
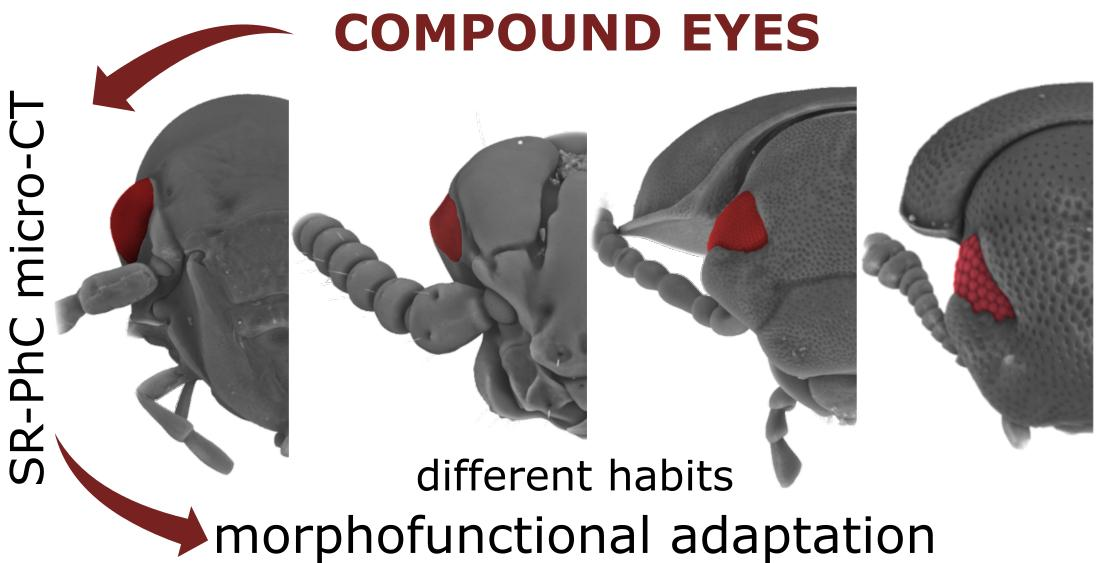
Exploring Compound Eyes in Adults of Four Coleopteran Species Using Synchrotron X-ray Phase-Contrast Microtomography (SR-PhC Micro-CT)

 , ,
, ,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Sample Preparation
2.3. Phase-Contrast Micro-Computed Tomography (PhC micro-CT) and Data Acquisition
2.4. Computer-Based 3D Reconstruction and Segmentation
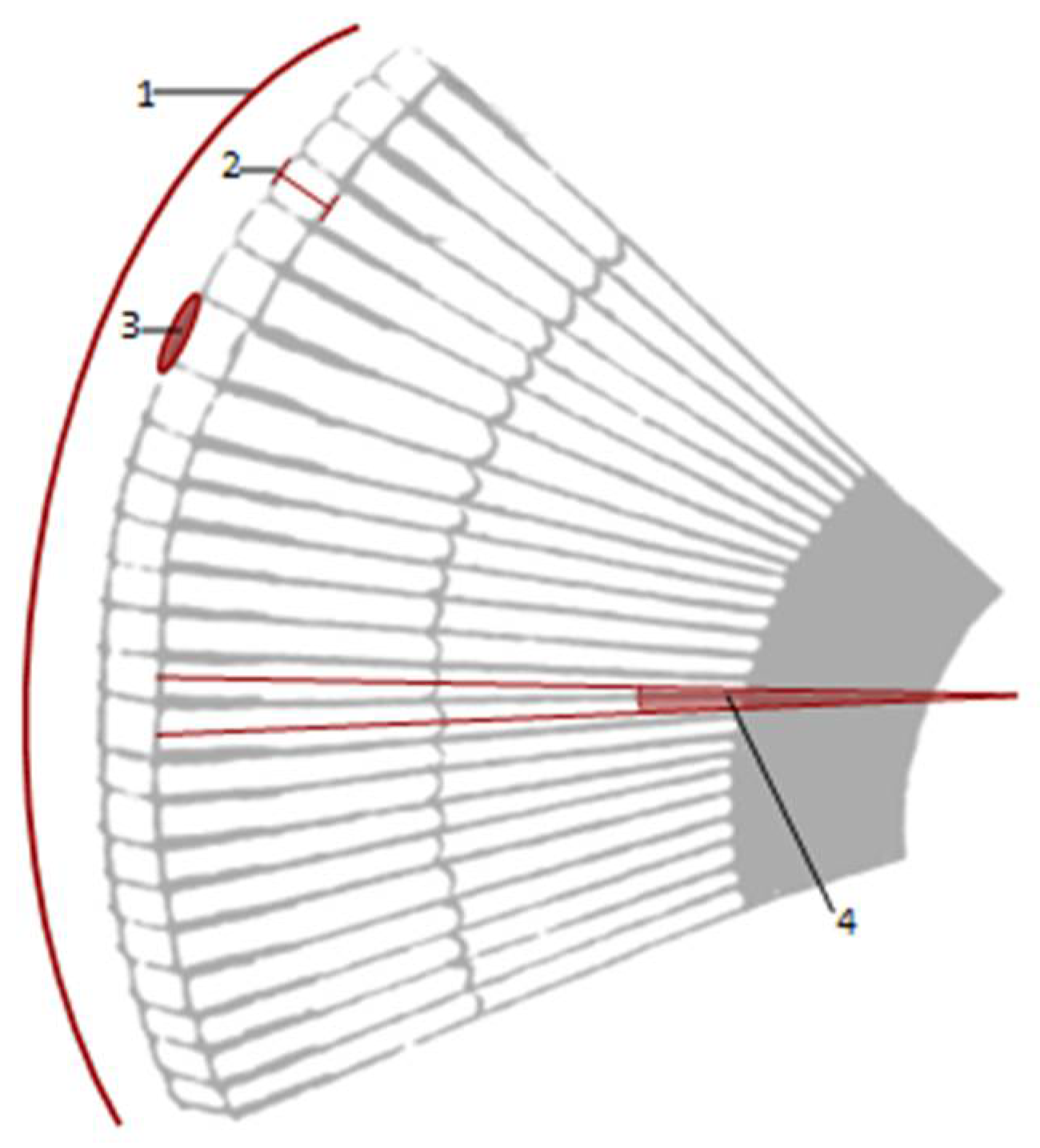
2.5. Image Analyses and Measurements
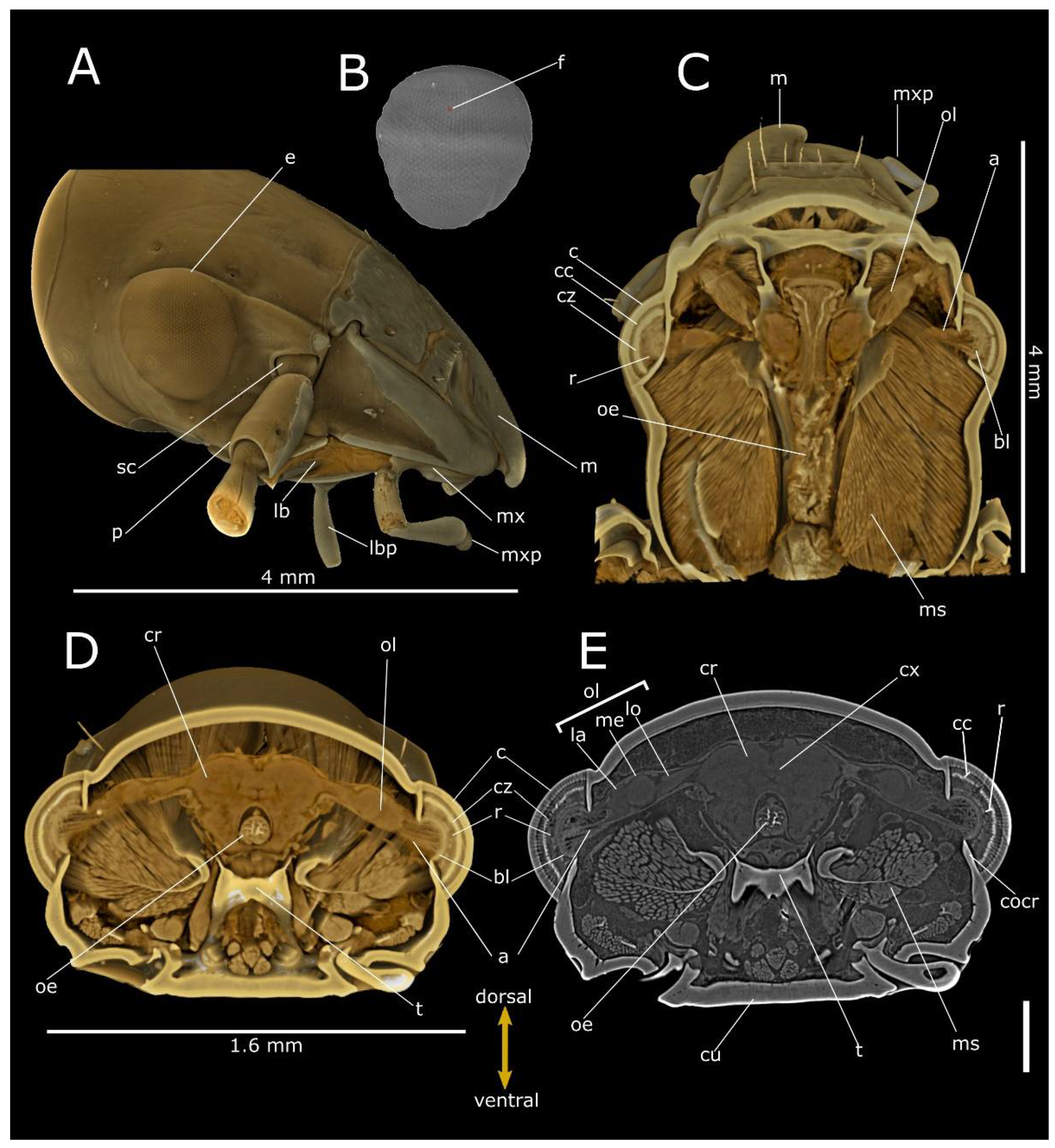
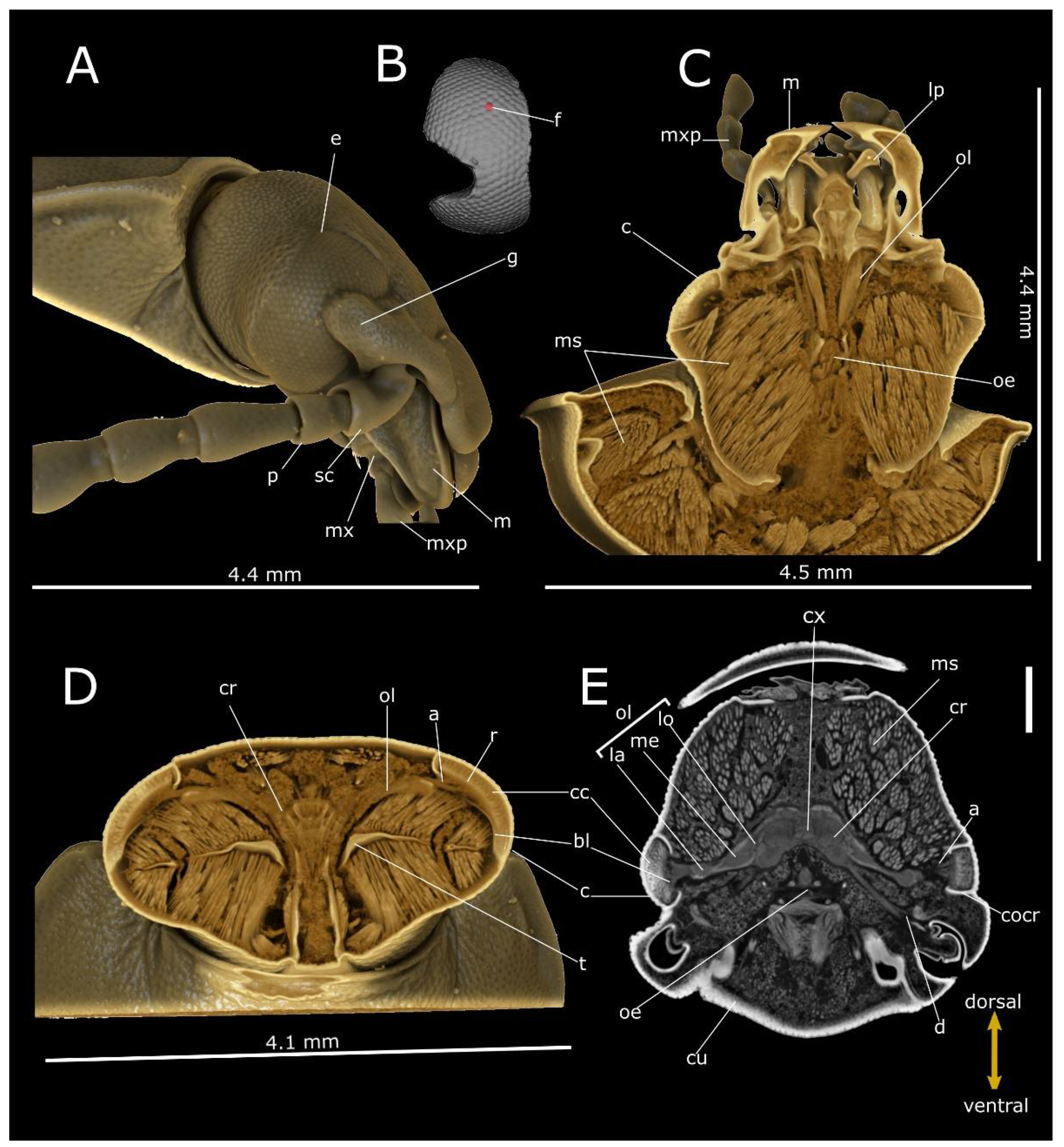
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hörnschemeyer, T.; Beutel, R.G.; Pasop, F. Head structures of Priacma serrata Leconte (Coleptera, Archostemata) inferred from X-ray tomography. J. Morphol. 2002, 252, 298–314. [Google Scholar] [CrossRef]
- Betz, O.; Wegst, U.; Weide, D.; Heethoff, M.; Helfen, L.; LEE, W.; Cloetens, P. Imaging applications of synchrotron X-ray phase-contrast microtomography in biological morphology and biomaterials science. I. General aspects of the technique and its advantages in the analysis of millimetre-sized arthropod structure. J. Microsc. 2007, 227, 51–71. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, K.; Zhu, P.; Wu, Z.; Zhou, H. 3D configuration of mandibles and controlling muscles in rove beetles based on micro-CT technique. Anal. Bioanal. Chem. 2011, 401, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, F.; Gorb, S.N.; Büsse, S. Comparative morphology of the thorax musculature of adult Anisoptera (Insecta: Odonata): Functional aspects of the flight apparatus. Arthropod Struct. Dev. 2018, 47, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Bernhardt, G.; Raine, N.E.; Abel, R.L.; Sykes, D.; Ahmed, F.; Pedroso, I.; Gill, R.J. Exploring miniature insect brains using micro-CT scanning techniques. Sci. Rep. 2016, 6, 21768. [Google Scholar] [CrossRef] [PubMed]
- Donato, S.; Vommaro, M.L.; Tromba, G.; Giglio, A. Synchrotron X-ray phase contrast micro tomography to explore the morphology of abdominal organs in Pterostichus melas italicus Dejean, 1828 (Coleoptera, Carabidae). Arthropod Struct. Dev. 2021, 62, 101044. [Google Scholar] [CrossRef]
- Alba-Alejandre, I.; Alba-Tercedor, J.; Vega, F.E. Anatomical study of the coffee berry borer (Hypothenemus hampei) using micro-computed tomography. Sci. Rep. 2019, 9, 17150. [Google Scholar] [CrossRef]
- Küpper, S.C.; Klass, K.D.; Uhl, G.; Eberhard, M.J.B. Comparative morphology of the internal female genitalia in two species of Mantophasmatodea. Zoomorphology 2019, 138, 73–83. [Google Scholar] [CrossRef]
- Alba-Alejandre, I.; Alba-Tercedor, J.; Hunter, W.B. Anatomical study of the female reproductive system and bacteriome of Diaphorina citri Kuwayama, (Insecta: Hemiptera, Liviidae) using micro-computed tomography. Sci. Rep. 2020, 10, 7161. [Google Scholar] [CrossRef]
- Alba-Alejandre, I.; Hunter, W.B.; Alba-Tercedor, J. Micro-CT study of male genitalia and reproductive system of the Asian citrus psyllid, Diaphorina citri Kuwayama, 1908 (Insecta: Hemiptera, Liviidae). PLoS ONE 2018, 13, e0202234. [Google Scholar] [CrossRef]
- Mattei, A.L.; Riccio, M.L.; Avila, F.W.; Wolfner, M.F. Integrated 3D view of postmating responses by the Drosophila melanogaster female reproductive tract, obtained by micro-computed tomography scanning. Proc. Natl. Acad. Sci. USA 2015, 112, 8475–8480. [Google Scholar] [CrossRef] [PubMed]
- Dierick, M.; Cnudde, V.; Masschaele, B.; Vlassenbroeck, J.; Van Hoorebeke, L.; Jacobs, P. Micro-CT of fossils preserved in amber. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2007, 580, 641–643. [Google Scholar] [CrossRef]
- Pohl, H.; Wipfler, B.; Grimaldi, D.; Beckmann, F.; Beutel, R.G. Reconstructing the anatomy of the 42-million-year-old fossil †Mengea tertiaria (Insecta, Strepsiptera). Naturwissenschaften 2010, 97, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Kehlmaier, C.; Dierick, M.; Skevington, J.H. Micro-CT studies of amber inclusions reveal internal genitalic features of big-headed flies, enabling a systematic placement of Metanephrocerus Aczél, 1948 (Insecta: Diptera: Pipunculidae). Arthropod Syst. Phylogeny 2014, 72, 23–36. [Google Scholar]
- Pepinelli, M.; Currie, D.C. The identity of giant black flies (Diptera: Simuliidae) in Baltic amber: Insights from large-scale photomicroscopy, micro-CT scanning and geometric morphometrics. Zool. J. Linn. Soc. 2017, 181, 846–866. [Google Scholar] [CrossRef]
- Lak, M.; Azar, D.; Nel, A.; Neraudeau, D.; Tafforeau, P. The oldest representative of the Trichomyiinae (Diptera: Psychodidae) from the Lower Cenomanian French amber studied with phase-contrast synchrotron X-ray imaging. Invertebr. Syst. 2008, 22, 471–478. [Google Scholar] [CrossRef]
- Oliva, P.; Di Trapani, V.; Arfelli, F.; Brombal, L.; Donato, S.; Golosio, B.; Longo, R.; Mettivier, G.; Rigon, L.; Taibi, A. Experimental optimization of the energy for breast-CT with synchrotron radiation. Sci. Rep. 2020, 10, 17430. [Google Scholar] [CrossRef]
- Rigon, L. x-Ray Imaging with Coherent Sources. Compr. Biomed. Phys. 2014, 4, 193–220. [Google Scholar]
- Vommaro, M.L.; Donato, S.; Giglio, A. Virtual sections and 3D reconstructions of female reproductive system in a carabid beetle using synchrotron X-ray phase-contrast microtomography. Zool. Anz. 2022, 298, 123–130. [Google Scholar] [CrossRef]
- Westneat, M.W.; Betz, O.; Blob, R.W.; Fezzaa, K.; Cooper, W.J.; Lee, W.-K. Tracheal respiration in insects visualized with synchrotron X-ray imaging. Science 2003, 299, 558–560. [Google Scholar] [CrossRef]
- Socha, J.J.; Förster, T.D.; Greenlee, K.J. Issues of convection in insect respiration: Insights from synchrotron X-ray imaging and beyond. Respir. Physiol. Neurobiol. 2010, 173, S65–S73. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Tafforeau, P. Virtual dissection using phase-contrast X-ray synchrotron microtomography: Reducing the gap between fossils and extant species. Syst. Entomol. 2011, 36, 573–580. [Google Scholar] [CrossRef]
- Klowden, M.J. Physiological Systems in Insects; Academic Press: Cambridge, MA, USA, 2013; ISBN 0124159702. [Google Scholar]
- Chapman, R.F. The Insects: Structure and Function; Cambridge University Press: Cambridge, MA, USA, 2012; ISBN 1107310458. [Google Scholar]
- Warrant, E.J.; McIntyre, P.D. Arthropod eye design and the physical limits to spatial resolving power. Prog. Neurobiol. 1993, 40, 413–461. [Google Scholar] [CrossRef]
- Rutowski, R.L. Variation of eye size in butterflies: Inter-and intraspecific patterns. J. Zool. 2000, 252, 187–195. [Google Scholar] [CrossRef]
- Bauer, T.; Desender, K.; Morwinsky, T.; Betz, O. Eye morphology reflects habitat demands in three closely related ground beetle species (Coleoptera: Carabidae). J. Zool. 1998, 245, 467–472. [Google Scholar] [CrossRef]
- Bauer, T.; Kredler, M. Morphology of the compound eyes as an indicator of life-style in carabid beetles. Can. J. Zool. 1993, 71, 799–810. [Google Scholar] [CrossRef]
- Talarico, F.; Brandmayr, P.; Giglio, A.; Massolo, A.; Brandmayr, T.Z. Morphometry of eyes, antennae and wings in three species of Siagona (Coleoptera, Carabidae). ZooKeys 2011, 100, 203–214. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B. Compound eyes of insects and crustaceans: Some examples that show there is still a lot of work left to be done. Insect Sci. 2015, 22, 461–481. [Google Scholar] [CrossRef]
- Bauer, T. Different adaptation to visual hunting in three ground beetle species of the same genus. J. Insect Physiol. 1985, 31, 593–601. [Google Scholar] [CrossRef]
- Floreano, D.; Zufferey, J.-C. Insect vision: A few tricks to regulate flight altitude. Curr. Biol. 2010, 20, R847–R849. [Google Scholar] [CrossRef]
- Srinivasan, M.V.; Poteser, M.; Kral, K. Motion detection in insect orientation and navigation. Vis. Res. 1999, 39, 2749–2766. [Google Scholar] [CrossRef]
- Schnaitmann, C.; Pagni, M.; Reiff, D.F. Color vision in insects: Insights from Drosophila. J. Comp. Physiol. A 2020, 206, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Guo, W.; Huang, S.; Luo, X.; Tian, M.; Liu, W. Morphological adaptation of cave-dwelling ground beetles in china revealed by geometric morphometry (Coleoptera, Carabidae, Trechini). Insects 2021, 12, 1002. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M. Biological clocks and visual systems in cave-adapted animals at the dawn of speleogenomics. Integr. Comp. Biol. 2013, 53, 50–67. [Google Scholar] [CrossRef] [PubMed]
- Escalona, H.E.; Oberprieler, R.G. Undarobius, a new genus of cavernicolous weevils (Curculionidae: Entiminae: Leptopiini) from the Undara Lava Caves in north-eastern Australia, with an overview of anophthalmic and microphthalmic Australian Curculionidae. Zootaxa 2021, 5023, 207–222. [Google Scholar] [CrossRef]
- Chen, Q.-X.; Hua, B.-Z. Ultrastructure and morphology of compound eyes of the scorpionfly Panorpa dubia (Insecta: Mecoptera: Panorpidae). PLoS ONE 2016, 11, e0156970. [Google Scholar] [CrossRef]
- Guo, F.; Ning, S.; Feng, J.; Liu, B.; He, X. Ultrastructure and morphology of the compound eyes of the predatory bug Montandoniola moraguesi (Insecta: Hemiptera: Anthocoridae). Arthropod Struct. Dev. 2021, 61, 101030. [Google Scholar] [CrossRef]
- Borst, A. Drosophila’s view on insect vision. Curr. Biol. 2009, 19, R36–R47. [Google Scholar] [CrossRef]
- Feller, K.D.; Sharkey, C.R.; McDuffee-Altekruse, A.; Bracken-Grissom, H.D.; Lord, N.P.; Porter, M.L.; Schweikert, L.E. Surf and turf vision: Patterns and predictors of visual acuity in compound eye evolution. Arthropod Struct. Dev. 2021, 60, 101002. [Google Scholar] [CrossRef]
- Alba-Tercedor, J. Microtomographic study on the anatomy of adult male eyes of two mayfly species. Zoosymposia 2016, 11, 101–120. [Google Scholar] [CrossRef]
- Kim, B.H.; Seo, E.S.; Lim, J.H.; Lee, S.J. Synchrotron X-ray microscopic computed tomography of the pump system of a female mosquito. Microsc. Res. Tech. 2012, 75, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Ribi, W.; Senden, T.J.; Sakellariou, A.; Limaye, A.; Zhang, S. Imaging honey bee brain anatomy with micro-X-ray-computed tomography. J. Neurosci. Methods 2008, 171, 93–97. [Google Scholar] [CrossRef]
- Rother, L.; Kraft, N.; Smith, D.B.; el Jundi, B.; Gill, R.J.; Pfeiffer, K. A micro-CT-based standard brain atlas of the bumblebee. Cell Tissue Res. 2021, 386, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vega, D.; Wicklein, M.; Simonsen, T.J.; Garbout, A.; Ahmed, F.; Hall, M.J.R. Anatomical reconfiguration of the optic lobe during metamorphosis in the blow fly Calliphora vicina (Diptera: Calliphoridae) revealed by X-ray micro-computed tomography. Zool. Anz. 2021, 292, 139–149. [Google Scholar] [CrossRef]
- García, N.; Numa, C.; Bartolozzi, L.; Brustel, H.; Buse, J.; Norbiato, M.; Recalde, J.I.; Zapata, J.; Dodelin, B.; Alcázar, E.; et al. The Conservation Status and Distribution of Mediterranean Saproxylic Beetles; IUCN: Malaga, Spain, 2019; ISBN 978-2-8317-1946-7. [Google Scholar]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. A Red List of Italian Saproxylic Beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Entomol. 2015, 47, 53. [Google Scholar] [CrossRef]
- Hagstrum, D. Atlas of Stored-Product Insects and Mites; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 0128104317. [Google Scholar]
- Sunderland, K.D. Invertebrate Pest Control by Carabids. In The Agroecology of Carabid Beetles; Holland, J.M., Ed.; Intercept Limited: Andover, UK, 2002; pp. 165–214. [Google Scholar]
- Giglio, A.; Vommaro, M.L.; Gionechetti, F.; Pallavicini, A. Gut microbial community response to herbicide exposure in a ground beetle. J. Appl. Entomol. 2021, 145, 986–1000. [Google Scholar] [CrossRef]
- Giglio, A.; Giulianini, P.G.; Zetto, T.; Talarico, F. Effects of the pesticide dimethoate on a non-target generalist carabid, Pterostichus melas italicus (Dejean, 1828) (Coleoptera: Carabidae). Ital. J. Zool. 2011, 78, 471–477. [Google Scholar] [CrossRef]
- Giglio, A.; Brandmayr, P.; Pasqua, T.; Angelone, T.; Battistella, S.; Giulianini, P.G. Immune challenges trigger cellular and humoral responses in adults of Pterostichus melas italicus (Coleoptera, Carabidae). Arthropod Struct. Dev. 2015, 44, 209–217. [Google Scholar] [CrossRef]
- Trautner, J.; Geigenmüller, K. Tiger Beetles, Ground Beetles. Illustrated Key to the Cicindelidae and Carabidae of Europe; TRIOPS Verlag: Aichtal, Germany, 1987; ISBN 3924333041. [Google Scholar]
- Milutinović, B.; Stolpe, C.; Peuβ, R.; Armitage, S.A.O.; Kurtz, J. The red flour beetle as a model for bacterial oral infections. PLoS ONE 2013, 8, e64638. [Google Scholar] [CrossRef]
- Tromba, G.; Longo, R.; Abrami, A.; Arfelli, F.; Astolfo, A.; Bregant, P.; Brun, F.; Casarin, K.; Chenda, V.; Dreossi, D. The SYRMEP Beamline of Elettra: Clinical Mammography and Bio-medical Applications. In AIP Conference Proceedings; American Institute of Physics: College Park, MD, USA, 2010; Volume 1266, pp. 18–23. [Google Scholar]
- Giuliani, A.; Mazzoni, S.; Mele, L.; Liccardo, D.; Tromba, G.; Langer, M. Synchrotron phase tomography: An emerging imaging method for microvessel detection in engineered bone of craniofacial districts. Front. Physiol. 2017, 8, 769. [Google Scholar] [CrossRef]
- Brombal, L. X-ray Phase-Contrast Tomography: Underlying Physics and Developments for Breast Imaging; Springer Nature: Berlin, Germany, 2020; ISSN 2190-5052. [Google Scholar]
- Donato, S.; Arana Peña, L.M.; Bonazza, D.; Formoso, V.; Longo, R.; Tromba, G.; Brombal, L. Optimization of pixel size and propagation distance in X-ray phase-contrast virtual histology. J. Instrum. 2022; in press. [Google Scholar]
- Brun, F.; Massimi, L.; Fratini, M.; Dreossi, D.; Billé, F.; Accardo, A.; Pugliese, R.; Cedola, A. SYRMEP Tomo Project: A graphical user interface for customizing CT reconstruction workflows. Adv. Struct. Chem. Imaging 2017, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Paganin, D.; Mayo, S.C.; Gureyev, T.E.; Miller, P.R.; Wilkins, S.W. Simultaneous phase and amplitude extraction from a single defocused image of a homogeneous object. J. Microsc. 2002, 206, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Gureyev, T.E.; Nesterets, Y.I.; Kozlov, A.; Paganin, D.M.; Quiney, H.M. On the “unreasonable” effectiveness of transport of intensity imaging and optical deconvolution. JOSA A 2017, 34, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Brombal, L.; Donato, S.; Dreossi, D.; Arfelli, F.; Bonazza, D.; Contillo, A.; Delogu, P.; Di Trapani, V.; Golosio, B.; Mettivier, G. Phase-contrast breast CT: The effect of propagation distance. Phys. Med. Biol. 2018, 63, 24NT03. [Google Scholar] [CrossRef] [PubMed]
- Piai, A.; Contillo, A.; Arfelli, F.; Bonazza, D.; Brombal, L.; Cova, M.A.; Delogu, P.; Di Trapani, V.; Donato, S.; Golosio, B. Quantitative characterization of breast tissues with dedicated CT imaging. Phys. Med. Biol. 2019, 64, 155011. [Google Scholar] [CrossRef]
- Limaye, A. Drishti: A volume exploration and presentation tool. In Proceedings of the Developments in X-ray Tomography VIII, San Diego, CA, USA, 12–16 August 2012; Volume 8506, p. 85060X. [Google Scholar]
- Rasband, W.S. Imagej; US National Institutes of Health: Bethesda, Maryland, USA, 2011. Available online: http://imagej.nih.gov/ij/ (accessed on 11 April 2022).
- Friedrich, F.; Matsumura, Y.; Pohl, H.; Bai, M.; Hörnschemeyer, T.; Beutel, R.G. Insect morphology in the age of phylogenomics: Innovative techniques and its future role in systematics. Entomol. Sci. 2014, 17, 1–24. [Google Scholar] [CrossRef]
- Wipfler, B.; Pohl, H.; Yavorskaya, M.I.; Beutel, R.G. A review of methods for analysing insect structures—the role of morphology in the age of phylogenomics. Curr. Opin. Insect Sci. 2016, 18, 60–68. [Google Scholar] [CrossRef]
- Ploomi, A.; Merivee, E.; Rahi, M.; Bresciani, J.; Ravn, H.P.; Luik, A.; Sammelselg, V. Antennal sensilla in ground beetles (Coleoptera, Carabidae). Agron. Res. 2003, 1, 221–228. [Google Scholar]
- Altner, H.; Loftus, R. Ultrastructure and Function of Insect Thermo- And Hygroreceptors. Annu. Rev. Entomol. 1985, 30, 273–295. [Google Scholar] [CrossRef]
- Giglio, A.; Brandmayr, P.; Ferrero, E.A.; Giulianini, P.G.; Perrotta, E.; Talarico, F.F.; Zetto Brandmayr, T. Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922) (Coleoptera, Carabidae) and possible correlations between size and shape and the larval feeding habits. Zool. Anz. 2008, 247, 209–221. [Google Scholar] [CrossRef]
- Giglio, A.; Perrotta, E.; Talarico, F.; Zetto Brandmayr, T.; Ferrero, E.A. Sensilla on maxillary and labial palps in a helicophagous ground beetle larva (Coleoptera, Carabidae). Acta Zool. 2013, 94, 324–330. [Google Scholar] [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S. Aquatic insect sensilla: Morphology and function. Aquat. Insects 2019, 139–166. [Google Scholar]
- Faucheux, M.J.; Németh, T.; Hoffmannova, J.; Kundrata, R. Scanning electron microscopy reveals the antennal micromorphology of Lamprodila (Palmar) festiva (Coleoptera: Buprestidae), an invasive pest of ornamental cupressaceae in western palaearctic. Biology 2020, 9, 375. [Google Scholar] [CrossRef] [PubMed]
- Makarova, A.A.; Meyer-Rochow, V.B.; Polilov, A.A. Morphology and scaling of compound eyes in the smallest beetles (Coleoptera: Ptiliidae). Arthropod Struct. Dev. 2019, 48, 83–97. [Google Scholar] [CrossRef]
- Harrison, F.W. Microscopic Anatomy of Invertebrates; Wiley-Liss: Wilmington, DE, USA, 1991; ISBN 0471568422. [Google Scholar]
- Sombke, A.; Lipke, E.; Michalik, P.; Uhl, G.; Harzsch, S. Potential and limitations of X-ray micro-computed tomography in arthropod neuroanatomy: A methodological and comparative survey. J. Comp. Neurol. 2015, 523, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Horridge, G. The compound eye of insects. Sci. Am. 1977, 237, 108–121. [Google Scholar] [CrossRef]
- Giglio, A.; Mazzei, A.; Vommaro, M.L.; Brandmayr, P. Antennal sensilla in an anophthalmic wood-dwelling species, Clinidium canaliculatum, Costa 1839 (Coleoptera, Rhysodidae). Microsc. Res. Tech. 2022, 85, 1005–1015. [Google Scholar] [CrossRef]
- Mazzei, A.; Audisio, P.; Taglianti, A.V.; Brandmayr, P. Geographical distribution and conservation status of the threatened saproxylic beetles Rhysodes sulcatus (Fabricius, 1787), Clinidium canaliculatum (O.G. Costa, 1839) and Omoglymmius germari (Ganglbauer, 1891) in Italy (Coleoptera: Rhysodidae). Fragm. Entomol. 2019, 51, 89–96. [Google Scholar] [CrossRef]
- Land, M.F. Visual acuity in insects. Annu. Rev. Entomol. 1997, 42, 147–177. [Google Scholar] [CrossRef]
- Vácha, M.; Půžová, T.; Drštková, D. Effect of light wavelength spectrum on magnetic compass orientation in Tenebrio molitor. J. Comp. Physiol. A 2008, 194, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Yinon, U. The visual mechanisms of Tenebrio molitor: Some aspects of the spectral response. J. Exp. Biol. 1970, 53, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Yinon, U. The Visual Mechanisms of Tenebrio molitor: Changes in the Electro-Retinogram as Function of the Stimulus Duration. J. Exp. Biol. 1971, 54, 737–744. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
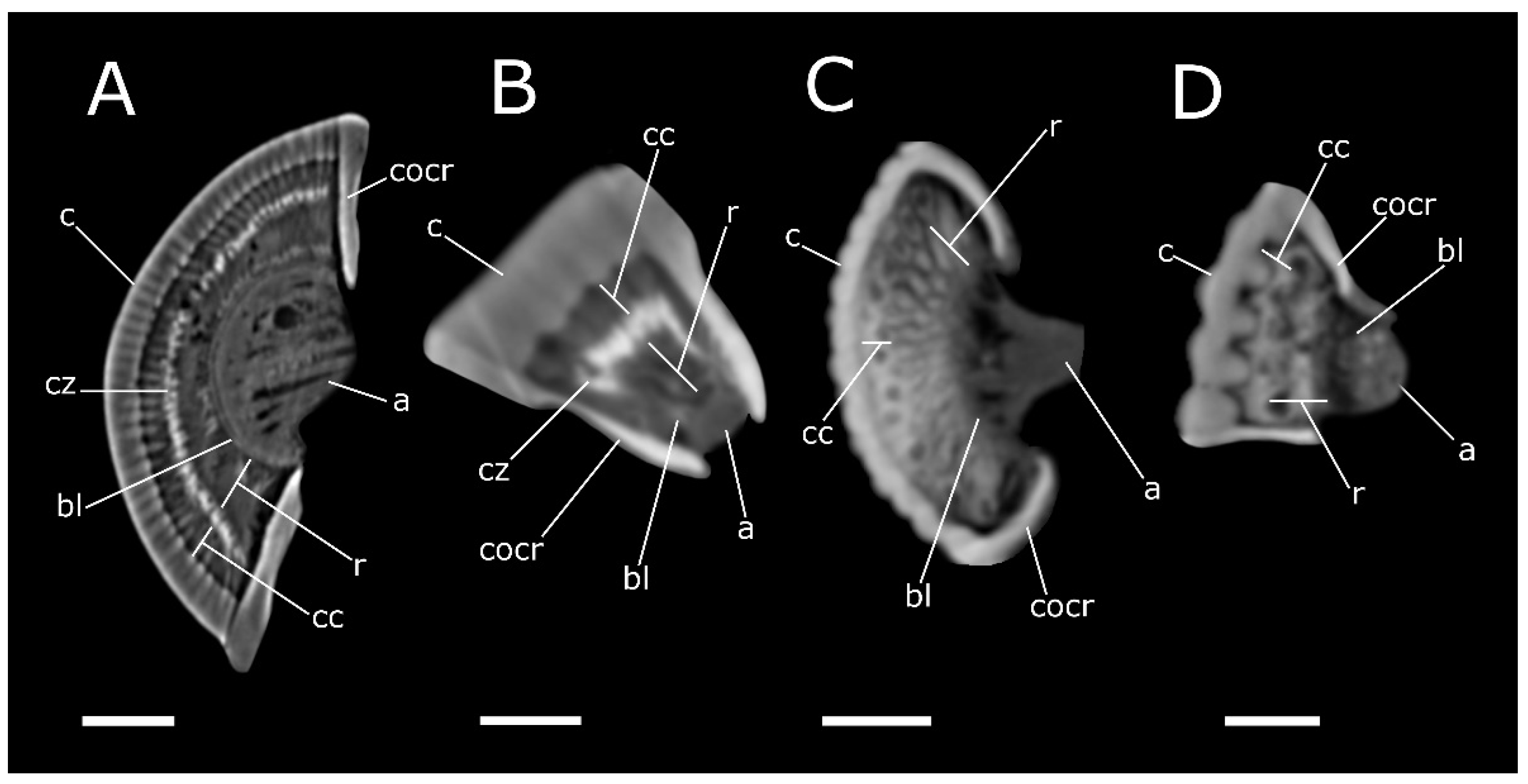{kind=link}
| Species | N. of Ommatidia | Corneal Facet Surface Area (µm2) | Total Surface of Facets a (µm2) | Cornea Surface b (µm2) | Facet Density c | Corneal Facet Lens Thickness (µm) | Interocular Distance (mm) | Head Size d (mm) |
|---|---|---|---|---|---|---|---|---|
| Tribolium castaneum | 92 | (1.02 ± 0.14) × 103 | (9.40 ± 1.31) × 104 | 9.0 × 104 | 1022 | 22.8 ± 0.97 | 0.43 | 0.7–0.7 |
| Tenebrio molitor | 440 | (1.97 ± 0.26) × 103 | (8.67 ± 1.13) × 105 | 8.2 × 105 | 536.6 | 38.7 ± 1.03 | 1.85 | 2.68–2.8 |
| Pterostichus melas italicus | 1380 | (0.50 ± 0.07) × 103 | (6.96 ± 0.99) × 105 | 6.9 × 105 | 2000 | 64.9 ± 1.68 | 2.73 | 3.39–3.45 |
| Clinidium canaliculatum | 70 | (0.29 ± 0.06) × 103 | (2.09 ± 0.56) × 104 | 4.5 × 104 | 1555 | 50.94 ± 0.63 | 0.79 | 0.97–1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giglio, A.; Vommaro, M.L.; Agostino, R.G.; Lo, L.K.; Donato, S. Exploring Compound Eyes in Adults of Four Coleopteran Species Using Synchrotron X-ray Phase-Contrast Microtomography (SR-PhC Micro-CT). Life 2022, 12, 741. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050741
Giglio A, Vommaro ML, Agostino RG, Lo LK, Donato S. Exploring Compound Eyes in Adults of Four Coleopteran Species Using Synchrotron X-ray Phase-Contrast Microtomography (SR-PhC Micro-CT). Life. 2022; 12(5):741. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050741
Chicago/Turabian StyleGiglio, Anita, Maria Luigia Vommaro, Raffaele Giuseppe Agostino, Lai Ka Lo, and Sandro Donato. 2022. "Exploring Compound Eyes in Adults of Four Coleopteran Species Using Synchrotron X-ray Phase-Contrast Microtomography (SR-PhC Micro-CT)" Life 12, no. 5: 741. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050741