
Action of Extracellular Proteases of Aspergillus flavus and Aspergillus ochraceus Micromycetes on Plasma Hemostasis Proteins

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
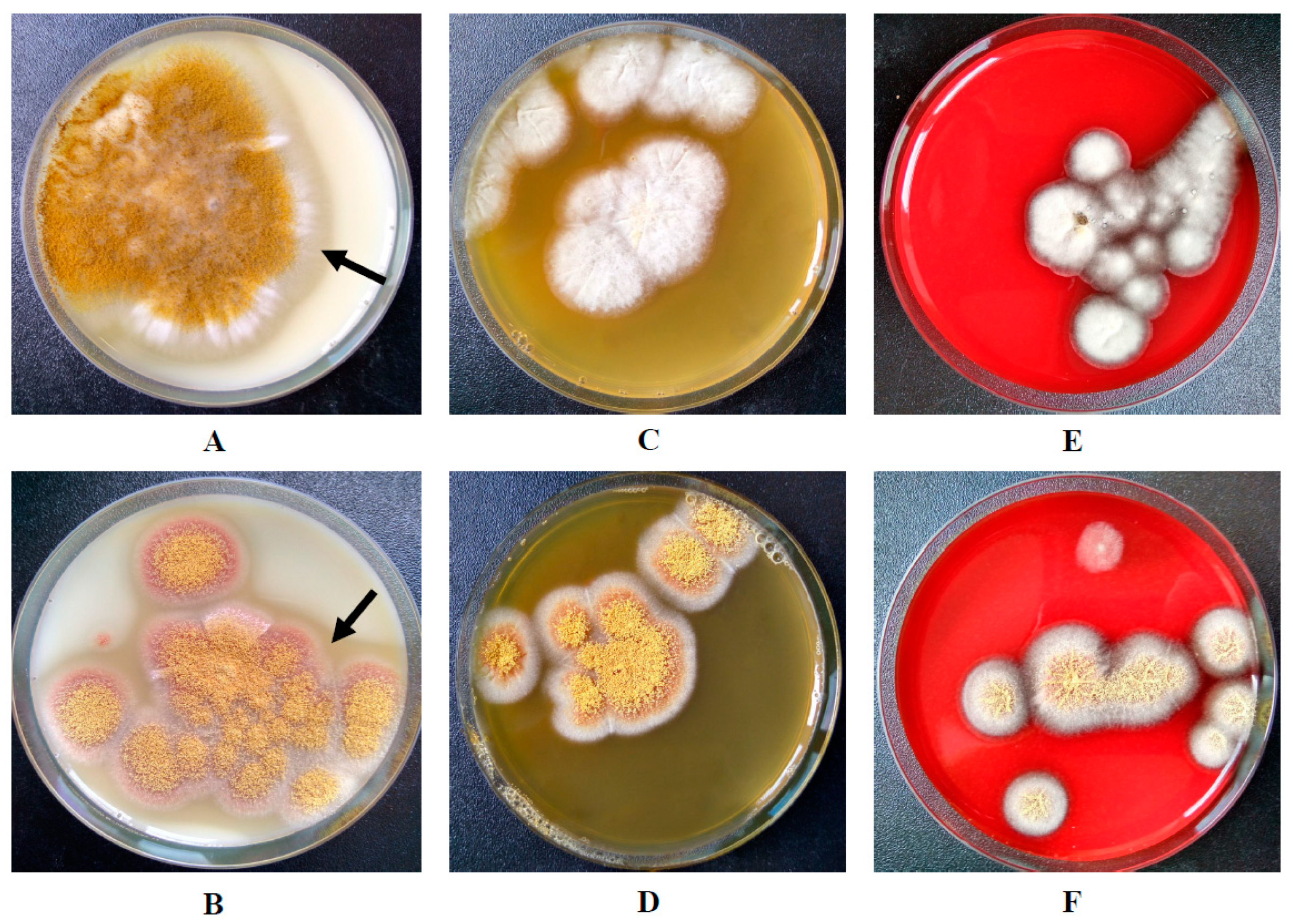
2.1. Fungal Strains
2.2. Proteolytic Potential Determination
2.3. Submerged Cultivation and Proteases Preparations Obtaining
2.4. Fibrin Zymography
2.5. Clotting Assay
2.6. Determination of Proteolytic Activity
2.7. Study of Fungal Proteases Effect on Different Deficient Plasmas
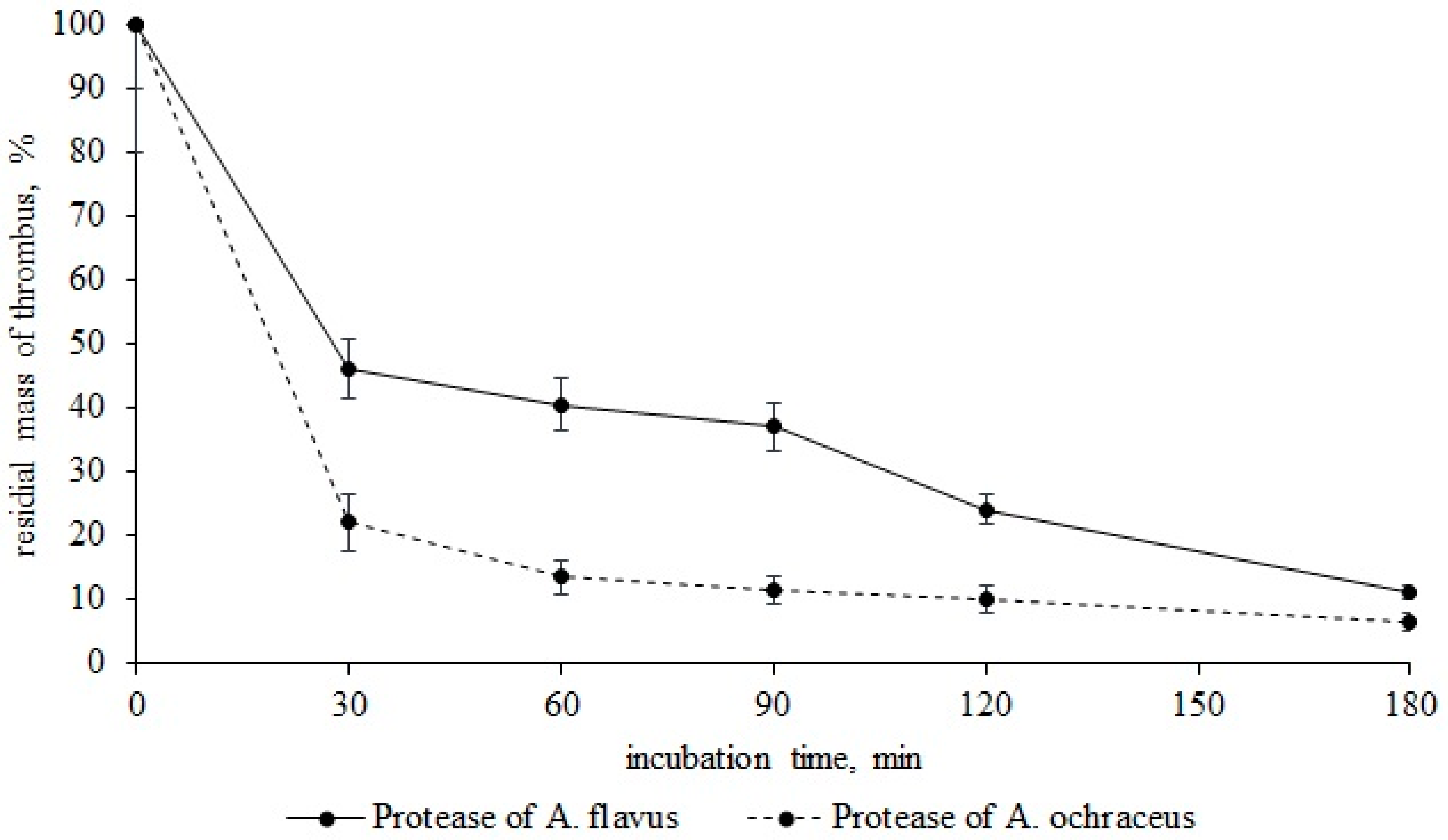
2.8. Thrombolysis Studies
2.9. Protein Determination
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrialy and medicaly important fungal genus Aspergillus. Genome Biol. 2017, 18, 1–45. [Google Scholar] [CrossRef] [Green Version]
- Jeanvoine, A.; Rocchi, S.; Bellanger, A.P.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus: A global phenomenon originating in the environment? Med. Mal. Infect. 2020, 50, 389–395. [Google Scholar] [CrossRef]
- Girard, V.; Dieryckx, C.; Job, C.; Job, D. Secretomes: The fungal strike force. Proteomics 2013, 13, 597–608. [Google Scholar] [CrossRef]
- Frisvad, J.C. Taxonomy, chemodiversity, and chemoconsistency of Aspergillus, Penicillium, and Talaromyces species. Front. Microbiol. 2015, 5, 773. [Google Scholar] [CrossRef] [Green Version]
- di Cologna, N.M.D.; Gómez-Mendoza, D.P.; Zanoelo, F.F.; Giannesi, G.C.; Guimarães, N.C.A.; Moreira, L.R.S.; Filho, E.X.F.; Ricart, C.A.O. Exploring Trichoderma and Aspergillus secretomes: Proteomics approaches for the identification of enzymes of biotechnological interest. Enz. Microb. Technol. 2018, 109, 1–10. [Google Scholar] [CrossRef]
- Lowe, R.G.; Howlett, B.J. Indifferent, affectionate, or deceitful: Lifestyles and secretomes of fungi. PLoS Pathog. 2012, 8, e1002515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvam, R.M.; Nithya, R.; Devi, P.N.; Shree, R.S.; Nila, M.V.; Demonte, N.L.; Thangavel, C.; Maheshwari, J.J.; Lalitha, P.; Prajna, N.V.; et al. Exoproteome of Aspergillus flavus corneal isolates and saprophytes: Identification of proteoforms of an oversecreted alkaline protease. J. Proteom. 2015, 115, 23–35. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, H.K.; Sarkar, B.C. Effect of substrate and fermentation conditions on pectinase and cellulase production by Aspergillus niger NCIM 548 in submerged (SmF) and solid-state fermentation (SSF). Food Sci. Biotechnol. 2011, 20, 1289. [Google Scholar] [CrossRef]
- Reen, F.J.; Phelan, J.P.; Woods, D.F.; Shanahan, R.; Cano, R.; Clarke, S.; McGlacken, G.P.; O’Gara, F. Harnessing bacterial signals for suppression of biofilm formation in the nosocomial fungal pathogen Aspergillus fumigatus. Front. Microbiol. 2016, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Hashemi, S.J.; Zibafar, E.; Zoll, J.; Hedayati, M.T.; Mouton, J.W.; Melchers, W.J.G.; Verweij, P.E. Azole-resistant Aspergillus fumigatus, Iran. Emerg. Infect. Dis. 2013, 19, 832–834. [Google Scholar] [CrossRef] [PubMed]
- Szalewski, D.A.; Hinrichs, V.S.; Zinniel, D.K.; Barletta, R.G. The pathogenicity of Aspergillus fumigatus, drug resistance, and nanoparticle delivery. Can. J. Microbiol. 2018, 64, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Verweij, P.E. Aspergillus fumigatus and pan-azole resistance: Who should be concerned? Curr. Opin. Infect. Dis. 2020, 33, 290–297. [Google Scholar] [CrossRef]
- Denning, D.W.; Park, S.; Lass-Florl, C.; Fraczek, M.G.; Kirwan, M.; Gore, R.; Smith, J.; Bueid, A.; Moore, C.B.; Bowyer, P.; et al. High-frequency triazole resistance found in nonculturable Aspergillus fumigatus from lungs of patients with chronic fungal disease. Clin. Infect. Dis. 2011, 52, 1123–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedayati, M.T.; Mayahi, S.; Denning, D.W. A study on Aspergillus species in houses of asthmatic patients from Sari City, Iran and a brief review of the health effects of exposure to indoor Aspergillus. Environ. Monit. Assess. 2010, 168, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef]
- Osmolovskiy, A.A.; Zvonareva, E.S.; Kreyer, V.G.; Baranova, N.A.; Egorov, N.S. The effect of micromycete extracellular proteases of the Aspergillus genus on the proteins of haemostatic system. Russ. J. Bioorg. Chem. 2014, 40, 634–639. [Google Scholar] [CrossRef]
- Sharkova, T.S.; Kurakov, A.V.; Osmolovskiy, A.A.; Matveeva, E.O.; Kreyer, V.G.; Baranova, N.A.; Egorov, N.S. Screening of producers of proteases with fibrinolytic and collagenolytic activities among micromycete. Microbiology (Mikrobiologiya) 2015, 84, 359–364. [Google Scholar]
- Farnell, E.; Rousseau, K.; Thornton, D.J.; Bowyer, P.; Herrick, S.E. Expression and secretion of Aspergillus fumigatus proteases are regulated in response to different protein substrates. Fungal Biol. 2012, 116, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Osmolovskiy, A.A.; Lukianova, A.A.; Zvonareva, E.S.; Kreyer, V.G.; Baranova, N.A.; Egorov, N.S. Combined microbiological approach to screening of producers of proteases with hemostasis system proteins activity among micromycetes. Biotechbol. Rep. 2018, 19, e00265. [Google Scholar] [CrossRef]
- Vesterberg, O. Isoelectric focusing of proteins. Methods Enzymol. 1971, 22, 389–412. [Google Scholar]
- Davis, B.J. Disc Electrophoresis. II. Method and application to human serum proteins. Ann. N. Y. Acad. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef]
- Chung, D.-M.; Choi, N.S.; Maeng, P.J.; Chun, H.K.; Kim, S.-H. Purification and characterization of a novel fibrinolytic enzyme from chive (Allium tuberosum). Food Sci. Biotechnol. 2012, 126, 349–355. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, D.; Wang, Z.; Hou, J.; Tyagi, R.; Liang, Y.; Hu, Y. Purification and characterization of a novel, highly potent fibrinolytic enzyme from Bacillus subtilis DC27 screened from Douchi, a traditional Chinese fermented soybean food. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmolovskiy, A.A.; Popova, E.A.; Kreyer, V.G.; Baranova, N.A.; Egorov, N.S. Fibrinolytic and collagenolytic activity of extracellular proteinases of the strains of micromycetes Aspergillus ochraceus L-1 and Aspergillus ustus 1. Moscow Univ. Biol. Sci. Bull. 2016, 71, 62–66. [Google Scholar] [CrossRef]
- Kotb, E.; Helal, G.E.-D.A.; Edries, F.M. Screening for fibrinolytic filamentous fungi and enzymatic properties of the most potent producer, Aspergillus brasiliensis AUMC 9735. Biologia 2015, 70, 1565–1574. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Orekhova, A.V.; Osmolovskiy, A.A.; Kreyer, V.G.; Baranova, N.A.; Egorov, N.S. Possibility for application of extracellular protease of micromycete Aspergillus ochraceus for determining factor X content in human blood plasma. Moscow Univ. Biol. Sci. Bull. 2019, 74, 117–120. [Google Scholar] [CrossRef]
- Osmolovskiy, A.A.; Kreier, V.G.; Baranova, N.A.; Egorov, N.S. Properties of extracellular plasmin-like proteases of Aspergillus ochraceus micromycete. Appl. Biochem. Microbiol. 2017, 53, 429–434. [Google Scholar] [CrossRef]
- Galiakberova, A.A.; Bednenko, D.M.; Kreyer, V.G.; Osmolovskiy, A.A.; Egorov, N.S. Formation and properties of extracellular proteinase of micromycete Aspergillus flavus O-1, active toward fibrillary proteins. Appl. Biochem. Microbiol. 2021, 57, 586–593. [Google Scholar]
- Richard, J.L.; Thurston, J.R. Rapid hematogenous dissemination of Aspergillus fumigatus and A. flavus spores in Turkey poults following aerosol exposure. Avian Dis. 1983, 27, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zhao, C.H.; Yin, X.Y.; Chen, Y.L.; Zhai, H.Y.; Xu, C.W.; Wang, Y.; Ge, F.J.; Xu, J.M. Aspergillus niger bloodstream infection in gastric cancer after common hepatic artery embolization: A case report. Exp. Ther. Med. 2017, 14, 1427–1432. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Xu, L.; Yu, S.; Hong, W.; Huang, M.; Xu, P. Therapeutics targeting the fibrinolytic system. Exp. Mol. Med. 2020, 52, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, T.P.; Conniff, A.E.S.; Moura, J.A.S.; Batista, J.M.S.; Costa, R.M.P.B.; Porto, C.S.; Takaki, G.M.C.; Porto, T.S.; Porto, A.L.F. Protease from Mucor subtilissimus UCP 1262: Evaluation of several specific protease activities and purification of a fibrinolytic enzyme. An. Acad. Bras. Cienc. 2020, 92, e20200882. [Google Scholar] [CrossRef]
- Kornienko, E.I.; Osmolovskiy, A.A.; Kreyer, V.G.; Baranova, N.A.; Kotova, I.B.; Egorov, N.S. Characteristics and properties of the complex of proteolytic enzymes of the thrombolytic action of the micromycete Sarocladium strictum. Appl. Biochem. Microbiol. 2021, 57, 57–64. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protease | Albuminolysis, UTyr/mg of Protein | Hemoglobinolysis, UTyr/mg of Protein | |
|---|---|---|---|
| Human Albumin | Bovine Albumin | ||
| A. flavus | 482.4 | 350.8 | 1064.2 |
| A. ochraceus | 357.3 | 603.7 | 887.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osmolovskiy, A.A.; Schmidt, L.; Orekhova, A.V.; Komarevtsev, S.K.; Kreyer, V.G.; Shabunin, S.V.; Egorov, N.S. Action of Extracellular Proteases of Aspergillus flavus and Aspergillus ochraceus Micromycetes on Plasma Hemostasis Proteins. Life 2021, 11, 782. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080782
Osmolovskiy AA, Schmidt L, Orekhova AV, Komarevtsev SK, Kreyer VG, Shabunin SV, Egorov NS. Action of Extracellular Proteases of Aspergillus flavus and Aspergillus ochraceus Micromycetes on Plasma Hemostasis Proteins. Life. 2021; 11(8):782. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080782
Chicago/Turabian StyleOsmolovskiy, Alexander A., Laura Schmidt, Anastasia V. Orekhova, Sergey K. Komarevtsev, Valeriana G. Kreyer, Sergey V. Shabunin, and Nikolay S. Egorov. 2021. "Action of Extracellular Proteases of Aspergillus flavus and Aspergillus ochraceus Micromycetes on Plasma Hemostasis Proteins" Life 11, no. 8: 782. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080782