
Functional Properties of Cancer Epithelium and Stroma-Derived Exosomes in Head and Neck Squamous Cell Carcinoma


1
Department of Oral Pathology, College of Stomatology, Ningxia Medical University, South Sheng Li Street 804, Yinchuan 750004, China
2
Key Laboratory of Stomatology of Fujian Province, School and Hospital of Stomatology, Fujian Medical University, Yang Qiao Middle Road 246, Fuzhou 350004, China
3
State Key Laboratory of Oral Diseases, West China College of Stomatology, Sichuan University, South Renmin Road, Sec. 3, No. 14, Chengdu 610041, China
4
College of Public Health and Management, Ningxia Medical University, South Sheng Li Street 1160, Yinchuan 750004, China
5
State Key Laboratory of Oral Diseases, National Center of Stomatology, National Clinical Research Center for Oral Diseases, Frontier Innovation Center for Dental Medicine Plus, West China Hospital of Stomatology, Sichuan University, South Renmin Road, Sec. 3, No. 14, Chengdu 610041, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Life 2022, 12(5), 757; https://0-doi-org.brum.beds.ac.uk/10.3390/life12050757
Submission received: 8 May 2022
/
Accepted: 13 May 2022
/
Published: 20 May 2022
(This article belongs to the Special Issue Exosomes: Biogenesis, Biologic Function and Clinical Potential)
Abstract
:Stroma–cancer cell crosstalk involves a complex signaling network that contributes to tumor progression, including carcinogenesis, angiogenesis, migration, invasion, and therapy resistance in cancers. Exosomes, as extracellular membranous nanovesicles released by almost all types of cells, including tumor cells and stromal cells, play a critical role in signal delivery and material communication, in which the characteristics of their parent cells are reflected. The tumor or stroma-derived exosomes mediate cell–cell communication in the tumor microenvironment by transporting DNA, RNA, proteins, lipids, and metabolites. Recent studies on head and neck squamous cell carcinoma (HNSCC) have demonstrated that tumor-derived exosomes support various tumor biological behaviors, whereas the functional roles of stroma-derived exosomes remain largely unknown. Although these exosomes are emerging as promising targets in early diagnosis, prognostic prediction, and pharmaceutical carriers for antitumor therapy, there are still multiple hurdles to be overcome before they can be used in clinical applications. Herein, we systematically summarize the promotive roles of the epithelium and stroma-derived exosomes in HNSCC and highlight the potential clinical applications of exosomes in the treatment of HNSCC.
1. Introduction
Head and neck cancer, including cancers in the oral cavity, oropharynx, hypopharynx, nasopharynx, and larynx, is the sixth most common type of cancer, with an estimated 600,000 new cases occurring globally each year [1,2]. Among these, more than 90% are head and neck squamous cell carcinoma (HNSCC) based on their histopathological type [3]. Most HNSCCs arise from stratified epithelia of the skin, oral cavity, and pharynx and in the larynx. Although the multidisciplinary approach using surgical resection with or without radio- and chemotherapy is still the gold standard for treatment, the five-year overall survival rate of HNSCC patients was reported to be merely 60% [4], indicating that novel targeting approaches for improving patient survival and quality of life with HNSCC are required. To date, most of the previous preclinical trials based on altered gene expression in cancer epithelial cells have failed, and one of the potential reasons is the limited understanding of the crosstalk between the cancer epithelium and the tumor microenvironment (TME) in therapeutic resistance. The TME, which is comprised of cancer-associated fibroblasts (CAFs), immune cells, and other stromal cells, plays an important role in tumor progression of HNSCC [5,6]. Since cancer stromal cells play promotive and supportive roles in tumor progression, traditional therapeutic paradigms are often insufficient to exterminate tumor cells. Thus, as we previously concluded [7], targeting the crosstalk between stromal cells and cancer epithelial cells is a promising avenue in anticancer therapy.
Exosomes are small extracellular vehicles (EVs) of endosomal origin with a diameter ranging from 40 to 160 nm that widely engage in the crosstalk between cancer cells and stromal cells [8]. The cargo of exosomes is abundant in proteins, lipids, RNA and DNA, and other compounds. Frequent exosome-guided cellular and molecular communication is reported to promote information transmission in cancers [9,10,11]. In the TME, stromal cells are connective cells including CAFs, mesenchymal stem cells (MSCs), and endothelial cells (ECs) that maintain the structural framework in tumor, and another class of stromal cells that maintains cancer homeostasis is immune cells including dendritic cells, B cells, T cells, macrophages, and natural killer (NK) cells [12]. These stromal cells produce various exosomes involved in tumorigenesis, tumor growth, cancer recurrence, and metastasis due to their versatile roles in promoting cancer cell proliferation, migration, invasion, and therapeutic resistance [13,14,15,16,17]. Therefore, exosomes have become candidates for noninvasive liquid diagnosis and monitoring targeting cancer treatment [18,19]. Herein, we summarize the roles of cancer epithelium/stroma-derived exosomes in mediating tumor progression and their diagnostic and therapeutic applications in HNSCC, which establishes a foundation for advances in exosome-directed early diagnosis, prognostic prediction, and antitumor therapy in HNSCC.
2. Biological Characteristics of Exosomes in HNSCC
Exosomes are nanosized vesicles, and those that are secreted by tumor cells are called tumor-derived exosomes (TDEs). The total amount of exosomes released by tumor cells is about 10-fold more than from normal cells [20]. Typically, TDE components include proteins, lipids, DNA, multiple RNA species, and other substances that can be taken up by recipient cells and have the function of regulating receptor cell gene expression, tumorigenesis, angiogenesis, TME reprogramming, and immune tolerance, thereby promoting metastasis and recurrence and therapy resistance [21,22,23]. Recently, various methods, including centrifugation, size exclusion chromatography (SEC), polymer-based precipitation, (immuno-)affinity capture, and microfluidic approaches, have been developed to isolate exosomes. TDEs and stroma-derived exosomes are a highly heterogeneous population of membrane vesicles, varying according to the type of cancer and even stage for the same tumor type, characterized by differences in size, secretion level, morphology, cargo, recipient cells, functional properties, and potential mechanisms (Table 1). TDEs range from 30 to 200 nm in size, whereas stroma-derived exosomes are 40–150 nm. Interestingly, the metastatic OSCC cells secrete larger exosomes than those derived from the parental OSCC cells [24], suggesting that larger size exosomes probably transport more molecular contents for metastasis at the late stage of tumors. Transmission electron microscopy (TEM) is commonly used for exosome visualization. In TEM images, the majority of exosomes secreted by tumor cells and stroma cells exhibit a round or spherical shape. For the cargo in exosomes, protein and miRNA are the two major types of content that have been considerably explored in the context of tumor progression in HNSCC. Previous studies showed that exosomes contain a vast array of proteins, including membrane trafficking and cytoskeletal proteins, signal transducers, and heat-shock proteins, in addition to a high abundance of exosomal microRNAs. The recipient cells of TDEs or stroma-derived exosomes suggest that the exosomes transport cargo from one cell to another in the TME and, thus, can initiate signaling responses for different biological behaviors in HNSCC.
3. The Effects of Cancer Epithelium Derived Exosomes on Tumor Progression in HNSCC
3.1. Role of Cancer Epithelium Derived Exosomes in Tumorigenesis
Tumorigenesis is the process in which normal epithelial cells transform into malignant cancer cells characterized by properties of high proliferation, evasion of apoptosis, immunosurveillance evasion, malignant invasion, and metastasis. A variety of potential mechanisms related to carcinogenesis in HNSCC and TDEs are involved in this process. For instance, Ono et al. provided in vitro evidence showing that metastatic oral cancer-derived exosomes play a role in promoting the epithelial–mesenchymal transition (EMT) process in normal epithelial cells, which is required for tumor initiation [27]. Mechanistically, they found that HSP90-enriched oral metastatic TDEs induce tumor-associated macrophage (TAM) polarization in M2 macrophages, supporting cancer progression, with simultaneous CDC37/HSP90α/β depletion attenuating the metastatic TDEs-driven tumor-initiating activity in oral cancer [27] (Figure 1A). Similarly, Pang et al. found that exosomal CMTM6 secreted by tumor cells induced polarization of M2-like macrophages through the ERK1/2 signaling pathway, thereby promoting malignant progression and participating in mediating crosstalk between cancer cells and macrophages in oral squamous cell carcinoma (OSCC) [42] (Figure 1A). Of note, the CMTM6 levels were positively associated with M2-like macrophage infiltration in the spontaneous OSCC induced by 4-nitroquinoline 1-oxide (4NQO) [42]. In another 4NQO model of carcinogenesis in OSCC, Beatrice et al. showed that application of TDEs isolated from murine or human OSCC cells were sufficient to induce carcinogenesis in mice bearing premalignant lesions by reducing T cell infiltration [33]. Ye et al. have shown that NPC-derived exogenous miRNAs affects T cell proliferation, differentiation and cytokine secretion, which may be related to accumulation of exogenous miRNAs targeting MAPK1 and JAK/STAT pathways [30], indicating the bidirectional effects of exosomes between cancer cells and stromal cells in NPC. Altogether, this evidence suggests that tumor-derived exosomes mainly mediate tumor initiation through crosstalk between cancer cells and immune cells, especially affecting the feedback regulation of TAM polarization and the immunosuppressive effects.
3.2. Role of Cancer Epithelium Derived Exosomes in Angiogenesis
Angiogenesis is the formation of new blood vessels from the existing vasculature and it plays a central role in providing oxygen and nutrients to tumors [43]. It has been reported that exosomes are enriched in angiogenesis-related factors, including angiogenic proteins, miRNAs, and mRNAs [44], and accumulating evidence suggests that TDEs promote angiogenesis during tumor progression. Vascular endothelial growth factor (VEGF) is one of the most important angiogenesis-related factors in cancer. For instance, in HNSCC, TDEs containing oncogenic EGFR are taken up by endothelial cells both in vitro and in vivo, activating the autocrine VEGF/VEGFR-2 pathway to induce angiogenesis [45] (Figure 1B). Wang et al. showed that a strong pro-angiogenic effect of TDEs shuttling angiogenin, bFGF, and VEGF could stimulate endothelial cells for angiogenesis by activated STAT3, c-Jun N-terminal kinase (JNK), and p53 signal pathways in vivo [46]. Membrane phosphatidylserine mediates the exchange of TDEs by blocking membrane phosphatidylserine with diannexin as demonstrated in mouse models, and tumors therefore exhibited lower microvascular densities than the controls [45]; Bao et al. also found a relationship between microvascular density and prognosis in nasopharyngeal carcinoma (NPC), indicating that TDE-derived miR-23a promotes angiogenesis by targeting TSGA10, and the high levels of miR-23a in metastatic and premetastatic tissues suggests a close relationships between overall survival time of NPC patients, play a promotive role in angiogenesis [47] (Figure 1B).
In another NPC study, TDE-derived miR-9 inhibited endothelial tube formation by targeting MDK and regulating PDK/AKT signaling pathways [48] (Figure 1B), suggesting that different types of miRNAs in TDEs exert different functions in angiogenesis. Interestingly, in contrast to its role in NPC, Zhang et al. found that TDE-derived miR-9 promotes angiogenesis by activating the JAK-STAT pathway in endothelial cells for various types of cancers, including non-small cell lung cancer, melanoma, pancreatic cancer, and colorectal cancer [49] (Figure 1B). Thus, the effect of TDE-derived miR-9 in angiogenesis appears to be tumor/tissue-specific, and any therapeutic strategies designed to target TDE-derived miR-9 should focus on, or be aware of, its paradoxical roles in the mechanisms of angiogenesis depending on the type of cancer.
3.3. Role of Cancer Epithelium Derived Exosomes in Recurrence and Metastasis
In OSCC, about one-third of patients develop local recurrence or distant metastasis after routine treatment [50]. TDEs are reported to play an important role in promoting recurrence and metastasis in HNSCC. For instance, Sakha et al. showed that miR-1246 derived by human oral cancer cells could improve cells’ motility and invasive ability by directly targeting DENN/MADD domain-containing 2D (DENND2D) to activate the ERK and AKT signaling pathways, thereby increasing the incidence of cancer metastasis [34] (Figure 1C). Similarly, Cai et al. found that OSCC-derived exosomes promote M2 macrophage polarization through a mechanism involving the SOCS1/STAT6 signaling pathway in macrophages activated by miR-29a-3p, promoting the invasion of OSCC cells [51] (Figure 1). In esophageal cancer, Liao et al. found that exosomal miR-21 targets programmed cell death 4 (PDCD4) and then activates the downstream c-Jun N-terminal kinase (JNK) cascade, subsequently promoting recurrence and distant metastasis [52] (Figure 1C). Mechanistically, exosome shuttling miR-21 repressed PDCD4 expression by binding to 3′-UTR and increased the expression of MMP-2 and MMP-9 [52]. It suggests that exosome derived miR-21 correlates closely with esophageal cancer metastasis and has the potential to be a biomarker for blocking metastasis. Notably, in HNSCC, one of the most important prognostic determinants is the presence of lymph node metastasis, and lymphatic metastatic spread is correlated with a significant decrease in the patient survival rate. Mouse- and human-derived TDEs may carry microRNA species that change the genetic and protein profiles, thus favoring metastasis formation [53]. In support of this notion, Li et al. found that a hypoxic microenvironment can stimulate OSCC to generate miR-21-rich exosomes in a HIF-1α- and HIF-2α-dependent manner, thereby promoting lymph node metastasis both in vivo and in vitro [36]. Additionally, by HIF-1α or HIF-2α knockdown, exosomal miR-21 levels were significantly decreased in hypoxic conditions [36], indicating that the transcription of exomal miR-21 might be mediated by HIF-1α/HIF-2α. In another study, Ono et al. provided data demonstrating an abundant secretion of HSP90-enriched TDEs in OSCC with lymph node metastasis [24]. Interestingly, by using small interfering RNA to knockdown either HSP90α or HSP90β, there was no apparent effect on the survival of the metastatic OSCC cells; however, the simultaneous knockdown of HSP90α and HSP90β significantly reduced their survival [24], demonstrating that TDE-enclosed HSP90 may be a potential prognostic biomarker and therapeutic target in OSCC with lymph node metastasis. In summary, this evidence suggests that TDEs play a critical role in the recurrence and metastasis of HNSCC by enhancing cancer cell invasion, migration, and survival.
3.4. Role of Cancer Epithelium Derived Exosomes in Therapeutic Resistance
Clinically, for those HNSCC patients not undergoing surgical therapy, the addition of chemotherapy, radiation therapy, and/or immunotherapy is necessary for improving their prognosis. Previous studies have suggested that exosomes can induce therapeutic resistance by transferring miRNAs, mRNAs, DNA, proteins, and purine metabolites [54,55]. For instance, Liu et al. demonstrated that TDEs from cisplatin-resistant OSCC cells convey miR-21 to parental cells and induce cisplatin resistance by targeting phosphatase and tensin homolog (PTEN) and PDCD4; the chemoresistance of parental OSCC cells can be attenuated by the inhibitor (GW4869), which suppresses exosome release [37] (Figure 1D), thus suggesting that TDEs may play a role in promoting chemoresistance. For radiotherapy, it has been reported that miR-20a-5p secreted by radiation-resistant nasopharyngeal carcinoma cells is transferred to adjacent cells through exosomes, which inhibits Rab27B by targeting Rab27B3′-UTR (a target of miR-20a-5p), leading to radiation tolerance [56] (Figure 1D). Of note, chemoradiation therapy is already one of the major treatments for HNSCC, though it results in more systemic adverse effects than radiotherapy or chemotherapy alone. Furthermore, Beck et al. found that various exosomes secreted by hypoxic tumor tissues after radiotherapy increase invasion and metastasis in HNSCC, promoting drug resistance during radiotherapy [57] and indicating the need to suppress chemotherapy resistance induced by TDEs during radiotherapy. Additionally, since HNSCC cells escape immunosurveillance for recurrence or metastasis, an in-depth understanding of the potential resistance mechanisms and design of immunotherapeutic approaches is urgently needed. Targeting TDEs may be a promising candidate as another immunotherapeutic avenue. Ludwig et al. performed a clinical study showing that purine metabolites, including adenosine and 5′-GMP in plasma-derived exosomes, increased in patients with early-stage HNSCC with no lymph node metastasis [28], suggesting that immunosuppressive adenosine in the exosomes may play a promotive role in early tumor progression. Interestingly, by comparing the exosome profiles in patients with or without lymph node metastasis, they found that levels of most purine metabolites including adenosine and its derivatives were higher in patients without lymph node metastasis [28]. An alternative hypothesis, we suppose, is that purine metabolites might be mainly used for cell proliferation in the primary site at an early stage for tumor growth while fewer purine metabolites are packaged into exosomes in the circulation for metastatic tumor cells. Therefore, exosome-enclosed purine metabolites may be valuable candidates for a targeted immunotherapeutic approach and sensitive biomarkers for prognostic prediction in HNSCC.
4. Cellular Origins of Cancer Stroma Derived Exosomes and Their Roles in HNSCC Tumor Progression
4.1. Cancer-Associated Fibroblast-Derived Exosomes in HNSCC
We previously systematically summarized the evidence for CAFs, the major cellular component of stromal cells in the TME, which indicated they are critical for cancer occurrence and progression due to their versatile roles [7]. Studies have shown that CAF-derived exosomes shuttle cargo, such as proteins and miRNAs, to perform their various oncogenic functions. For instance, in their proteomic analysis in which they identified 4247 proteins, Principe et al. found that microfibrillar-associated protein 5 (MFAP5) in CAF-derived exosomes promotes proliferation and metastasis of OSCC by activating the MAPK and AKT signaling pathways [58] (Figure 2A). In another study, Li et al. found that the reduced expression of miR-34a-5p in CAF-derived exosomes promotes the proliferation and metastasis of OSCC by binding the downstream target of AXL; furthermore, the miR-34a-5p/AXL axis induces the epithelial–mesenchymal transition (EMT) process for cancer metastasis via the AKT/GSK-3β/β-catenin signal cascade [40] (Figure 2A). Similarly, in OSCC, Sun et al. showed that miR-382-5p was overexpressed in CAFs compared with the fibroblasts from adjacent normal tissue and that CAF-derived exosomal miR-382-5p promotes OSCC cell migration and invasion [59] (Figure 2A). Studies on CAF-mediated therapy resistance have recently reported the use of CAF targeting as an approach for reversing tumor chemoresistance in CAF-mediated anticancer therapy [7,60]. For example, Qin et al. found that exosomal miR-196a secreted by CAFs is resistant to cisplatin and thus enhances cisplatin resistance, and miR-196a could be transferred from CAFs to tumor cells through exosomes, subsequently promoting the survival and proliferation of HNSCC cells [38]. Interestingly, Yeung and colleagues demonstrated that transfer of CAF-derived exosomal miR21 to cancer cells increases chemoresistance to paclitaxel in ovarian cancer cells [61] (Figure 2A). Altogether, the aforementioned evidence suggests that targeting of the miRNAs shuttled in CAF-derived exosomes may be a promising strategy in cancer treatment that addresses issues of resistance, and any targeted strategy based on this strategy should consider tumor/tissue specificity when attempting to optimize therapeutic efficacy.
4.2. Tumor-Associated Macrophage-Derived Exosomes in HNSCC
Induced by chemokines and cytokines, among others, TAM polarization can be guided toward the M1 (with pro-inflammatory and antitumor effects) and M2 (with anti-inflammatory and pro-tumor functions) phenotypes to affect tumor growth, migration, and therapeutic resistance in an exosome-dependent manner. For instance, Lee et al. found that macrophages generated from THP-1 induced by phorbol 12-myristate 13-acetate (PMA) are rich in disintegrin and metalloproteinase 15 (ADAM15) and result in delayed tumor growth in vivo, and the tumor volume was also diminished in mice treated with ADAM15-blocked exosomes [62] (Figure 2B), suggesting that ADAM15 exosomes derived from macrophages play a suppressive role in tumor growth and stimulation by PMA for these kinds of macrophages may result in polarization into the M1 phenotype for antitumor effects. Interestingly, Xiao et al. demonstrated that THP-1 treated with exosomes from OSCC cells exhibited an M1-like but not an M2-like phenotype in promoting migration [32], supporting the notion that TAMs are highly heterogeneous stromal cells whose crosstalk with cancer cells is mediated by multiple signals, a point that should be considered in antitumor therapy in OSCC based on the conversion of TAMs from the M2 to M1 phenotype. Of note, the exosomes secreted by THP-1 derived macrophages depends not only on PMA simulation but also the surrounding status including the lipopolysaccharide (LPS) [63] and IL-4 [64,65]. This suggests that the exosomes from THP-1 derived macrophages are highly sensitive to the exogenous stimulus, in part because the exosomes in different cancers exhibit heterogeneity and specificity. Most importantly, macrophage-derived exosomes might reduce the sensitivity of OSCC cells to chemother- apeutic drugs. For example, Tomita et al. found that exosomes derived from the THP-1 and PHM activated Akt/glycogen synthase kinase 3 β signaling pathways to promote proliferation by reducing the proliferation inhibition of 5- FU and CDDP, and that inhibitors of PI3K (LY294002) and Akt (mk-226) may attenuate the decreased exosome-induced chemotherapy sensitivity in OSCC [16]. Similarly, in gastric cancer, TAM-derived-exosome-enclosed miR-21 diminishes chemotherapy sensitivity [14] (Figure 2B). In summary, this evidence indicates that the use of TAM-derived exosomes may be considered a therapeutic approach for anticancer therapy, especially for improving chemosensitivity in HNSCC treatment.
4.3. Exosomes Derived from T Cells, B Cells, and NK Cells in HNSCC
Although T cell-derived exosomes have been widely explored in terms of immunity and antitumor effects [66,67,68,69,70], few studies reported on the relation to HNSCC [17]. In esophageal SCC, Min et al. reported that T cell-derived exosomes could promote the metastasis of esophageal cancer cells by inducing EMT through upregulation of β-catenin and the NF-κB/snail pathway [17] (Figure 2C). For B cell-derived exosomes, Saunderson et al. suggested that B cell vesicles were involved in inducing effective cytotoxic T lymphocyte (CTL) responses in DH LMP2A mice [71]. Of note, human B cell-derived lymphoblastic-like cell line (LCL)-derived MHCII+FasL+ exosomes could induce CD4+ T cell apoptosis in an antigen-specific dependent manner and this process might also be mediated by the apoptosis-inducing molecule of FasL [72] (Figure 2C). This evidence suggests that the potential interplay between antigen-specific T and B cell-derived exosomes may contribute to maintaining coordination in tumor immunity. Similarly, there is less evidence related to NK cell-derived exosomes affecting the biological behaviors of HNSCC, while TDEs regulate NK cells through cell–cell communication. For example, Wang et al. found that NAP1 of TDEs enhances the cytotoxicity of NK cells through the IRF-3 pathway [26]. Clinically, Ludwig et al. detected that exosomes in HNSCC patients reduce the expression levels of NKG2D in NK cells [73], indicating the possibility of plasma exosomes as biomarkers for the prognostic prediction of HNSCC. Taken together, these stromal cell-derived exosomes promote tumor progression and attenuate the therapeutic effects by altering the physiological state of recipient cells, indicating that targeting stromal cell-derived exosomes might be an effective strategy for anti-tumor treatment in HNSCC.
5. Theranostic Applications of Exosomes in HNSCC
5.1. Application of Exosomes in the Diagnosis of HNSCC
Clinically, histopathological and radiological results are widely used in diagnosing HNSCC. However, since the biopsy has inevitable side effects, minimally invasive detection of targeting exosomes in early tumors has become an important research field. CD9, caveolin 1 (CAV1), and tumor rejection antigen 1 (gp96) proteins, which undergo exosomal shuttling, have great diagnostic value in HNSCC [74]. By comparing eight different HNSCC cell lines with normal human oral keratinocytes, Qadir et al. found that the centrosome protein CEP55 is detectable in tumor exosomes but not normal cells [75], indicating that exosomal CEP55 may serve as a potential diagnostic marker for HNSCC. Regarding miRNAs in exosomes, Langevin et al. found that in exosomes isolated from HNSCC patients, the transcription levels of some TDE-enclosed miRNAs, including miR-486-5p, miR-486-3p, and miR-10b-5p, were significantly increased when compared with exosomes isolated from noncancer patients and control cells [76], suggesting that specific TDE-shuttled miRNAs may be valuable diagnostic biomarkers for HNSCC. Recently, such exosomes have been isolated from serum, plasma, and saliva for diagnosing HNSCC (Table 2). In human papilloma virus-16 (HPV-16)-associated oropharyngeal cancer, Kannan and colleagues found that serum exosomal HPV-16-E7 and SIRPA protein levels were higher than in healthy controls [77], illustrating that this approach may be used for early cancer diagnosis. Furthermore, Li et al. found that miR-21 levels were significantly higher in the circulating serum exosomes of OSCC patients than of healthy volunteers [36]. Li et al. demonstrated that PF4V1, CXCL7, F13A1, and ApoA1 from serum exosomes might be related to the metastasis of OSCC, which would help diagnose lymph node metastasis in OSCC [78]. For the plasma-derived exosomes, Schröck et al. demonstrated that SEPT9 and SHOX2 DNA methylation levels of ccfDNA from plasma could be biomarkers for diagnosis in HNSCC patients [79]. Of note, He et al. found a significant increase in salivary exosomal miR-24-3p in 45 preoperative OSCC patients compared with 10 normal controls [80], indicating the potential of salivary exosomal miR-24-3p as a diagnostic biomarker for OSCC. In summary, these results suggest that exosomes and their cargo from serum, plasma, and saliva might be used as biomarkers for the noninvasive diagnosis of HNSCC.
5.2. Application of Exosomes in the Prognostic Prediction of HNSCC
Studies have reported that exosomes and exosomal components can be used as biomarkers for cancer prognosis. For example, plasma-derived EBV DNA has been considered a classic biomarker for prognosis and monitoring response in the clinical treatment of HNSCC [89,90]. Previous studies have found that exosomal miR-21 levels were associated with tumor stage and lymph node metastasis in patients with oral, laryngeal, and esophageal SCC [36,91,92]. Similarly, Theodoraki et al. isolated exosomes from the plasma of HNSCC patients and found higher exosomal PD-L1 expression in patients locating in UICC stage III/IV or with positive lymph nodes than those patients who were UICC stage I/II or without lymph node metastasis [93], indicating that the expression of PD-L1 in exosome circulation may serve as a biomarker for predicting tumor prognosis. Other types of exosomal cargo, including epidermal growth factor receptor (EGFR), phosphorylated EGFR, HSP90, and Fas, have been found to have potential as prognostic biomarkers of HNSCC [24,94,95]. Notably, since no single exosomal biomarker is specific enough for use in predicting the prognosis, Li et al. suggested that a combination of CD9, CD63, and HSP70 can be used as predictors of lymph node metastasis in OSCC [78]. The aforementioned evidence indicates that exosomes and their cargo may be promising biomarkers for prognostic prediction, whereas the potential of more specific exosomal markers requires further exploration dependent on improved separation and purification technologies.
5.3. Application of Exosomes in the Treatment of HNSCC
TDEs are cell-derived nanoparticles that, due to their low immunogenicity, strong ability to cross physiological barriers, good biological distribution, and bioavailability, play an important role as potential anticancer drug carriers [96]. For example, in NPC, exosomal miR-34c enhances radiosensitivity mainly through inhibiting EMT and targeting β-catenin to suppress the development of malignancy, suggesting that the combination of IR and miR-34c-overexpressing exosomes may be an effective approach for radioresistant patients [97]. In another study, Jiang et al. found that exosomal miR-197-3p not only reduced proliferation and migration of NPC cells, but also reduced tumor growth and radiation resistance of NPC cells by regulating AKT/mTOR phosphorylation activation and HSPA5-mediated autophagy [98]. Similarly, in OSCC, exosomal-mediated miR-30a transfer in regaining sensitivity of cisplatin-resistant cancer cells’ induced apoptosis and autophagy via Beclin1 and Bcl2 regulation [99]. These evidences suggest that some exosomal miRNAs might be the biomarkers of increased overall survival and/or favorable outcome in HNSCC. In head and neck cancer, Concha et al. found that JAK2/STAT1 plays a role in EGFR-mediated immune evasion, and that targeting this signal may be beneficial in blocking PD-L1 upregulation [100]. In oral cancer, Xie et al. confirmed that human bone marrow mesenchymal stem cell-derived exosomes loaded with miR-101-3p regulated MMP2 levels and inhibited migration and invasion of OSCC cells in vitro and in vivo [101], providing a new theoretical basis for treating oral cancer. Additionally, Li et al. found that oxygen pressure in tumor microenvironments regulates myeloid-derived suppressor cell (MDSC) function in an miR-21/PTEN/PD-L1 axis-dependent manner by altering the TDE content to coordinate the antitumor and tumor-promoting γδt cell balance in OSCC [102]. The aforementioned observations demonstrate that exosomes represent bioavailable vehicles that can deliver drugs, proteins, miRNAs, and other molecules to prevent tumor progression and improve therapeutic efficacy via crosstalk between tumor cells and stromal cells.
6. Challenges and Future Perspectives
The crosstalk between the epithelium and stroma provides a suitable microenvironment for tumor progression in HNSCC. Exosomes act as mediators of intercellular communication with the advantage of presence in all body fluids, stable biological activity, biological compatibility, diverse sensitivity, and having specific molecules, including proteins and miRNAs, which have effects on many aspects of intercellular substance and signal transduction, including tumorigenesis, development, metastasis, recurrence, treatment of drug resistance, radiation resistance, and immunosuppression in HNSCC. Although TDEs have already been extensively studied in the context of pathogenesis and cancer development, we must broaden our understanding of stroma-derived exosomes and their functional roles in HNSCC. In HNSCC, exosomes have recently emerged as promising and valuable therapeutic targets for antitumor therapy. However, there are still multiple hurdles that must be overcome for the translation of results from bench to clinic. First, a standard method for the separation and purification of exosomes from different liquid samples is urgently needed. Second, the functional mechanisms of exosomes and their contents, especially in the case of stroma-derived exosomes, remain largely unknown. Whether functional exosomes are derived from tumor cells or the host response to the tumor must be determined and confirmed. Third, exosomes have the potential to be used as biomarkers for early diagnosis, nanoscale carriers for antitumor drugs, and tools for prognostic prediction; however, few examples have reached the stage of clinical application. Since exosomes are a highly heterogeneous population of membrane vesicles and no highly specific biomarkers have yet been confirmed to be suitable for use in prediction, an alternative approach combining several biomarkers for preclinical experiments or applications is needed. Overall, information on the specificity, safety, efficacy, and functional properties of exosomes is urgently needed before TDEs and/or stroma-derived exosomes can serve as targeting candidates for anticancer therapy in HNSCC.
Author Contributions
F.W. conceived and designed the study; Y.L. and S.G. drafted the manuscript; S.G., Y.L., Q.H. and F.W. searched and reviewed the literatures, and made the figures and tables. All of the authors critically reviewed and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. 82002884), National Key Research and Development Program of China (No. 2020YFA0714001), Science and Technology Program of Chengdu City (No. 2021-YF05-02031-SN), and Scientific Research Project of Ningxia colleges and universities (No. NGY2020039).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
A part of the figures were created with BioRender.com. Due to space limitations, we apologize for not being able to cite and discuss all relevant work from different laboratories published to date.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| bFGF | basic fibroblastic growth factor |
| CAFs | cancer-associated fibroblasts |
| CAV1 | caveolin 1 |
| CTL | cytotoxic T lymphocyte |
| CAFs | cancer-associated fibroblasts |
| DNA | deoxyribonucleic aid |
| ECs | endothelial cells |
| EGFR | epidermal growth factor receptor |
| EMT | mesenchymal transition |
| ESCC | esophageal squamous cell carcinoma |
| ER | endoplasmic reticulum |
| EVs | extracellular vehicles |
| HIF-1α | hypoxia inducible factor 1 alpha |
| HNSCC | head and neck squamous cell carcinoma |
| HNC | head and neck cancer |
| HUVECs | human umbilical vein endothelial cells |
| HSP90 | heat shock protein90 |
| LCL | lymphoblastic-like cell line |
| LSCC | laryngeal squamous cell carcinoma |
| LOXL2 | lysyl oxidase like 2 |
| MDSC | myeloid-derived suppressor cell |
| MFAP5 | microfibrillar-associated protein 5 |
| MSCs | mesenchymal stem cells |
| MMP13 | matrix metalloproteinase-13 |
| mini-SEC | mini size-exclusion chromatography |
| mRNA | messenger ribonucleic acid |
| miRNA | micro ribonucleic acid |
| N/A | not available |
| NADDP1 | nucleosome assembly protein 1 |
| NK | nature killer |
| NKG2D | natural killer Group 2 member D |
| NPC | nasopharyngeal carcinoma |
| 4NQO | 4-nitroquinoline 1-oxide |
| OSCC | oral squamous cell carcinoma |
| PD-L1 | programmed death ligand 1 |
| PVOPC | human papilloma virus-16 associated oropharyngeal cancer |
| PDCD4 | programmed cell death 4 |
| PTEN | phosphatase and tensin homolog |
| RNA | ribonucleic acid |
| SEC | size exclusion chromatography |
| TAMs | tumor associated macrophages |
| TEM | transmission electron microscopy |
| TDEs | tumor-derived exosomes |
| TME | tumor microenvironment |
| VEGF | vascular endothelial growth factor |
| WB | western blot |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebnoether, E.; Muller, L. Diagnostic and Therapeutic Applications of Exosomes in Cancer with a Special Focus on Head and Neck Squamous Cell Carcinoma (HNSCC). Int. J. Mol. Sci. 2020, 21, 4344. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Sun, X.; Chen, Z.; Du, J.; Wu, Y. Head and Neck Squamous Cell Carcinoma: Risk Factors, Molecular Alterations, Immunology and Peptide Vaccines. Int. J. Pept. Res. Ther. 2022, 28, 19. [Google Scholar] [CrossRef]
- Mishra, V.; Singh, A.; Chen, X.; Rosenberg, A.J.; Pearson, A.T.; Zhavoronkov, A.; Savage, P.A.; Lingen, M.W.; Agrawal, N.; Izumchenko, E. Application of liquid biopsy as multi-functional biomarkers in head and neck cancer. Br. J. Cancer 2022, 126, 361–370. [Google Scholar] [CrossRef]
- Knops, A.M.; South, A.; Rodeck, U.; Martinez-Outschoorn, U.; Harshyne, L.A.; Johnson, J.; Luginbuhl, A.J.; Curry, J.M. Cancer-Associated Fibroblast Density, Prognostic Characteristics, and Recurrence in Head and Neck Squamous Cell Carcinoma: A Meta-Analysis. Front. Oncol. 2020, 10, 565306. [Google Scholar] [CrossRef]
- Economopoulou, P.; Perisanidis, C.; Giotakis, E.I.; Psyrri, A. The emerging role of immunotherapy in head and neck squamous cell carcinoma (HNSCC): Anti-tumor immunity and clinical applications. Ann. Transl. Med. 2016, 4, 173. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Yang, J.; Liu, J.; Wang, Y.; Mu, J.; Zeng, Q.; Deng, S.; Zhou, H. Signaling pathways in cancer-associated fibroblasts and targeted therapy for cancer. Signal. Transduct. Target. Ther. 2021, 6, 218. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, 6977. [Google Scholar] [CrossRef]
- Xie, Z.; Gao, Y.; Ho, C.; Li, L.; Jin, C.; Wang, X.; Zou, C.; Mao, Y.; Wang, X.; Li, Q.; et al. Exosome-delivered CD44v6/C1QBP complex drives pancreatic cancer liver metastasis by promoting fibrotic liver microenvironment. Gut 2022, 71, 568–579. [Google Scholar] [CrossRef]
- Nonaka, T.; Wong, D.T.W. Liquid Biopsy in Head and Neck Cancer: Promises and Challenges. J. Dent. Res. 2018, 97, 701–708. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, L.; Shen, N.; Ning, X.; Wu, D.; Jiang, K.; Huang, X. M1 macrophage-derived exosomes and their key molecule lncRNA HOTTIP suppress head and neck squamous cell carcinoma progression by upregulating the TLR5/NF-κB pathway. Cell Death Dis. 2022, 13, 183. [Google Scholar] [CrossRef] [PubMed]
- Nikolopoulou, P.A.; Koufaki, M.A.; Kostourou, V. The Adhesome Network: Key Components Shaping the Tumour Stroma. Cancers 2021, 13, 525. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Sang, Y.; Song, X.; Zhang, D.; Wang, L.; Zhao, W.; Liang, Y.; Zhang, N.; Yang, Q. Exosomal miR-500a-5p derived from cancer-associated fibroblasts promotes breast cancer cell proliferation and metastasis through targeting USP28. Theranostics 2021, 11, 3932–3947. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Chen, L.; Yuan, X.; Luo, Q.; Liu, Y.; Xie, G.; Ma, Y.; Shen, L. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J. Exp. Clin. Cancer Res. 2017, 36, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, M.; Ren, X. New insights into the biological impacts of immune cell-derived exosomes within the tumor environment. Cancer Lett. 2018, 431, 115–122. [Google Scholar] [CrossRef]
- Tomita, R.; Sasabe, E.; Tomomura, A.; Yamamoto, T. Macrophage-derived exosomes attenuate the susceptibility of oral squamous cell carcinoma cells to chemotherapeutic drugs through the AKT/GSK3beta pathway. Oncol. Rep. 2020, 44, 1905–1916. [Google Scholar]
- Min, H.; Sun, X.; Yang, X.; Zhu, H.; Liu, J.; Wang, Y.; Chen, G.; Sun, X. Exosomes Derived from Irradiated Esophageal Carcinoma-Infiltrating T Cells Promote Metastasis by Inducing the Epithelial-Mesenchymal Transition in Esophageal Cancer Cells. Pathol. Oncol. Res. 2018, 24, 11–18. [Google Scholar] [CrossRef]
- Tian, Y.; Li, S.; Song, J.; Ji, T.; Zhu, M.; Anderson, G.J.; Wei, J.; Nie, G. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014, 35, 2383–2390. [Google Scholar] [CrossRef]
- Zheng, R.; Zhang, K.; Tan, S.; Gao, F.; Zhang, Y.; Xu, W.; Wang, H.; Gu, D.; Zhu, L.; Li, S.; et al. Exosomal circLPAR1 functions in colorectal cancer diagnosis and tumorigenesis through suppressing BRD4 via METTL3-eIF3h interaction. Mol. Cancer 2022, 21, 49. [Google Scholar] [CrossRef]
- Taverna, S.; Pucci, M.; Alessandro, R. Extracellular vesicles: Small bricks for tissue repair/regeneration. Ann. Transl. Med. 2017, 5, 83. [Google Scholar] [CrossRef] [Green Version]
- Azulay, E.E.; Cooks, T.; Elkabets, M. Potential oncogenic roles of mutant-p53-derived exosomes in the tumor-host interaction of head and neck cancers. Cancer Immunol. Immunother. 2020, 69, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Wu, Y.; Wu, Y.; Wang, P.; Vadgama, J.V. Tumor-Derived Exosomes in Tumor-Induced Immune Suppression. Int. J. Mol. Sci. 2022, 23, 1461. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.; Xie, Y.; Huang, M.; Gu, J. Tumor-derived exosomes in hypoxic microenvironment: Release mechanism, biological function and clinical application. J. Cancer 2022, 13, 1685–1694. [Google Scholar] [CrossRef]
- Ono, K.; Eguchi, T.; Sogawa, C.; Calderwood, S.K.; Futagawa, J.; Kasai, T.; Seno, M.; Okamoto, K.; Sasaki, A.; Kozaki, K.I. HSP-enriched properties of extracellular vesicles involve survival of metastatic oral cancer cells. J. Cell Biochem. 2018, 119, 7350–7362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Gillespie, D.G.; Menshikova, E.V.; Jackson, E.K.; Whiteside, T.L. Tumor-derived exosomes promote angiogenesis via adenosine A2B receptor signaling. Angiogenesis 2020, 23, 599–610. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, X.; Zhu, X.; Chen, W.; Zhang, J.; Chen, W. Oral cancer-derived exosomal NAP1 enhances cytotoxicity of natural killer cells via the IRF-3 pathway. Oral Oncol. 2018, 76, 34–41. [Google Scholar] [CrossRef]
- Ono, K.; Sogawa, C.; Kawai, H.; Tran, M.T.; Taha, E.A.; Lu, Y.; Oo, M.W.; Okusha, Y.; Okamura, H.; Ibaragi, S.; et al. Triple knockdown of CDC37, HSP90-alpha and HSP90-beta diminishes extracellular vesicles-driven malignancy events and macrophage M2 polarization in oral cancer. J. Extracell. Vesicles 2020, 9, 1769373. [Google Scholar] [CrossRef]
- Ludwig, N.; Gillespie, D.G.; Reichert, T.E.; Jackson, E.K.; Whiteside, T.L. Purine Metabolites in Tumor-Derived Exosomes May Facilitate Immune Escape of Head and Neck Squamous Cell Carcinoma. Cancers 2020, 12, 1602. [Google Scholar] [CrossRef]
- Shan, Y.; You, B.; Shi, S.; Shi, W.; Zhang, Z.; Zhang, Q.; Gu, M.; Chen, J.; Bao, L.; Liu, D.; et al. Hypoxia-Induced Matrix Metalloproteinase-13 Expression in Exosomes from Nasopharyngeal Carcinoma Enhances Metastases. Cell Death Dis. 2018, 9, 382. [Google Scholar] [CrossRef]
- Ye, S.-B.; Li, Z.-L.; Luo, D.-H.; Huang, B.-J.; Chen, Y.-S.; Zhang, X.-S.; Cui, J.; Zeng, Y.-X.; Li, J. Tumor-derived exosomes promote tumor progression and T-cell dysfunction through the regulation of enriched exosomal microRNAs in human nasopharyngeal carcinoma. Oncotarget 2014, 5, 5439–5452. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.; Shi, S.; Yue, H.; You, B.; Shan, Y.; Zhu, Z.; Bao, L.; You, Y. Exosomal miR-17–5p promotes angiogenesis in nasopharyngeal carcinoma via targeting BAMBI. J. Cancer 2019, 10, 6681–6692. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Zhang, J.; Chen, W.; Chen, W. M1-like tumor-associated macrophages activated by exosome-transferred THBS1 promote malignant migration in oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzo, B.M.; Ludwig, N.; Hong, C.S.; Sharma, P.; Fabian, K.P.; Fecek, R.J.; Storkus, W.J.; Whiteside, T.L. Tumor-derived exosomes promote carcinogenesis of murine oral squamous cell carcinoma. Carcinogenesis 2020, 41, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Sakha, S.; Muramatsu, T.; Ueda, K.; Inazawa, J. Exosomal microRNA miR-1246 induces cell motility and invasion through the regulation of DENND2D in oral squamous cell carcinoma. Sci. Rep. 2016, 6, 38750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, L.; Zhou, X.; Luo, X.; Liu, K.; Jiang, E.; Chen, Y.; Shao, Z.; Shang, Z. OSCC Exosomes Regulate miR-210–3p Targeting EFNA3 to Promote Oral Cancer Angiogenesis through the PI3K/AKT Pathway. Biomed. Res. Int. 2020, 2020, 2125656. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Chen, G.; Sun, D.; Lei, M.; Li, Y.; Zhou, C.; Li, X.; Xue, W.; Wang, H.; Liu, C.; et al. Exosomes containing miR-21 transfer the characteristic of cisplatin resistance by targeting PTEN and PDCD4 in oral squamous cell carcinoma. Acta Biochim. Biophys. Sin. 2017, 49, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Guo, H.; Wang, X.; Zhu, X.; Yan, M.; Wang, X.; Xu, Q.; Shi, J.; Lu, E.; Chen, W.; et al. Exosomal miR-196a derived from cancer-associated fibroblasts confers cisplatin resistance in head and neck cancer through targeting CDKN1B and ING5. Genome Biol. 2019, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qin, X.; Yan, M.; Shi, J.; Xu, Q.; Li, Z.; Yang, W.; Zhang, J.; Chen, W. Loss of exosomal miR-3188 in cancer-associated fibroblasts contributes to HNC progression. J. Exp. Clin. Cancer Res. 2019, 38, 151. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-Y.; Tao, Y.-W.; Gao, S.; Li, P.; Zheng, J.-M.; Zhang, S.-E.; Liang, J.; Zhang, Y. Cancer-associated fibroblasts contribute to oral cancer cells proliferation and metastasis via exosome-mediated paracrine miR-34a-5p. EBioMedicine 2018, 36, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Ai, Y.; Wei, H.; Wu, S.; Tang, Z.; Li, X.; Zou, C. Exosomal LncRNA LBX1-AS1 Derived from RBPJ Overexpressed-Macrophages Inhibits Oral Squamous Cell Carcinoma Progress via miR-182–5p/FOXO3. Front. Oncol. 2021, 11, 605884. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Wang, S.-S.; Zhang, M.; Jiang, J.; Fan, H.-Y.; Wu, J.-S.; Wang, H.-F.; Liang, X.-H.; Tang, Y.-L. OSCC cell-secreted exosomal CMTM6 induced M2-like macrophages polarization via ERK1/2 signaling pathway. Cancer Immunol. Immunother. 2021, 70, 1015–1029. [Google Scholar] [CrossRef] [PubMed]
- Patel-Hett, S.; D’Amore, P.A. Signal transduction in vasculogenesis and developmental angiogenesis. Int. J. Dev. Biol. 2011, 55, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, N.; Kerbel, R.S. Angiogenesis as a therapeutic target. Nature 2005, 438, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Al-Nedawi, K.; Meehan, B.; Kerbel, R.S.; Allison, A.C.; Rak, J. Endothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFR. Proc. Natl. Acad. Sci. USA 2009, 106, 3794–3799. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; De Veirman, K.; Faict, S.; Frassanito, M.A.; Ribatti, D.; Vacca, A.; Menu, E. Multiple myeloma exosomes establish a favourable bone marrow microenvironment with enhanced angiogenesis and immunosuppression. J. Pathol. 2016, 239, 162–173. [Google Scholar] [CrossRef]
- Bao, L.; You, B.; Shi, S.; Shan, Y.; Zhang, Q.; Yue, H.; Zhang, J.; Zhang, W.; Shi, Y.; Liu, Y.; et al. Metastasis-associated miR-23a from nasopharyngeal carcinoma-derived exosomes mediates angiogenesis by repressing a novel target gene TSGA10. Oncogene 2018, 37, 2873–2889. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Q.-H.; Wang, F.; Tan, J.-J.; Deng, Y.-Q.; Peng, X.-H.; Liu, X.; Zhang, B.; Xu, X.; Li, X.-P. Exosomal miR-9 inhibits angiogenesis by targeting MDK and regulating PDK/AKT pathway in nasopharyngeal carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 147. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, G.; Wu, X.; Jiang, Z.; Kasman, I.; Yao, J.; Guan, Y.; Oeh, J.; Modrusan, Z.; Bais, C.; Sampath, D.; et al. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J. 2012, 31, 3513–3523. [Google Scholar] [CrossRef]
- Hosni, A.; Huang, S.H.; Chiu, K.; Xu, W.; Su, J.; Bayley, A.; Bratman, S.V.; Cho, J.; Giuliani, M.; Kim, J.; et al. Predictors of Early Recurrence Prior to Planned Postoperative Radiation Therapy for Oral Cavity Squamous Cell Carcinoma and Outcomes Following Salvage Intensified Radiation Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2019, 103, 363–373. [Google Scholar] [CrossRef]
- Cai, J.; Qiao, B.; Gao, N.; Lin, N.; He, W. Oral squamous cell carcinoma-derived exosomes promote M2 subtype macrophage polarization mediated by exosome-enclosed miR-29a-3p. Am. J. Physiol. Cell Physiol. 2019, 316, C731–C740. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Liu, R.; Shi, Y.-J.; Yin, L.-H.; Pu, Y.-P. Exosome-shuttling microRNA-21 promotes cell migration and invasion-targeting PDCD4 in esophageal cancer. Int. J. Oncol. 2016, 48, 2567–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Liu, C.; Su, K.; Wang, J.; Liu, Y.; Zhang, L.; Li, C.; Cong, Y.; Kimberly, R.; Grizzle, W.E.; et al. Tumor exosomes inhibit differentiation of bone marrow dendritic cells. J. Immunol. 2007, 178, 6867–6875. [Google Scholar] [CrossRef] [Green Version]
- Namee, N.M.; O’Driscoll, L. Extracellular vesicles and anti-cancer drug resistance. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Steinbichler, T.B.; Dudás, J.; Skvortsov, S.; Ganswindt, U.; Riechelmann, H.; Skvortsova, I.-I. Therapy resistance mediated by exosomes. Mol. Cancer 2019, 18, 58. [Google Scholar] [CrossRef]
- Huang, D.; Bian, G.; Pan, Y.; Han, X.; Sun, Y.; Wang, Y.; Shen, G.; Cheng, M.; Fang, X.; Hu, S. MiR-20a-5p promotes radio-resistance by targeting Rab27B in nasopharyngeal cancer cells. Cancer Cell Int. 2017, 17, 32. [Google Scholar] [CrossRef] [Green Version]
- Beck, C.; Piontek, G.; Haug, A.; Bas, M.; Knopf, A.; Stark, T.; Mißlbeck, M.; Rudelius, M.; Reiter, R.; Brandstetter, M.; et al. The kallikrein-kinin-system in head and neck squamous cell carcinoma (HNSCC) and its role in tumour survival, invasion, migration and response to radiotherapy. Oral Oncol. 2012, 48, 1208–1219. [Google Scholar] [CrossRef]
- Principe, S.; Mejia-Guerrero, S.; Ignatchenko, V.; Sinha, A.; Ignatchenko, A.; Shi, W.; Pereira, K.; Su, S.; Huang, S.H.; O’Sullivan, B.; et al. Proteomic Analysis of Cancer-Associated Fibroblasts Reveals a Paracrine Role for MFAP5 in Human Oral Tongue Squamous Cell Carcinoma. J. Proteome Res. 2018, 17, 2045–2059. [Google Scholar] [CrossRef]
- Sun, L.-P.; Xu, K.; Cui, J.; Yuan, D.-Y.; Zou, B.; Li, J.; Liu, J.-L.; Li, K.-Y.; Meng, Z.; Zhang, B. Cancer-associated fibroblast-derived exosomal miR3825p promotes the migration and invasion of oral squamous cell carcinoma. Oncol. Rep. 2019, 42, 1319–1328. [Google Scholar]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Au Yeung, C.L.; Co, N.-N.; Tsuruga, T.; Yeung, T.-L.; Kwan, S.-Y.; Leung, C.S.; Li, Y.; Lu, E.S.; Kwan, K.; Wong, K.-K.; et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun. 2016, 7, 11150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.D.; Koo, B.-H.; Kim, Y.H.; Jeon, O.-H.; Kim, D.-S. Exosome release of ADAM15 and the functional implications of human macrophage-derived ADAM15 exosomes. FASEB J. 2012, 26, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Matsuba, T.; Niki, T.; Hattori, T. Stimulation of THP-1 Macrophages with LPS Increased the Production of Osteopontin-Encapsulating Exosome. Int. J. Mol. Sci. 2020, 21, 8490. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 81. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Ma, T.; Huang, B.; Lin, L.; Zhou, Y.; Yan, J.; Zou, Y.; Chen, S. Macrophage-derived exosomal microRNA-501–3p promotes progression of pancreatic ductal adenocarcinoma through the TGFBR3-mediated TGF-β signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 310. [Google Scholar] [CrossRef] [Green Version]
- Veerman, R.E.; Akpinar, G.G.; Eldh, M.; Gabrielsson, S. Immune Cell-Derived Extracellular Vesicles—Functions and Therapeutic Applications. Trends Mol. Med. 2019, 25, 382–394. [Google Scholar] [CrossRef]
- Muller, L.; Simms, P.; Hong, C.S.; Nishimura, M.I.; Jackson, E.K.; Watkins, S.C.; Whiteside, T.L. Human tumor-derived exosomes (TEX) regulate Treg functions via cell surface signaling rather than uptake mechanisms. Oncoimmunology 2017, 6, e1261243. [Google Scholar] [CrossRef]
- Seo, N.; Shirakura, Y.; Tahara, Y.; Momose, F.; Harada, N.; Ikeda, H.; Akiyoshi, K.; Shiku, H. Activated CD8+ T cell extracellular vesicles prevent tumour progression by targeting of lesional mesenchymal cells. Nat. Commun. 2018, 9, 435. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhang, H.; Li, W.; Deng, Y.; Munegowda, M.A.; Chibbar, R.; Qureshi, M.; Xiang, J. Dendritic cells recruit T cell exosomes via exosomal LFA-1 leading to inhibition of CD8+ CTL responses through downregulation of peptide/MHC class I and Fas ligand-mediated cytotoxicity. J. Immunol. 2010, 185, 5268–5278. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Singh, S.P.; Elkahloun, A.G.; Wu, W.; Abu-Asab, M.S.; Roberts, D.D. CD47-dependent immunomodulatory and angiogenic activities of extracellular vesicles produced by T cells. Matrix Biol. 2014, 37, 49–59. [Google Scholar] [CrossRef]
- Saunderson, S.C.; McLellan, A.D. Role of Lymphocyte Subsets in the Immune Response to Primary B Cell-Derived Exosomes. J. Immunol. 2017, 199, 2225–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinker, M.W.; Lizzio, V.; Reed, T.J.; Fox, D.A.; Lundy, S.K. Human B Cell-Derived Lymphoblastoid Cell Lines Constitutively Produce Fas Ligand and Secrete MHCII+FasL+ Killer Exosomes. Front. Immunol. 2014, 5, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, S.; Floros, T.; Theodoraki, M.N.; Hong, C.S.; Jackson, E.K.; Lang, S.; Whiteside, T.L. Suppression of Lymphocyte Functions by Plasma Exosomes Correlates with Disease Activity in Patients with Head and Neck Cancer. Clin. Cancer Res. 2017, 23, 4843–4854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar]
- Qadir, F.; Aziz, M.A.; Sari, C.P.; Ma, H.; Dai, H.; Wang, X.; Raithatha, D.; Da Silva, L.G.L.; Hussain, M.; Poorkasreiy, S.P.; et al. Transcriptome reprogramming by cancer exosomes: Identification of novel molecular targets in matrix and immune modulation. Mol. Cancer 2018, 17, 97. [Google Scholar] [CrossRef] [Green Version]
- Langevin, S.; Kuhnell, D.; Parry, T.; Biesiada, J.; Huang, S.; Wise-Draper, T.; Casper, K.; Zhang, X.; Medvedovic, M.; Kasper, S. Comprehensive microRNA-sequencing of exosomes derived from head and neck carcinoma cells in vitro reveals common secretion profiles and potential utility as salivary biomarkers. Oncotarget 2017, 8, 82459–82474. [Google Scholar] [CrossRef] [Green Version]
- Kannan, A.; Hertweck, K.L.; Philley, J.V.; Wells, R.B.; Dasgupta, S. Genetic Mutation and Exosome Signature of Human Papilloma Virus Associated Oropharyngeal Cancer. Sci. Rep. 2017, 7, 46102. [Google Scholar] [CrossRef]
- Li, C.; Zhou, Y.; Liu, J.; Su, X.; Qin, H.; Huang, S.; Huang, X.; Zhou, N. Potential Markers from Serum-Purified Exosomes for Detecting Oral Squamous Cell Carcinoma Metastasis. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1668–1681. [Google Scholar] [CrossRef] [Green Version]
- Schröck, A.; Leisse, A.; de Vos, L.; Gevensleben, H.; Dröge, F.; Franzen, A.; Wachendörfer, M.; Schröck, F.; Ellinger, J.; Teschke, M.; et al. Free-Circulating Methylated DNA in Blood for Diagnosis, Staging, Prognosis, and Monitoring of Head and Neck Squamous Cell Carcinoma Patients: An Observational Prospective Cohort Study. Clin. Chem. 2017, 63, 1288–1296. [Google Scholar] [CrossRef]
- He, L.; Ping, F.; Fan, Z.; Zhang, C.; Deng, M.; Cheng, B.; Xia, J. Salivary exosomal miR-24–3p serves as a potential detective biomarker for oral squamous cell carcinoma screening. Biomed. Pharmacother. 2020, 121, 109553. [Google Scholar] [CrossRef]
- Theodoraki, M.N.; Yerneni, S.; Gooding, W.E.; Ohr, J.; Clump, D.A.; Bauman, J.E.; Ferris, R.L.; Whiteside, T.L. Circulating exosomes measure responses to therapy in head and neck cancer patients treated with cetuximab, ipilimumab, and IMRT. Oncoimmunology 2019, 8, 1593805. [Google Scholar] [CrossRef] [PubMed]
- Beccard, I.J.; Hofmann, L.; Schroeder, J.C.; Ludwig, S.; Laban, S.; Brunner, C.; Lotfi, R.; Hoffmann, T.K.; Jackson, E.K.; Schuler, P.J.; et al. Immune Suppressive Effects of Plasma-Derived Exosome Populations in Head and Neck Cancer. Cancers 2020, 12, 1997. [Google Scholar] [CrossRef] [PubMed]
- Theodoraki, M.N.; Laban, S.; Jackson, E.K.; Lotfi, R.; Schuler, P.J.; Brunner, C.; Hoffmann, T.K.; Whiteside, T.L.; Hofmann, L. Changes in circulating exosome molecular profiles following surgery/(chemo)radiotherapy: Early detection of response in head and neck cancer patients. Br. J. Cancer 2021, 125, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Theodoraki, M.N.; Matsumoto, A.; Beccard, I.; Hoffmann, T.K.; Whiteside, T.L. CD44v3 protein-carrying tumor-derived exosomes in HNSCC patients’ plasma as potential noninvasive biomarkers of disease activity. Oncoimmunology 2020, 9, 1747732. [Google Scholar] [CrossRef] [Green Version]
- Sanada, T.; Islam, A.; Kaminota, T.; Kirino, Y.; Tanimoto, R.; Yoshimitsu, H.; Yano, H.; Mizuno, Y.; Okada, M.; Mitani, S.; et al. Elevated exosomal lysyl oxidase like 2 is a potential biomarker for head and neck squamous cell carcinoma. Laryngoscope 2020, 130, e327–e334. [Google Scholar] [CrossRef]
- Houali, K.; Wang, X.; Shimizu, Y.; Djennaoui, D.; Nicholls, J.; Fiorini, S.; Bouguermouh, A.; Ooka, T. A new diagnostic marker for secreted Epstein-Barr virus encoded LMP1 and BARF1 oncoproteins in the serum and saliva of patients with nasopharyngeal carcinoma. Clin. Cancer Res. 2007, 13, 4993–5000. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Guo, X.; Li, X.; Liao, C.; Wang, X.; He, K. Plasma-Derived Exosomal microRNA-130a Serves as a Noninvasive Biomarker for Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J. Oncol. 2021, 2021, 5547911. [Google Scholar] [CrossRef]
- Gai, C.; Camussi, F.; Broccoletti, R.; Gambino, A.; Cabras, M.; Molinaro, L.; Carossa, S.; Camussi, G.; Arduino, P.G. Salivary extracellular vesicle-associated miRNAs as potential biomarkers in oral squamous cell carcinoma. BMC Cancer 2018, 18, 439. [Google Scholar] [CrossRef] [Green Version]
- Gourzones, C.; Gelin, A.; Bombik, I.; Klibi, J.; Vérillaud, B.; Guigay, J.; Lang, P.; Témam, S.; Schneider, V.; Amiel, C.; et al. Extra-cellular release and blood diffusion of BART viral micro-RNAs produced by EBV-infected nasopharyngeal carcinoma cells. Virol. J. 2010, 7, 271. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Birkeland, A.C. Liquid Biopsies in Head and Neck Cancer: Current State and Future Challenges. Cancers 2021, 13, 1874. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Lu, J.; Sun, Y.; Xiao, H.; Liu, M.; Tian, L. Combined detection of serum exosomal miR-21 and HOTAIR as diagnostic and prognostic biomarkers for laryngeal squamous cell carcinoma. Med. Oncol. 2014, 31, 148. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kamohara, H.; Kinoshita, K.; Kurashige, J.; Ishimoto, T.; Iwatsuki, M.; Watanabe, M.; Baba, H. Clinical impact of serum exosomal microRNA-21 as a clinical biomarker in human esophageal squamous cell carcinoma. Cancer 2013, 119, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Theodoraki, M.N.; Yerneni, S.S.; Hoffmann, T.K.; Gooding, W.E.; Whiteside, T.L. Clinical Significance of PD-L1+ Exosomes in Plasma of Head and Neck Cancer Patients. Clin. Cancer Res. 2018, 24, 896–905. [Google Scholar] [PubMed] [Green Version]
- Kim, J.W.; Wieckowski, E.; Taylor, D.D.; Reichert, T.E.; Watkins, S.; Whiteside, T.L. Fas ligand-positive membranous vesicles isolated from sera of patients with oral cancer induce apoptosis of activated T lymphocytes. Clin. Cancer Res. 2005, 11, 1010–1020. [Google Scholar]
- Raulf, N.; Lucarelli, P.; Thavaraj, S.; Brown, S.; Vicencio, J.M.; Sauter, T.; Tavassoli, M. Annexin A1 regulates EGFR activity and alters EGFR-containing tumour-derived exosomes in head and neck cancers. Eur. J. Cancer 2018, 102, 52–68. [Google Scholar] [CrossRef] [Green Version]
- Arrighetti, N.; Corbo, C.; Evangelopoulos, M.; Pastò, A.; Zuco, V.; Tasciotti, E. Exosome-like Nanovectors for Drug Delivery in Cancer. Curr. Med. Chem. 2019, 26, 6132–6148. [Google Scholar] [CrossRef]
- Wan, F.-Z.; Chen, K.-H.; Sun, Y.-C.; Chen, X.-C.; Liang, R.-B.; Chen, L.; Zhu, X.-D. Exosomes overexpressing miR-34c inhibit malignant behavior and reverse the radioresistance of nasopharyngeal carcinoma. J. Transl. Med. 2020, 18, 12. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Tang, Q.; Gong, J.; Jiang, W.; Chen, Y.; Zhou, Q.; Aldeen, A.; Wang, S.; Li, C.; Lv, W.; et al. Radiosensitizer EXO-miR-197–3p Inhibits Nasopharyngeal Carcinoma Progression and Radioresistance by Regulating the AKT/mTOR Axis and HSPA5-mediated Autophagy. Int. J. Biol. Sci. 2022, 18, 1878–1895. [Google Scholar] [CrossRef]
- Kulkarni, B.; Gondaliya, P.; Kirave, P.; Rawal, R.; Jain, A.; Garg, R.; Kalia, K. Exosome-mediated delivery of miR-30a sensitize cisplatin-resistant variant of oral squamous carcinoma cells via modulating Beclin1 and Bcl2. Oncotarget 2020, 11, 1832–1845. [Google Scholar] [CrossRef]
- Concha-Benavente, F.; Srivastava, R.M.; Trivedi, S.; Lei, Y.; Chandran, U.; Seethala, R.R.; Freeman, G.J.; Ferris, R.L. Identification of the Cell-Intrinsic and -Extrinsic Pathways Downstream of EGFR and IFNγ That Induce PD-L1 Expression in Head and Neck Cancer. Cancer Res. 2016, 76, 1031–1043. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Du, L.Y.; Guo, F.; Li, X.; Cheng, B. Exosomes derived from microRNA-101–3p-overexpressing human bone marrow mesenchymal stem cells suppress oral cancer cell proliferation, invasion, and migration. Mol. Cell Biochem. 2019, 458, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cao, B.; Liang, X.; Lu, S.; Luo, H.; Wang, Z.; Wang, S.; Jiang, J.; Lang, J.; Zhu, G. Microenvironmental oxygen pressure orchestrates an anti- and pro-tumoral γδ T cell equilibrium via tumor-derived exosomes. Oncogene 2019, 38, 2830–2843. [Google Scholar] [CrossRef] [PubMed]
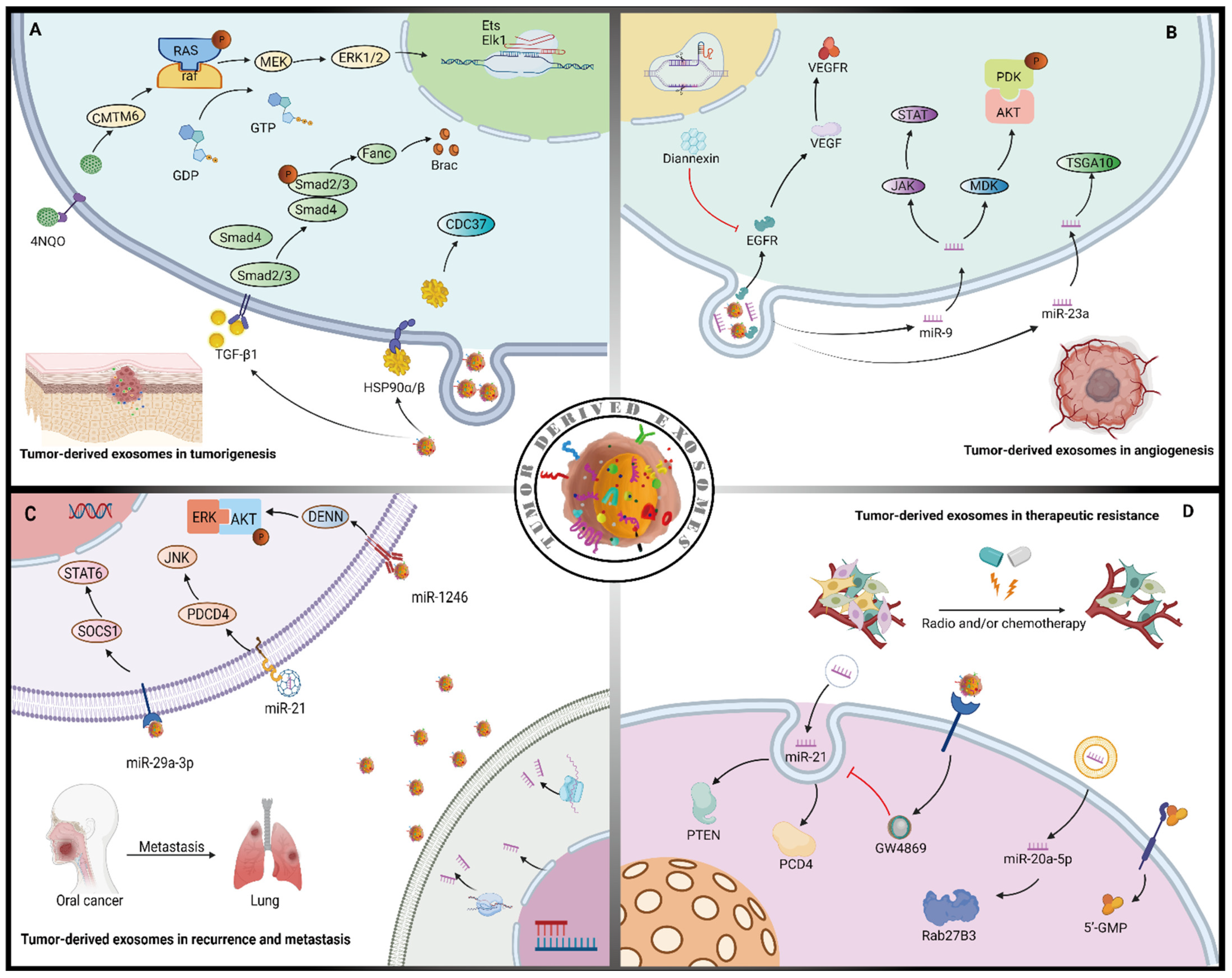
Figure 1.
The effects of tumor-derived exosomes in tumor progression. (A) Tumor-derived exosomes rich in protein (HSP90α/β,TGF-β1) transfer to adjacent cells to promote tumorigenesis through the immunosuppressive effects. Tumor-derived exosomes CMTM6 induce polarization of M2-like macrophages through the MEK/ERK1/2 signaling pathway. (B) Tumor-derived exosomes miRNA (miR-9, miR-23a) and protein (EGFR) raise the formation of new blood vessels during tumor progression via the autocrine VEGF/VEGFR pathway. (C) Tumor-derived exosomes miRNA (miR-29a-3p, miR-21, miR-1246), which can be transferred to recipient cells and increase the incidence of recurrence and metastasis of cancer by activating the STAT6, JNK, and ERK signaling pathways, respectively. (D) Tumor-derived exosomes miR-21 conveyed to parental cells can be attenuated by the inhibitor (GW4869) promoting chemoresistance. Tumor-derived exosomes miR-20a-5p transfer to adjacent cells leading to radiation tolerance. Back arrows: promotion; red “T” arrows: inhibition.
Figure 1.
The effects of tumor-derived exosomes in tumor progression. (A) Tumor-derived exosomes rich in protein (HSP90α/β,TGF-β1) transfer to adjacent cells to promote tumorigenesis through the immunosuppressive effects. Tumor-derived exosomes CMTM6 induce polarization of M2-like macrophages through the MEK/ERK1/2 signaling pathway. (B) Tumor-derived exosomes miRNA (miR-9, miR-23a) and protein (EGFR) raise the formation of new blood vessels during tumor progression via the autocrine VEGF/VEGFR pathway. (C) Tumor-derived exosomes miRNA (miR-29a-3p, miR-21, miR-1246), which can be transferred to recipient cells and increase the incidence of recurrence and metastasis of cancer by activating the STAT6, JNK, and ERK signaling pathways, respectively. (D) Tumor-derived exosomes miR-21 conveyed to parental cells can be attenuated by the inhibitor (GW4869) promoting chemoresistance. Tumor-derived exosomes miR-20a-5p transfer to adjacent cells leading to radiation tolerance. Back arrows: promotion; red “T” arrows: inhibition.

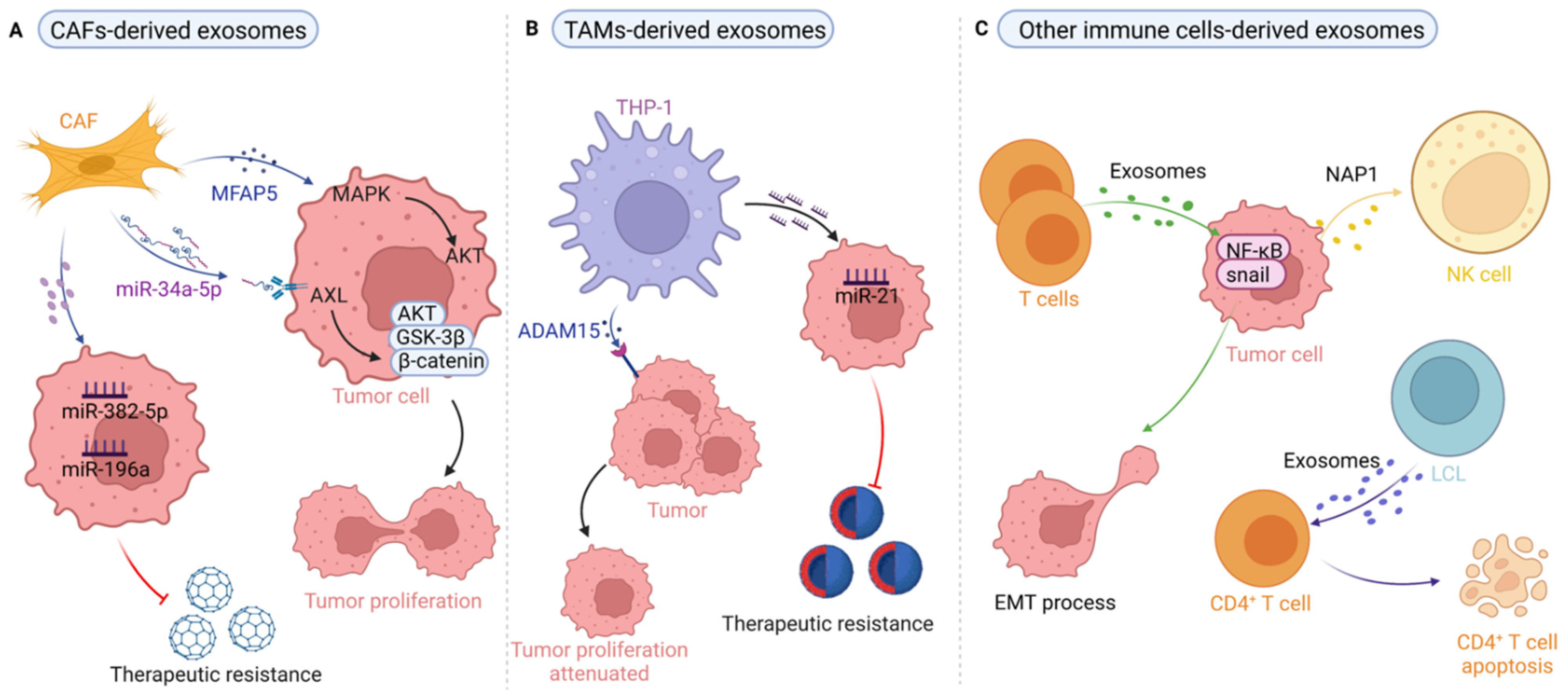
Figure 2.
The role of stroma-derived exosomes in tumor progression. (A) CAFs-derived exosomes rich in miRNA (miR-382-5p, miR-196a) transferred to tumor cells promote tumor cells migration and invasion, and induce therapeutic resistance. CAFs-derived exosomes miRNA (miR-34a-5p), protein (MFAP5) transfer to tumor cells to facilitate tumor cell proliferation. (B) TAMs-derived exosomes miRNA (miR-21) transfer to tumor cells and raise the survival and proliferation of tumor cells with induced therapeutic resistance. TAMs-derived exosomes protein (ADAM15), inhibited tumor proliferation. (C) T cell-derived exosomes promote the metastasis of tumor cells by inducing EMT. TDEs NAP1 enhance the cytotoxicity of NK cells. B-cell-derived lymphoblastic-like cell line (LCL) derived exosomes transfer to CD4+ T cell and induce CD4+ T cell apoptosis. Back arrows: promotion; red “T” arrows: inhibition.
Figure 2.
The role of stroma-derived exosomes in tumor progression. (A) CAFs-derived exosomes rich in miRNA (miR-382-5p, miR-196a) transferred to tumor cells promote tumor cells migration and invasion, and induce therapeutic resistance. CAFs-derived exosomes miRNA (miR-34a-5p), protein (MFAP5) transfer to tumor cells to facilitate tumor cell proliferation. (B) TAMs-derived exosomes miRNA (miR-21) transfer to tumor cells and raise the survival and proliferation of tumor cells with induced therapeutic resistance. TAMs-derived exosomes protein (ADAM15), inhibited tumor proliferation. (C) T cell-derived exosomes promote the metastasis of tumor cells by inducing EMT. TDEs NAP1 enhance the cytotoxicity of NK cells. B-cell-derived lymphoblastic-like cell line (LCL) derived exosomes transfer to CD4+ T cell and induce CD4+ T cell apoptosis. Back arrows: promotion; red “T” arrows: inhibition.


{kind=link}
{kind=link}
Table 1.
Biological characteristics of exosomes in HNSCC.
| Cancer Type | Exosomal Cargo | Concentration | Diameter | TEM Images | Labeling | Donor Cells | Recipient Cells | Mechanism | Activated Pathway | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| Epithelium-derived exosomes | ||||||||||
| HNC | CD39/CD73 | N/A | N/A | Round | N/A | UMSCC47 | Macrophage | Promote angiogenesis | A2BR | [25] |
| HNSCC | NAP1 | N/A | 103.1 nm | Oval | N/A | CAL 27, SCC25 | NK cells | Enhances cytotoxicity of NKcells | IRF-3 | [26] |
| HNSCC | HSP90 | 50 µg/mL | 70–150 nm | Round | N/A | OSCC cell lines | RT7 | Initiate EMT | N/A | [27] |
| HNSCC | Purine metabolites | N/A | 80–150 nm | Spherical | N/A | UMSCC47 cells | N/A | Facilitate immune escape | N/A | [28] |
| NPC | MMP13 | N/A | N/A | Round | PKH67 | CNE-2 | Normoxic CNE2 cells | Enhance metastases | N/A | [29] |
| NPC | miR-20a-5p | N/A | 30–100 nm | Rounded | N/A | NPCTW03 cells | T-cell | Enhance migration and invasion | MARK | [30] |
| NPC | miR-17-5p | N/A | N/A | Spherical | PKH67 | CNE-2 | HUVECs | Promote angiogenesis | AKT/VEGF-A | [31] |
| OSCC | THBS1 | N/A | 103 nm | Oval | N/A | SCC25, CAL27 | Macrophage | Promote migration | p38, Akt, SAPK/JNK | [32] |
| OSCC | HSP90 | N/A | 80–111 nm | Round | N/A | HSC-3, HSC-3-M330 | HSC-3-M330 | Decline metastasis | N/A | [24] |
| OSCC | PD-L1, FasL | N/A | 30–150 nm | Round | N/A | SCCVII, SCC90 | T lymphocytes | Promote carcinogenesis | N/A | [33] |
| OSCC | miR-1246 | N/A | 30–100 nm | Round | PKH26 | HOC313-LM | HOC313-P | Increase migration and invasion | ERK, AKT | [34] |
| OSCC | miR-210-3p | N/A | 30–120 nm | Round | N/A | CAL27 | HUVECs | Promote angiogenesis | PI3K/AKT | [35] |
| OSCC | miR-21 | 10 μg/mL | 50–200 nm | Round | PKH67 | OSCC cells | OSCC cells | Increase metastasis | N/A | [36] |
| OSCC | miR-21 | N/A | 30–160 nm | Round | DiI | HSC-3-R | OSCC cells | Enhanced drug resistance | DNA damage | [37] |
| Stromal-derived exosomes | ||||||||||
| HNC | miR-196a | N/A | 100 mm | Cup shape | DiO | CAFs | CAL 27, SCC-25, HN4 | Promotes cisplatin resistance | N/A | [38] |
| HNC | miR-3188 | N/A | 30–150 nm | Spherical | N/A | CAFs | HN4 | Inhibits tumor growth | N/A | [39] |
| OSCC | miR-34a-5p | N/A | 40–120 nm | Oval | N/A | CAFs | CAL27, SCC15 | Promote proliferation metastasis | AKT/GSK-3β/β-catenin | [40] |
| OSCC | LncRNA LBX1-AS1 | N/A | N/A | Round | N/A | Macrophage | SCC-4, CAL-27 | Inhibits tumor growth | FOXO3, LBX1-AS1 | [41] |
HNC: head and neck cancer. NPC: nasopharyngeal carcinoma. HNSCC: head and neck squamous cell carcinoma. OSCC: oral squamous cell carcinoma. ESCC: esophageal squamous cell carcinoma. CAFs: cancer-associated fibroblasts. HSP90: heat shock protein90. MMP13: matrix metalloproteinase-13. HUVECs: human umbilical vein endothelial cells. HOC313-P: HOC313 parent cell line. N/A: not available.
Table 2.
Theranostic applications of exosomes in HNSCC.
| Tumors | Molecules | Cargo | Source | Samples | Isolation Methods | Exosomes Confirmed | Change and Function in the Diseases | Indication | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| HNC | Protein | CD39 | Plasma | 38 | mini-SEC | WB, TEM | Suppressed lymphocyte functions | Cancer biomarkers | [73] |
| HNSCC | Protein | CD3 | Plasma | 43 | mini-SEC | SEM | Varied during and post ipilimumab therapy | Cancer biomarkers | [81] |
| HNSCC | Protein | CD45(−) | Plasma | 41 | mini-SEC | WB, TEM | Higher in high stage compared to low stage | Diagnosis | [82] |
| HNSCC | Protein | PD-L1 | Plasma | 17 | mini-SEC | WB, TEM | Decreased in therapy and increase at recurrence | Cancer biomarkers | [83] |
| HNSCC | Protein | CD44v3+ | Plasma | 44 | mini-SEC | WB, TEM | Elevated in stage III/IV and metastases | Cancer biomarkers | [84] |
| HNSCC | Protein | LOXL2 | Serum | 36 | Ultracentrifugation | WB | Correlated with low-grade HNSCC | Therapy target | [85] |
| HNSCC | miRNA | miR-486-5p miR-486-3p | Saliva | 5 | Ultracentrifugation | EB, TEM | Elevated in saliva of cancer | Diagnosis | [76] |
| HPVOPC | Protein | MUC16 SIRPA | Sera | 7 | ExoQuickULTRA EV IsolationKit | WB | Augmented invasion and EMT | Diagnosis | [77] |
| NPC | Protein | BARF1 | Serum | 50 | Ultracentrifugation | WB | Increased in young NPC patients | Diagnosis | [86] |
| OSCC | miRNA | miR-24-3p | Saliva | 49 | ExoQuick-TC kit | WB | Maintain the proliferation of OSCC | Diagnosis | [80] |
| OSCC | miRNA | miR-21 | Plasma | 108 | ExoQuick-TC kit | WB, TEM | Increased migration and invasion of OSCC | Therapy target | [36] |
| OSCC | miRNA | miR-130a | Plasma | 184 | ExoQuick ULTRA EV IsolationKit | N/A | Increased and associated with poor prognosis | Diagnosis Prognosis | [87] |
| OSCC | Protein | PF4V1 CXCL7 F13A1 ApoA1 | Serum | 20 | ExoQuick-TC kit | WB, SEM | Related to the metastasis of OSCC | Diagnosis | [78] |
| OSCC | miRNA | miR-412-3p miR-512-3p | Saliva | 21 | Ultracentrifugation | WB, TEM | Up-regulated in cancer | Diagnosis | [88] |
HPVOPC: Human papilloma virus-16 (HPV-16) associated oropharyngeal cancer. HNSCC: head and neck carcinoma cells. OSCC: oral squamous cell carcinoma. NPC: nasopharyngeal carcinoma. LSCC: laryngeal squamous cell carcinoma. mini-SEC: mini size-exclusion chromatography. SEM: scanning electron microscope. TEM: transmission electron microscope. LOXL2: lysyl oxidase like 2. WB: western blot. N/A: not available.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Gao, S.; Hu, Q.; Wu, F. Functional Properties of Cancer Epithelium and Stroma-Derived Exosomes in Head and Neck Squamous Cell Carcinoma. Life 2022, 12, 757. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050757
AMA Style
Li Y, Gao S, Hu Q, Wu F. Functional Properties of Cancer Epithelium and Stroma-Derived Exosomes in Head and Neck Squamous Cell Carcinoma. Life. 2022; 12(5):757. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050757
Chicago/Turabian StyleLi, Yang, Shengtao Gao, Qi Hu, and Fanglong Wu. 2022. "Functional Properties of Cancer Epithelium and Stroma-Derived Exosomes in Head and Neck Squamous Cell Carcinoma" Life 12, no. 5: 757. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050757
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.