
Peptide Bond Formation between Aminoacyl-Minihelices by a Scaffold Derived from the Peptidyl Transferase Center

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
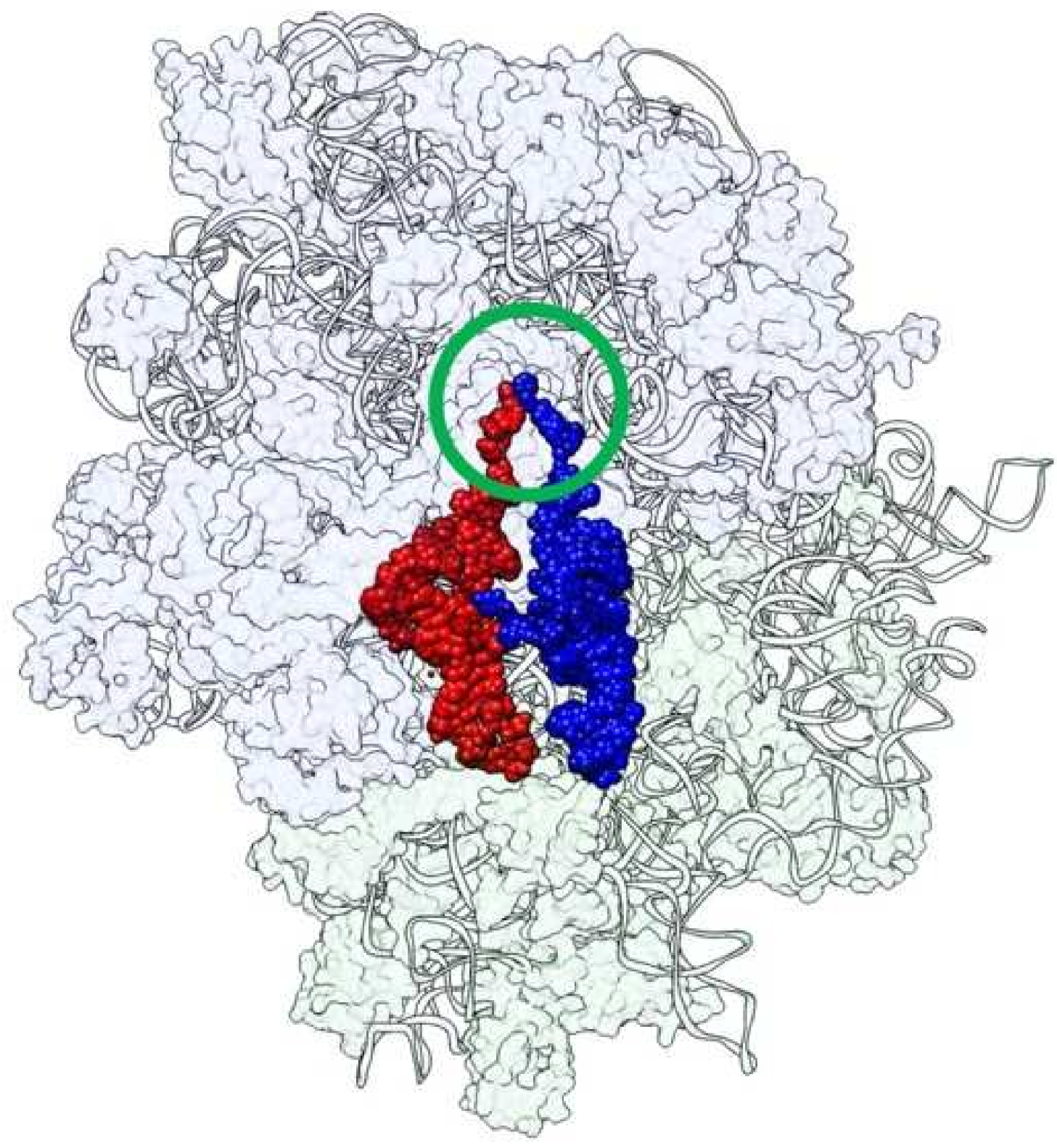
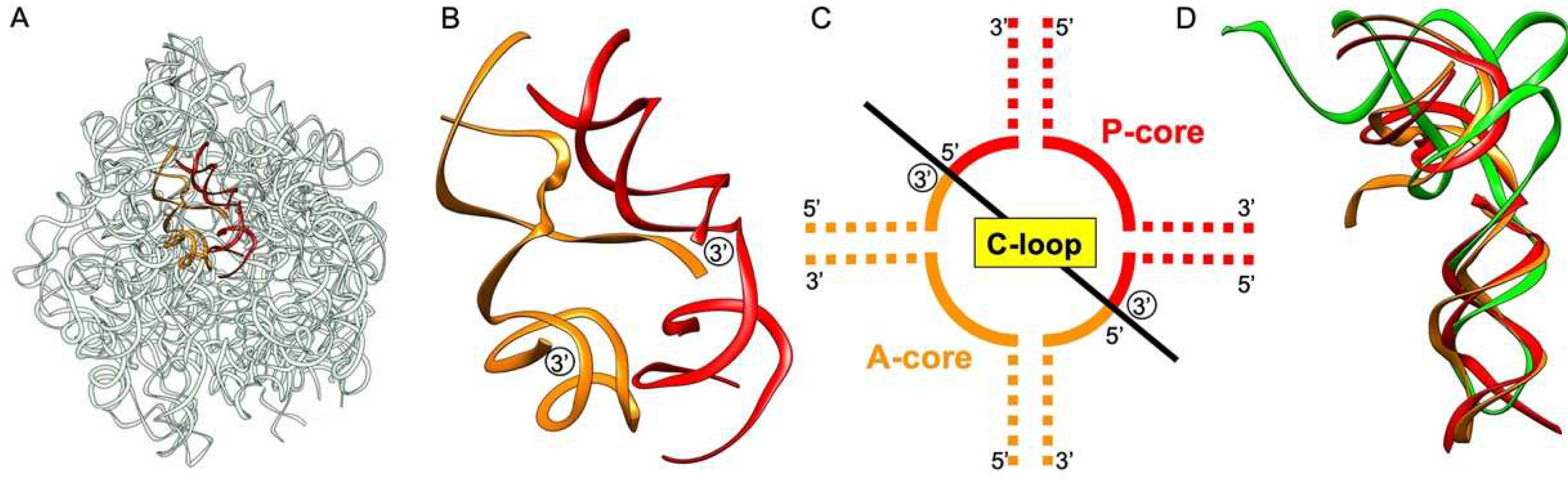
:1. Introduction
2. Materials and Methods
2.1. In Vitro Transcription
2.2. Expression and Purification of E. coli AlaRS442N
2.3. Electrophoretic Mobility Shift Assay
2.4. Preparation of Alanyl-MinihelixAla
2.5. Peptidyl Transfer Reaction Using Alanyl-MinihelixAla
2.6. Mass Spectrometry
3. Results
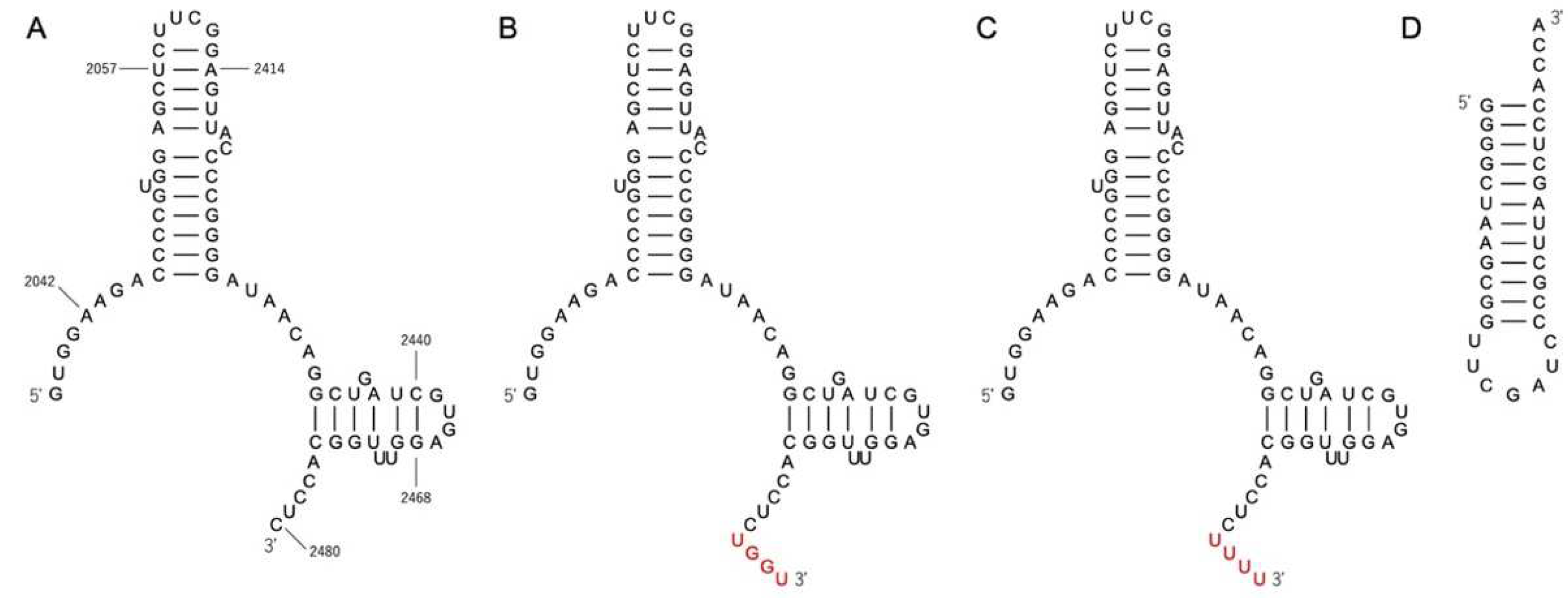
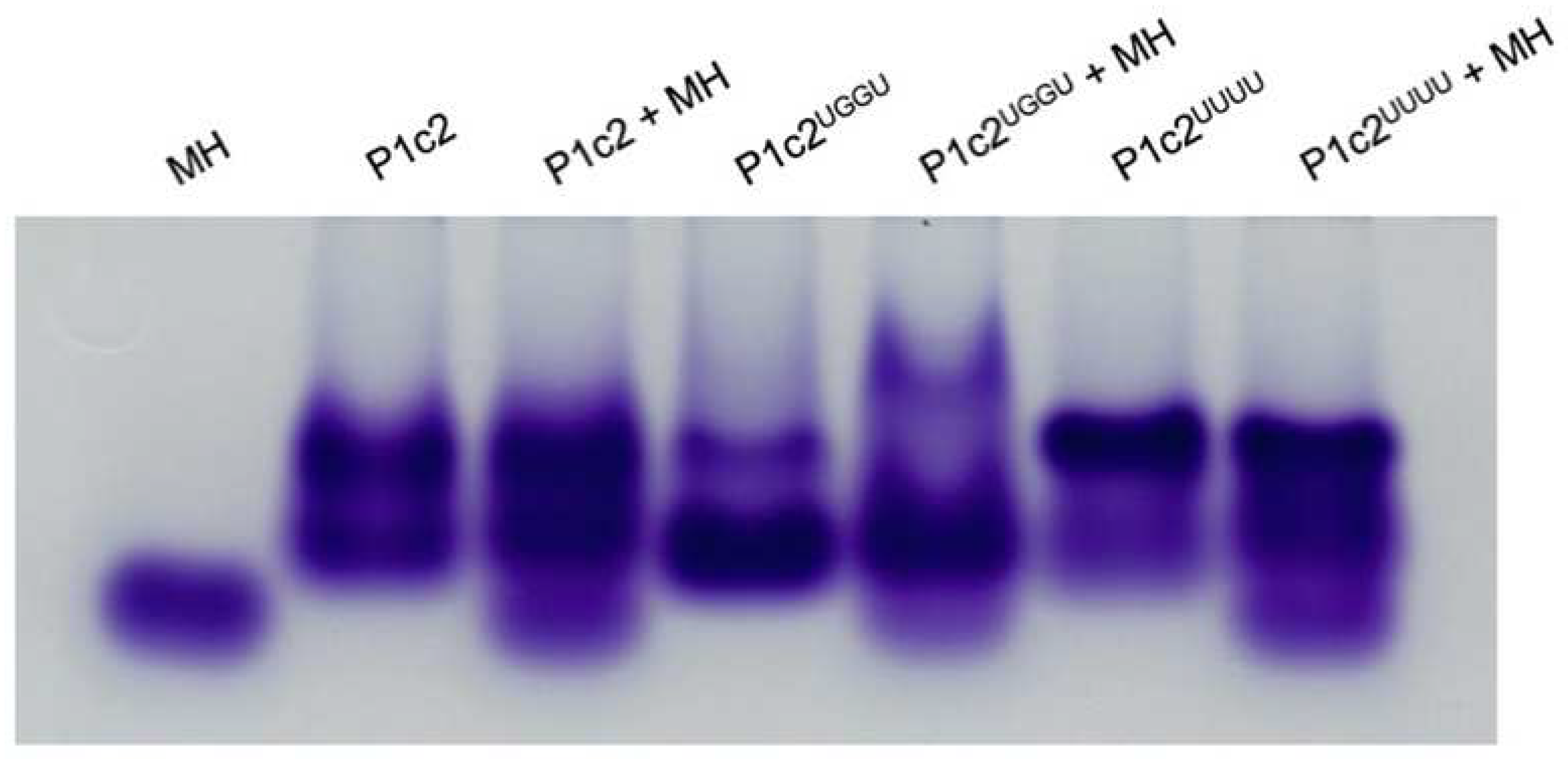
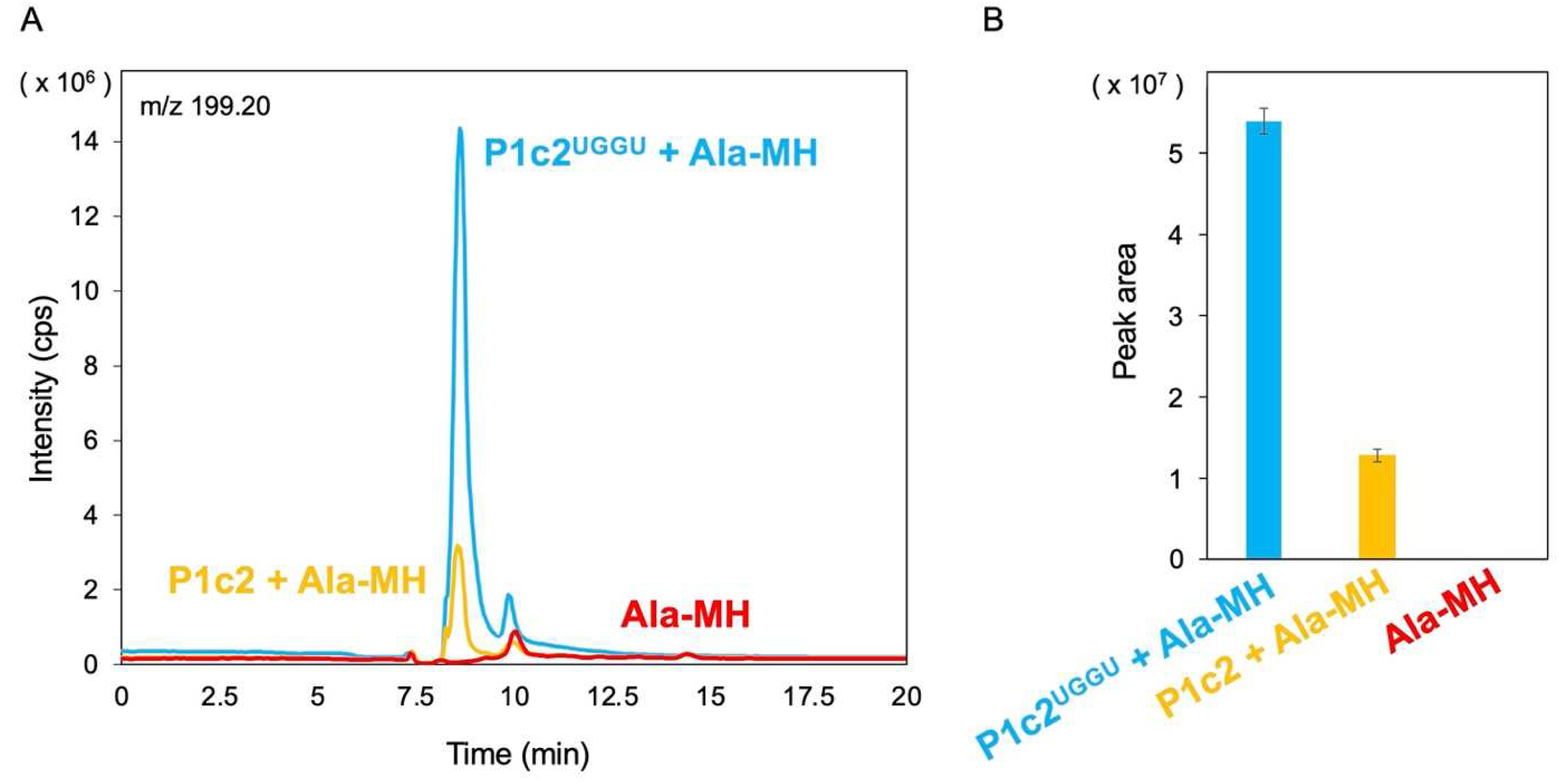
3.1. Gel Shift Assay of P1c2, P1c2UGGU, P1c2UUUU, and MinihelixAla
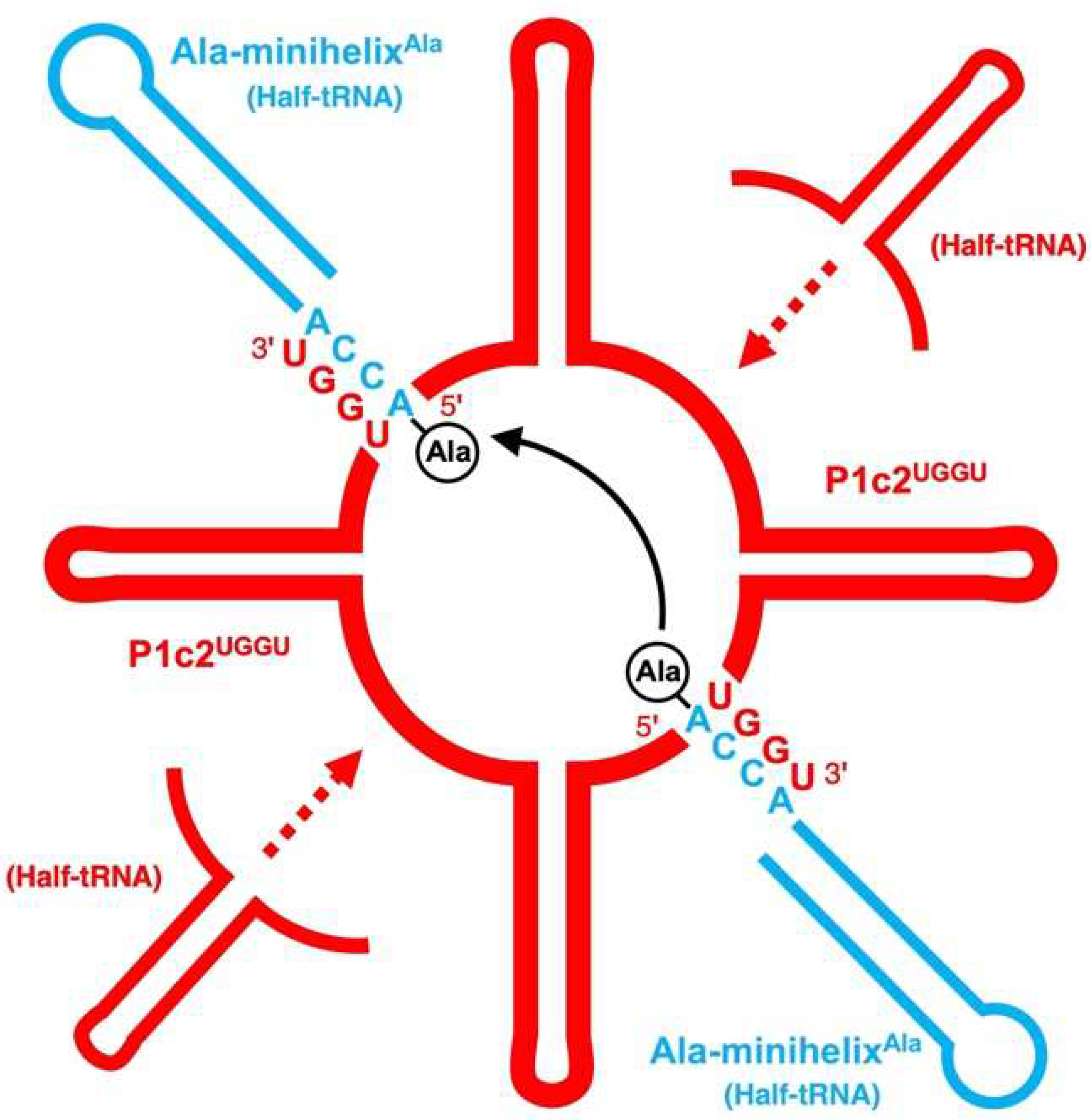
3.2. Attempt and Detection of Peptide Bond Formation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Noller, H.F. On the origin of the ribosome: Coevolution of subdomains of tRNA and rRNA. In The RNA World; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1993; pp. 137–156. [Google Scholar]
- Tamura, K.; Alexander, R.W. Peptide synthesis through evolution. Cell. Mol. Life Sci. 2004, 61, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Origins and early evolution of the tRNA molecule. Life 2015, 5, 1687–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Hansen, J.; Ban, N.; Moore, P.B.; Steitz, T.A. The structural basis of ribosome activity in peptide bond synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef] [Green Version]
- Noller, H.F.; Hoffarth, V.; Zimniak, L. Unusual resistance of peptidyl transferase to protein extraction procedures. Science 1992, 256, 1416–1419. [Google Scholar] [CrossRef]
- Cech, T.R. The ribosome is a ribozyme. Science 2000, 289, 878–879. [Google Scholar] [CrossRef]
- Tamura, K. The role of the CCA sequence of tRNA in the peptidyl transfer reaction. FEBS Lett. 1994, 353, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, R.M.; Weixlbaumer, A.; Loakes, D.; Kelley, A.C.; Ramakrishnan, V. Insights into substrate stabilization from snapshots of the peptidyl transferase center of the intact 70S ribosome. Nat. Struct. Mol. Biol. 2009, 16, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Rugby fans Rush to See Buddha Statue in Gifu Resembling Popular Kicker. The Japan Times. 22 November 2015. Available online: https://www.japantimes.co.jp/news/2015/11/22/national/rugby-fans-rush-to-buddha-statue-resembling-popular-player/ (accessed on 22 March 2022).
- Tamura, K. Ribosome evolution: Emergence of peptide synthesis machinery. J. Biosci. 2011, 36, 921–928. [Google Scholar] [CrossRef]
- Agmon, I. The dimeric proto-ribosome: Structural details and possible implications on the origin of life. Int. J. Mol. Sci. 2009, 10, 2921–2934. [Google Scholar] [CrossRef] [Green Version]
- Agmon, I.; Bashan, A.; Zarivach, R.; Yonath, A. Symmetry at the active site of the ribosome: Structural and functional implications. Biol. Chem. 2005, 386, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Davidovich, C.; Belousoff, M.; Wekselman, I.; Shapira, T.; Krupkin, M.; Zimmerman, E.; Bashan, A.; Yonath, A. The proto-ribosome: An ancient nano-machine for peptide bond formation. Isr. J. Chem. 2010, 50, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Agmon, I.; Davidovich, C.; Bashan, A.; Yonath, A. Identification of the prebiotic translation apparatus within the contemporary ribosome. Nat. Prec. 2009. [Google Scholar] [CrossRef]
- Bose, T.; Fridkin, G.; Davidovich, C.; Krupkin, M.; Dinger, N.; Falkovich, A.H.; Peleg, Y.; Agmon, I.; Bashan, A.; Yonath, A. Origin of life: Protoribosome forms peptide bonds and links RNA and protein dominated worlds. Nucleic Acids Res. 2022, 50, 1815–1828. [Google Scholar] [CrossRef]
- Schmeing, T.M.; Seila, A.C.; Hansen, J.L.; Freeborn, B.; Soukup, J.K.; Scaringe, S.A.; Strobel, S.A.; Moore, P.B.; Steitz, T.A. A pre-translocational intermediate in protein synthesis observed in crystals of enzymatically active 50S subunits. Nat. Struct. Biol. 2002, 9, 225–230. [Google Scholar] [CrossRef]
- Schimmel, P.; Alexander, R. Diverse RNA substrates for aminoacylation: Clues to origins? Proc. Natl. Acad. Sci. USA 1998, 95, 10351–10353. [Google Scholar] [CrossRef] [Green Version]
- Libas de Pouplana, R.; Turner, R.J.; Steer, B.A.; Schimmel, P. Genetic code origins: tRNAs older than their synthetases? Proc. Natl. Acad. Sci. USA 1998, 95, 11295–11300. [Google Scholar] [CrossRef] [Green Version]
- Sampson, J.R.; Uhlenbeck, O.C. Biochemical and physical characterization of an unmodified yeast phenylalanine transfer RNA transcribed in vitro. Proc. Natl. Acad. Sci. USA 1988, 85, 1033–1037. [Google Scholar] [CrossRef] [Green Version]
- Ohmori, S.; Wani, M.; Kitabatake, S.; Nakatsugawa, Y.; Ando, T.; Umehara, T.; Tamura, K. RNA aptamers for a tRNA-binding protein from Aeropyrum pernix with homologous counterparts distributed throughout evolution. Life 2020, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Mutsuro-Aoki, H.; Hamachi, K.; Kurihara, R.; Tamura, K. Aminoacylation of short hairpin RNAs through kissing-loop interactions indicates evolutionary trend of RNA molecules. Biosystems 2020, 197, 104206. [Google Scholar] [CrossRef]
- Sardesai, N.Y.; Green, R.; Schimmel, P. Efficient 50S ribosome-catalyzed peptide bond synthesis with an aminoacyl minihelix. Biochemistry 1999, 38, 12080–12088. [Google Scholar] [CrossRef] [PubMed]
- Schreier, A.A.; Schimmel, P.R. Transfer ribonucleic acid synthetase catalyzed deacylation of aminoacyl transfer ribonucleic acid in the absence of adenosine monophosphate and pyrophosphate. Biochemistry 1972, 11, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Francklyn, C.; Schimmel, P. Aminoacylation of RNA minihelices with alanine. Nature 1989, 337, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.P.; Martinis, S.A.; Schimmel, P. RNA tetraloops as minimalist substrates for aminoacylation. Biochemistry 1992, 31, 4931–4936. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Chong, Y.E.; Shapiro, R.; Beebe, K.; Yang, X.L.; Schimmel, P. Paradox of mistranslation of serine for alanine caused by AlaRS recognition dilemma. Nature 2009, 462, 808–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agmon, I. Sequence complementarity at the ribosomal Peptidyl Transferase Centre implies self-replicating origin. FEBS Lett. 2017, 591, 3252–3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimmel, P.; Giegé, R.; Moras, D.; Yokoyama, S. An operational RNA code for amino acids and possible relationship to genetic code. Proc. Natl. Acad. Sci. USA 1993, 90, 8763–8768. [Google Scholar] [CrossRef] [Green Version]
- Schimmel, P.; Ribas de Pouplana, L. Transfer RNA: From minihelix to genetic code. Cell 1995, 81, 983–986. [Google Scholar] [CrossRef] [Green Version]
- Di Giulio, M. On the origin of protein synthesis: A speculative model based on hairpin RNA structures. J. Theor. Biol. 1994, 171, 303–308. [Google Scholar] [CrossRef]
- Di Giulio, M. Was it an ancient gene codifying for a hairpin RNA that, by means of direct duplication, gave rise to the primitive tRNA molecule? J. Theor. Biol. 1995, 177, 95–101. [Google Scholar] [CrossRef]
- Ramaswamy, K.; Wei, K.; Suga, H. Minihelix-loop RNAs: Minimal structures for aminoacylation catalysts. Nucleic Acids Res. 2002, 30, 2162–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Burton, Z.F. Evolution of the genetic code. Transcription 2021, 12, 28–53. [Google Scholar] [CrossRef] [PubMed]
- Burton, Z.F. The 3-minihelix tRNA evolution theorem. J. Mol. Evol. 2020, 88, 234–242. [Google Scholar] [CrossRef]
- Tamura, K.; Schimmel, P. Chiral-selective aminoacylation of an RNA minihelix. Science 2004, 305, 1253. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Schimmel, P.R. Chiral-selective aminoacylation of an RNA minihelix: Mechanistic features and chiral suppression. Proc. Natl. Acad. Sci. USA 2006, 103, 13750–13752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, T.; Takahashi, S.; Tamura, K. Principles of chemical geometry underlying chiral selectivity in RNA minihelix aminoacylation. Nucleic Acids Res. 2018, 46, 11144–11152. [Google Scholar] [CrossRef] [PubMed]
- Arutaki, M.; Kurihara, R.; Matsuoka, T.; Inami, A.; Tokunaga, K.; Ohno, T.; Takahashi, H.; Takano, H.; Ando, T.; Mutsuro-Aoki, H.; et al. G:U-independent RNA minihelix aminoacylation by Nanoarchaeum equitans alanyl-tRNA synthetase: An insight into the evolution of aminoacyl-tRNA synthetases. J. Mol. Evol. 2020, 88, 501–509. [Google Scholar] [CrossRef]
- Bashan, A.; Agmon, I.; Zarivatch, R.; Schluenzen, F.; Harms, J.M.; Berisio, R.; Bartels, H.; Franceschi, F.; Auerbach, T.; Hansen, H.A.; et al. Structural basis of the ribosomal machinery for peptide bond formation, translocation, and nascent chain progression. Mol. Cell 2003, 11, 91–102. [Google Scholar] [CrossRef]
- Gartner, Z.J.; Liu, D.R. The generality of DNA-templated synthesis as a basis for evolving non-natural small molecules. J. Am. Chem. Soc. 2001, 123, 6961–6963. [Google Scholar] [CrossRef] [Green Version]
- Shimada, J.; Yamakawa, H. Ring-closure probabilities for twisted wormlike chains. Application to DNA. Macromolecules 1984, 17, 689–698. [Google Scholar] [CrossRef]
- Tamura, K.; Schimmel, P. Oligonucleotide-directed peptide synthesis in a ribosome- and ribozyme-free system. Proc. Natl. Acad. Sci. USA 2001, 98, 1393–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, T.; Suzuki, T. Ribosomal RNAs are tolerant toward genetic insertions: Evolutionary origin of the expansion segments. Nucleic Acids Res. 2008, 36, 3539–3551. [Google Scholar] [CrossRef] [PubMed]
- Sievers, A.; Beringer, M.; Rodnina, M.V.; Wolfenden, R. The ribosome as an entropy trap. Proc. Natl. Acad. Sci. USA 2004, 101, 7897–7901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlacher, M.D.; Polacek, N. Ribosomal catalysis: The evolution of mechanistic concepts for peptide bond formation and peptidyl-tRNA hydrolysis. RNA Biol. 2008, 5, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Pech, M.; Nierhaus, K.H. Ribosomal peptide-bond formation. Chem. Biol. 2008, 15, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Bloch, D.P.; McArthur, B.; Widdowson, R.; Spector, D.; Guimarães, R.C.; Smith, J. tRNA-rRNA sequence homologies: A model for the generation of a common ancestral molecule and prospects for its reconstruction. Orig. Life Evol. Biosph. 1984, 14, 571–578. [Google Scholar] [CrossRef]
- Guimarães, R.C. Self-referential encoding on modules of anticodon pairs-roots of the biological flow system. Life 2017, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Zöllner, N. Purine and pyrimidine metabolism. Proc. Nutr. Soc. 1982, 41, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P.E. Peptide nucleic acid (PNA): A model structure for the primordial genetic material? Orig. Life Evol. Biosph. 1993, 23, 323–327. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawabata, M.; Kawashima, K.; Mutsuro-Aoki, H.; Ando, T.; Umehara, T.; Tamura, K. Peptide Bond Formation between Aminoacyl-Minihelices by a Scaffold Derived from the Peptidyl Transferase Center. Life 2022, 12, 573. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040573
Kawabata M, Kawashima K, Mutsuro-Aoki H, Ando T, Umehara T, Tamura K. Peptide Bond Formation between Aminoacyl-Minihelices by a Scaffold Derived from the Peptidyl Transferase Center. Life. 2022; 12(4):573. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040573
Chicago/Turabian StyleKawabata, Mai, Kentaro Kawashima, Hiromi Mutsuro-Aoki, Tadashi Ando, Takuya Umehara, and Koji Tamura. 2022. "Peptide Bond Formation between Aminoacyl-Minihelices by a Scaffold Derived from the Peptidyl Transferase Center" Life 12, no. 4: 573. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040573