| Coef. | ES | Z | p-Value | |
|---|---|---|---|---|
| BJN | 2.23537 | 0.39043 | 5.725 | <0.0001 |
| BS | −0.31825 | 0.12705 | −2.505 | 0.01225 |
| Canopy openness | −0.02284 | 0.01086 | −2.104 | 0.03539 |
| DBH | 0.01085 | 0.00409 | 2.652 | 0.00801 |
| NMDS1 | NMDS2 | r2 | p-Value | |
|---|---|---|---|---|
| Forests | 0.261 | 0.000999 | ||
| BJN | −0.2736 | −0.0361 | ||
| BS | 0.2792 | 0.0368 | ||
| Canopy openness | −0.94759 | −0.3195 | 0.3426 | 0.000999 |
| DBH | −0.5131 | −0.85833 | 0.0643 | 0.042957 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina, J.; Quizhpe, W.; Déleg, J.; Gonzalez, K.; Aguirre, Z.; Aguirre, N.; Montaño, L.; Benítez, Á. Are Juglans neotropica Plantations Useful as a Refuge of Bryophytes Diversity in Tropical Areas? Life 2021, 11, 434. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050434
Medina J, Quizhpe W, Déleg J, Gonzalez K, Aguirre Z, Aguirre N, Montaño L, Benítez Á. Are Juglans neotropica Plantations Useful as a Refuge of Bryophytes Diversity in Tropical Areas? Life. 2021; 11(5):434. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050434
Chicago/Turabian StyleMedina, Jefferson, Wilson Quizhpe, Jorge Déleg, Karina Gonzalez, Zhofre Aguirre, Nikolay Aguirre, Luis Montaño, and Ángel Benítez. 2021. "Are Juglans neotropica Plantations Useful as a Refuge of Bryophytes Diversity in Tropical Areas?" Life 11, no. 5: 434. https://0-doi-org.brum.beds.ac.uk/10.3390/life11050434