
The Influences of Omega-3 Polyunsaturated Fatty Acids on the Development of Skin Cancers

Department of Dermatology, University of Occupational and Environmental Health, 1-1, Iseigaoka, Yahatanishi-Ku, Kitakyushu 807-8555, Japan
*
Author to whom correspondence should be addressed.
Diagnostics 2021, 11(11), 2149; https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11112149
Submission received: 5 November 2021
/
Revised: 16 November 2021
/
Accepted: 18 November 2021
/
Published: 19 November 2021
(This article belongs to the Section Pathology and Molecular Diagnostics)
Abstract
:Dietary nutrition intake is essential for human beings and influences various physiological and pathological actions in the human body. Among various nutritional factors, dietary intake of omega-3 polyunsaturated fatty acids (PUFAs) has been shown to have various beneficial effects against inflammatory diseases. In addition to their therapeutic potency against inflammation, omega-3 PUFAs have also been shown to have anti-tumor effects via various mechanisms, such as cell arrest and apoptosis. To date, limited information is available on these effects in cutaneous malignancies. In this review, we focused on the effect of omega-3 PUFAs on skin cancers, especially malignant melanoma, basal cell carcinoma, lymphoma, and squamous cell carcinoma and discussed the detailed molecular mechanism of the omega-3 PUFA-mediated anti-tumor response. We also explored the molecular mechanisms mediated by epigenetic modifications, cell adhesion molecules, and anti-tumor immune responses.
1. Introduction
The skin is a representative peripheral lymphoid tissue and is the outermost organ of the human body [1,2,3]. It is assumed that the skin is one of the sites most exposed to environmental factors [4,5,6], which occasionally triggers skin oncogenesis. Malignant tumors are derived from normal host body cells and often show unfavorable behaviors, such as invasion and metastasis, leading to fatal clinical outcomes. As the incidence of skin cancers has gradually increased worldwide, clinicians should focus on the prevention of cutaneous malignancies.
Nutrition from food is essential for sustaining lives and maintaining body structures in animals. Among various nutrient factors, fatty acids are a component of the cell membrane and contribute to signal network transduction in the body. Although there are many types of fatty acids with various physiological and pathological actions [7,8,9,10], omega-3 fatty acids, found in fish oil and nuts, have beneficial effects against human diseases and inflammatory responses [11]. In addition to their therapeutic potency [12,13,14], omega-3 fatty acids may also play a role in the development of cutaneous malignancies due to their anti-inflammatory actions [15]. In this review, we have focused on the impact of omega-3 fatty acids on cutaneous malignancies and discussed the detailed molecular mechanisms in addition to recent novel findings.
2. Omega-3 Fatty Acids
Omega-3 polyunsaturated fatty acids (PUFAs) are found in fish oil and nuts. These fatty acids consist of ≥18 carbon chains, including a double bond of three atoms that are away from the terminal methyl group. Omega-3 PUFAs have been reported including docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA). ALA is first converted to EPA and subsequently to DHA [11]. The liver is primarily responsible for these conversions. However, conversion enzymes are extremely limited in the human body [11]. Therefore, to obtain their benefits, the direct intake of DHA and EPA from food and supplements is necessary.
The omega-3 PUFAs are essential nutrients and cannot be synthesized de novo in sufficient quantities for normal physiological function [11]. α-linolenic fatty acid is an omega-3 PUFA member and a precursor of other omega-3 PUFAs mediated by elongase and desaturase activities. In addition, α-linolenic acid is an essential fatty acid because humans do not possess the enzymes to synthesize the compound; therefore, it must be obtained from dietary sources. α-linolenic fatty acid is first metabolized by ∆6-desaturase and subsequently elongase, leading to the conversion into EPA following ∆5-desaturase activity. Further elongase and ∆4-desaturase or ∆6-desaturase activity finally produce DHA [16].
Omega-3 PUFAs have anti-inflammatory effects against inflammatory diseases, including inflammatory bowel disease, psoriasis, and rheumatoid arthritis [11,17]. The metabolites of omega-3 PUFAs, such as resolvins, also demonstrate a strong anti-inflammatory potential. E-series resolvins are derived from EPA via the acetylated cyclooxygenase-2 or cytochrome P450 pathways. In contrast, D-series resolvins and protectins are converted from DHA by 15-lipoxygenase. Their metabolites have shown strong anti-inflammatory effects in various skin disease models [12,13,14]. These metabolites have also demonstrated anti-inflammatory effects in inflammatory disease models, such as psoriasis, contact hypersensitivity, asthma, and colitis. However, limited information is available on their role in malignancies, especially skin cancers.
3. Influence of Omega-3 Fatty Acids on Skin Cancers
Various malignancies are influenced by omega-3 PUFAs. In this section, we detail the effects of omega-3 PUFAs on skin cancers, especially malignant melanoma, squamous cell carcinoma, basal cell carcinoma, and lymphoma.
3.1. Malignant Melanoma
Melanoma is a melanocyte-derived malignancy with an unfavorable clinical course without radical treatment, as it is resistant to current chemotherapies [18,19]. Although current immune checkpoint inhibitor treatment and BRAF-targeted agents have dramatically expanded the therapeutic options and improved clinical outcomes, clinical outcomes are unsatisfactory. Therefore, some additional therapeutic options for the treatment of malignant melanoma are highly desirable.
The BRAF gene is located on chromosome 7 (7q34). It encodes the BRAF protein [20] and plays a role in the mitogen-activated protein kinase (MAPK) pathway activation [21]. This activation regulates the development of tumor cells, including apoptosis, cell growth, and proliferation (Figure 1). BRAF gene mutations impair these functions. The BRAFV600 mutations are a representative gene mutation in melanoma and are detected in approximately 50% of patients with malignant melanoma. This gene mutation causes activation of the MAPK downstream pathway [22]. The second most frequent mutation in malignant melanoma is NRAS, which is related to the regulation of PI3K and MAPK [23]. In acral and mucosal melanoma, C-KIT plays an important role in the malignant process [24]. Currently, there are several therapeutic options for advanced malignant melanoma. For example, immunotherapy against melanoma is currently being developed, including PD-1-or CTLA-4-targeted therapy [25]. Moreover, BRAF-targeted therapy may add another strategy for improving clinical outcomes in patients with BRAF-mutated melanoma.
As the risk factor of malignant melanoma, ultraviolet light exposure is one of the representative oncogenesis factors. UV radiation induces dysregulation of p53 in melanocytes, which is known to be a tumor suppressor gene, and contributes to the suppression of tumor apoptosis [26].
Several studies have reported that a decreased risk of melanoma is related to the intake of omega-3 PUFAs [27]. In one study, the intake of EPA or DHA was associated with an approximately 80% lower risk of malignant melanoma [27]. To optimize the benefit of omega-3 PUFAs, DHA was used in combination with chemotherapy. DHA plus doxorubicin has shown greater anti-tumor efficacy than doxorubicin alone in melanoma cells [28]. In one study, when DHA–paclitaxel was administered to patients with cutaneous and mucosal metastatic melanoma, 10% of patients had partial responses and 50% of patients had stable disease [29]. In another study, the efficacy of DHA–paclitaxel was demonstrated in patients with metastatic uveal melanoma. Among patients with liver metastases, 4.5% had a partial response lasting for 5 months and 32% had stable disease [30]. Dacarbazine is currently used for melanoma treatment. In a study of 393 chemotherapy-naive patients with metastatic melanoma, DHA–paclitaxel was found to be not inferior to dacarbazine [31].
Linoleic acid, EPA, and DHA can suppress murine melanoma cell growth [32]. Intravenously injected malignant murine melanoma cells cultured with EPA have shown impaired tumor invasion and metastasis. EPA inhibits the lipoxygenase pathway, which is required for tumor metastasis [33]. COX-2 is essential for tumor invasion. TNF-α upregulates COX-2 expression in melanoma cells, which is decreased in melanoma cells co-cultured with EPA or DHA, subsequently decreasing tumor invasion [34]. DHA also decreases the expression and stability of COX-2 mRNA [35], while CXCR4 contributes to the progression of melanoma. In contrast, EPA decreases CXCR4 levels [36]. DHA inhibits cell growth and reduces invasion and migration by inhibiting matrix metalloproteinases [37]. Furthermore, algal oil, which is rich in omega-3 PUFAs, has been shown to suppress lung metastasis of B16F10 melanoma by an autophagy mechanism mediated by inactivation of the p38 MAPK and mammalian target of rapamycin (mTOR) and activation of c-Jun N-terminal kinases (JNKs), which leads to the suppression of proinflammatory cytokine production [38].
As the detailed molecular mechanism of the anti-cancer effects of omega-3 PUFAs, the suppression of debris-induced tumor growth activation has been elucidated [39]. DHA and EPA induce the release of calcium and reactive oxygen species (ROS) production, which induces the ER stress-triggered apoptosis pathway and tumor suppressor gene induction, such as P53 [40]. In one study, decreased proliferation associated with oxidative protein and DNA damage was observed in EPA- and DHA-treated melanoma cells [41].
Although the synthase for omega-3 PUFA is limited in the human body, Fat-1 can convert omega-6 to omega-3 fatty acids. In transgenic mice, this showed a reduction in tumor growth. In vitro experiments have shown that the addition of EPA or PGE3 inhibited the growth of melanoma cells [42]. Treatment with omega-3-rich fish oil has also been shown to reduce melanoma development and leukotriene B4 (LTB4) production [43]. DHA inhibits cell growth in addition to the expression of cyclins and cyclin-dependent kinase inhibitors and the expression of pRb. DHA treatment results in an increase in hypophosphorylated Rb [44]. DHA also suppresses human melanoma cell growth in vivo. In animals, treatment with an omega-3 PUFA-enriched diet has demonstrated effectiveness in reducing tumors (69% smaller in weight of the tumor and 76% smaller in volume) compared to treatment with an omega-6 fatty acid-enriched diet [45].
Omega-3 PUFAs enhance the efficacy of anti-cancer agents, such as cisplatin (CDDP). EPA- and DHA-treated melanoma cells are sensitive to CDDP-induced suppression of tumor growth and invasion. Omega-3 PUFAs also drive the expression of ERCC1, DUSP6, and p-ERK, which activates ERCC1 and thereby improves the anti-tumor action of CDDP against melanoma cells [46].
Connexin 43 is related to chemosensitivity. It is suppressed in the tumor microenvironment. EPA improves chemosensitivity in murine melanoma by inducing connexin 43 expression, which is regulated by MAPK pathways [47]. In addition, DHA enhances the sensitivity of various tumor cells to ROS-inducing anti-cancer agents [48].
In contrast, another study reported an opposite effect. Fish oil EPA-28 enriched in EPA and DHA increases tumor growth and metastasis of subcutaneously injected B16 melanoma cells [49]. EPA-28 suppresses CD4+ T cell infiltration around the tumor in addition to the cytolytic activity of T cells and macrophages. However, the inflammatory tumor microenvironment drives tumor development. Debris derived from tumor cells in chemotherapy stimulate primary tumor growth, which is inhibited by anti-inflammatory agents that are known to have anti-tumor effects. For example, resolvin D1 (RvD1), RvD2, or RvE1 inhibits debris-stimulated cancer progression by enhancing the clearance of debris through macrophage phagocytosis in various types of tumors and suppresses inflammatory cytokines/chemokines by tumor cell debris-triggered human macrophages [50]. Overall, omega-3 PUFAs are expected to suppress melanoma development. However, omega-3 PUFAs may be disadvantageous in certain conditions in which anti-tumor immune responses to melanoma are expected.
3.2. Basal Cell Carcinoma and Omega-3 Fatty Acids
Basal cell carcinoma is a malignancy derived from epidermal basal keratinocytes and is commonly observed in sun-exposed sites. The clinical behavior is basically slow progressive local site invasion. Distant metastasis is rarely observed. Although surgical treatment is the gold standard for limited localized basal cell carcinoma, there is currently no therapeutic option for distant metastasis of invaded and unresectable basal cell carcinoma. Therefore, prevention is important.
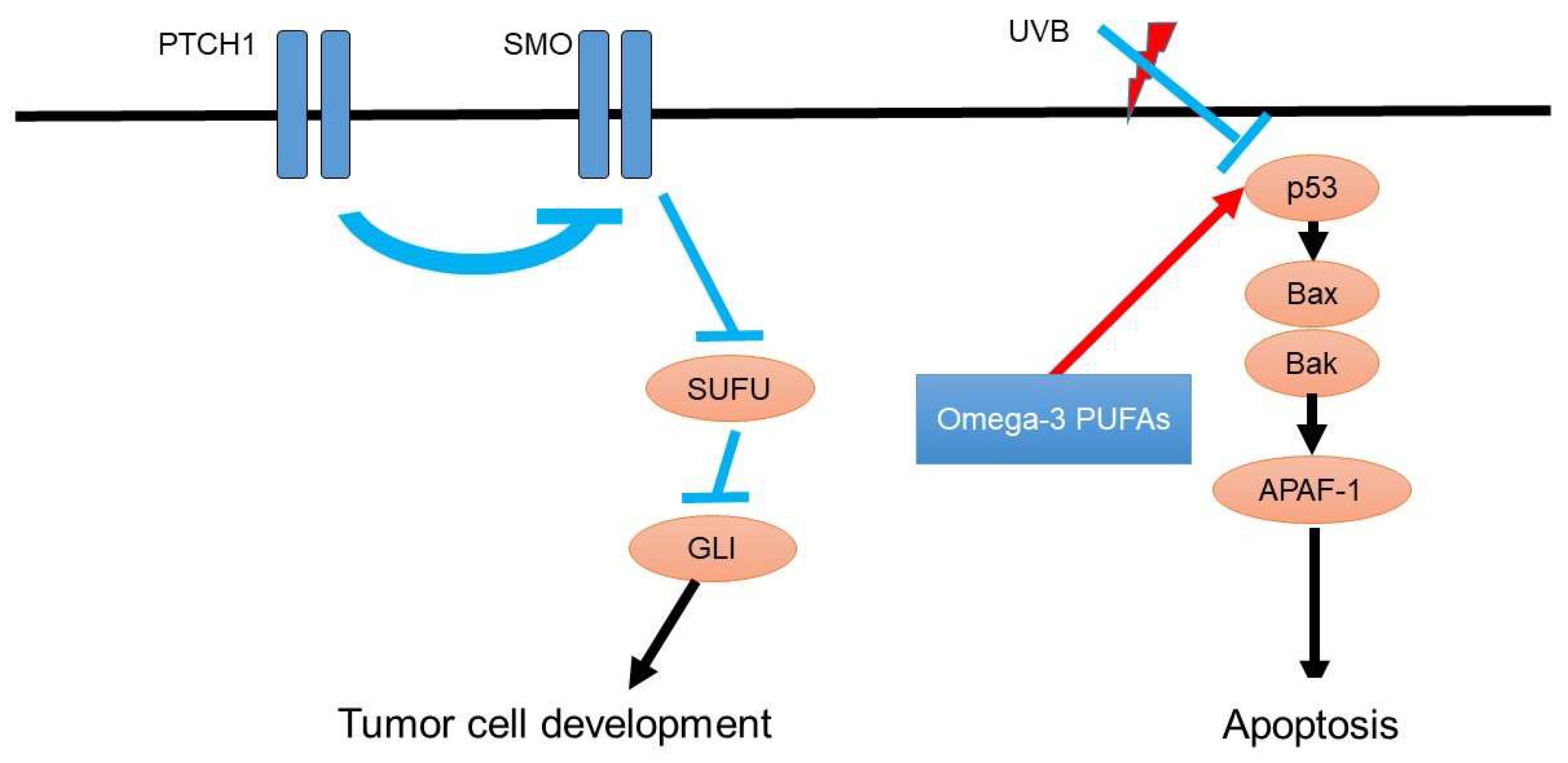
As the detailed oncogenesis of basal cell carcinoma, the hedgehog pathway gene mutation activates the patched homolog and smoothened homolog (Figure 2). Consistently, inhibition of this hedgehog signaling pathway suppresses the metastasis of basal cell carcinoma [51]. In addition to the hedgehog pathway, p53 mutations are also involved in the development of basal cell carcinoma [52]. For instance, ultraviolet light exposure induces p53 mutations and leads to tumorigenesis of basal cell carcinoma [53].
Ultraviolet light exposure also enhances the risk of basal cell carcinoma and downregulates p53 expression in the basal layer of the epidermal keratinocyte, leading to the contribution of the development of basal cell carcinoma [54].
A limited number of studies have focused on the efficacy of omega-3 PUFAs. One epidemiological study suggests that there is no clear evidence of an association between the intake of omega-3 fatty acids and the risk of basal cell carcinoma. Intake of linolenic acid was also negatively related to the risk of basal cell carcinoma [55]. Conversely, a Mendelian randomization study using PUFA level genome-wide association studies showed that alpha-linolenic acid and linoleic acid are associated with a lower risk of basal cell carcinoma [56].
3.3. ATLL and Omega-3 Fatty Acids
Adult T-cell leukemia/lymphoma is a malignancy related to human T-cell lymphotropic virus type I (HTLV-1)-infected mature CD4+ T cells [57,58]. Adult T-cell leukemia/lymphoma is classified into four clinical groups according to Shimoyama’s classification based on the severity, the number of abnormal lymphocytes, and organ involvement [58]. Skin lesions are observed in approximately 50% of adult T-cell leukemia/lymphoma patients. Assessment of skin lesions helps predict prognosis [59,60,61]. Although aggressive types—the acute and lymphoma types—of adult T-cell leukemia/lymphoma tend to have an unfavorable clinical course [62,63,64], the chronic and smoldering types are generally indolent and may be managed with “watchful waiting” [65].
A limited number of studies have focused on the therapeutic potential of ATLL. In one study, the combination of DHA and arsenic trioxide, interferon-alpha, and emodin showed a potent synergistic effect on cell cycle arrest and cell death in HTLV-1 infected cells [66]. This finding suggests that DHA is a possible candidate for the treatment of ATLL to obtain additional therapeutic effects.
3.4. Diffuse Large B-Cell Lymphoma and Omega-3 Fatty Acids
Diffuse large B-cell lymphoma is a malignancy of the B cells. It is a common type of non-Hodgkin cutaneous lymphoma [67]. Diffuse large B-cell lymphoma is commonly observed in the extremities and clinically manifests as solid nodules or tumors in the skin.
Dietary products with higher fat contents are positively associated with the risk of non-Hodgkin lymphoma, whereas a high intake of omega-3 fatty acids and seafood is inversely associated with the risk [68]. A low intake of omega-3 fatty acids is associated with unfavorable survival in patients with diffuse large B-cell lymphoma [69].
3.5. Cutaneous Squamous Cell Carcinoma (SCC)
Cutaneous SCC is a keratinocyte-derived cutaneous malignancy of the skin. Due to its increasing incidence worldwide, cutaneous SCC is a hot topic among clinicians [70,71]. Although SCC limited to the skin can be treated with surgical resection, cutaneous SCC with distant metastasis exhibit an unfavorable clinical course due to a limited number of effective therapeutic options [72,73].
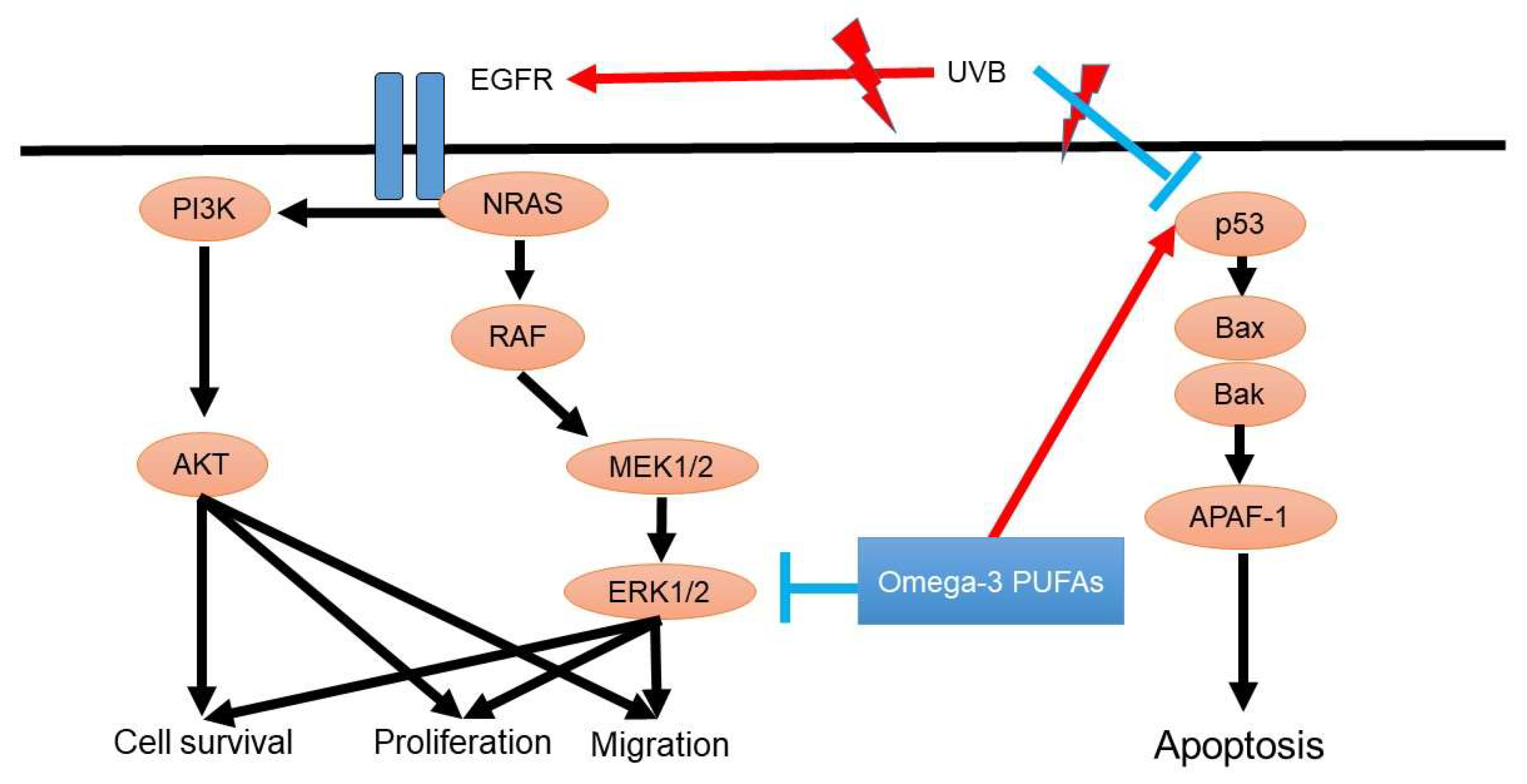
The p53 tumor suppressor gene is associated with oncogenesis in cutaneous SCC. P53 is crucial for the development of cutaneous SCC because p53 is responsible for cell apoptosis, DNA repair, and cell cycle arrest. In addition, EGFR-mediated PI3K/AKT signaling and NRAS/RAF/MEK/ERK signaling also contribute to the development of SCC (Figure 3).
PUFAs show an inhibitory effect on prostaglandin production by squamous carcinoma cells [74], which plays a role in the development of SCC. Extracellular signal-regulated kinase promotes cell apoptosis after omega-3 PUFA treatment. DHA promotes ROS production and activates c-Jun N-terminal kinase, leading to the enhancement of toxicity to normal keratinocytes [75]. RvD2 significantly reduces tumor size and cancer-derived cytokines/chemokines (TNF-α, IL-6, CXCL10, and MCP-1) [76]. EPA inhibits cell division [77] by releasing Ca2+ from intracellular stores, leading to the activation of protein kinase R, which causes cell cycle arrest in G1. Oral administration of EPA reduces tumor size by decreasing the expression of cyclin D1 in tumors. Omega-3 PUFAs inhibit tumor growth by combining with ionizing radiation [78]. Omega-3 PUFAs suppress angiogenesis, tumor proliferation, and cyclooxygenase-2 production.
Cancer-associated weight loss is an important issue in the treatment of malignancies. EPA has been shown to be potent in suppressing this response [79]. Seventy percent of the subjects who consumed EPA-containing food maintained or gained body weight. An EPA-containing nutritional supplement may also increase or maintain the body weight of patients with SCC-associated body weight loss.
Another randomized, single-blind, placebo-controlled clinical trial was conducted in patients with SCC to confirm the efficacy of EPA during treatment. In this study, EPA-supplemented diets decreased inflammatory cytokines and maintained body weight [80].
Furthermore, EPA may suppress the migration of SCC cells and reduce migration velocity and directionality in a dose-dependent manner [81].
4. Summary of the Effect of Omega-3 Fatty Acids against Each Skin Cancer
The detailed direct effects of omega-3 fatty acids against skin cancers are summarized and shown in Table 1. Epidemiological analyses have shown the beneficial potency of omega-3 PUFAs in melanoma [27], basal cell carcinoma [56], and diffuse large B cell lymphoma [69]. In addition, omega-3 PUFAs suppress tumor growth in melanoma [32,37,41,42,44,45] and squamous cell carcinoma [77], inhibiting migration and invasion in melanoma [33,34,37] and squamous cell carcinoma [81]. Omega-3 PUFAs also enhance the sensitivity to chemotherapy in melanoma [28,29,30,31,46,47,48] and radiosensitivity in squamous cell carcinoma [78]. Omega-3 PUFAs metabolites RvD1, RvD2, and RvE1 suppress debris-stimulated cancer progression in melanoma [50], and RvD2 reduces tumor size and cancer-derived cytokines/chemokines [76].
5. Epigenetic Modification by Omega-3 Fatty Acids
Normally, DNA is considered a single-line construction. However, DNA does not exist as a single line. DNA in the body is entwined with a DNA-binding protein called histone. Furthermore, it is packed into histones. When the bond between DNA and histone is loosened, gene transduction begins to read the DNA information. In vivo, the histone or DNA chemical modification-mediated epigenetic mechanism is a powerful regulatory mechanism of gene transduction [82,83]. These epigenetic modifications are involved in various human diseases [82,83,84]. Recent studies have reported that bacteria-derived short-chain fatty acids can modify histone acetylation by suppressing HDACs, creating open chromatin sites and inducing gene transduction [83].
A recent study demonstrated that omega-3 fatty acids may have a role in epigenetic modification, which in turn, has significant multiple anti-tumor roles [85]. There is an interaction between the anti-tumor role of omega-3 PUFAs and the DNA demethylation pathway leading to the anti-cancer mechanism of omega-3 PUFAs mediated by altering the patterns of ten-eleven translocation (TET) 1 expression.
TET1 is also involved in cancers such as basal cell carcinoma [86], SCC [87], melanoma [88], and lymphomas [89]. UV radiation is involved in the pathogenesis of tumor development related to TET1. In the normal skin and cutaneous cell lines, UVB exposure induces the upregulation of DNMT1 and downregulates TETs [87]. In mouse models, silencing of DNMT1 and overexpression of TET1 and TET2 can lead to an increase in ID4 expression [87], which in turn, reduces cell proliferation, migration, and invasion, increases apoptosis in cutaneous SCC cell lines, and reduces tumorigenesis [87]. TET1 is also downregulated on NF-κB activation through the binding of p65 to its consensus sequence in the TET1 promoter [88].
6. P53 and Omega-3 PUFAs
p53 is a classic tumor suppressor gene that is responsible for the induction of cell apoptosis and cell cycle arrest. p53 is involved in tumorigenesis and the development of tumors. There are various studies regarding the efficacy of omega-3 PUFAs on p53 suppression, such as in colorectal adenocarcinoma [90,91]. Although the effect of omega-3 PUFAs on p53 expression in skin cancers is unclear, further investigation to clarify the actual impact of omega-3 PUFAs should be considered.
7. ERK Inhibition and Omega-3 PUFAs
The MEK–ERK–BRAF pathway has been highlighted by clinicians because the efficacy of BRAF and/or BRAF-related pathway inhibition dramatically improves anti-tumor effects. RAS is a non-BRAF oncogene activated in malignant melanoma and enhances tumor functions, such as cell growth and apoptosis. RAS promotes the downstream pathways of MEK/ERK, MAPK, and PI3K/AKT signaling. However, BRAF mutation in melanoma, especially BRAFV600 mutations, activates the downstream pathway of MAPK and positively drives the development of malignant melanoma.
Several studies have reported the anti-tumor effect of omega-3 PUFAs against malignancies, but not in skin cancers. Omega-3 PUFAs have been shown to suppress tumor growth mediated by ERK suppression in retinoblastoma [92], leukemia cells [93], breast cancer [94], and colon cancer [95]. The effect of omega-3 PUFAs on melanoma cells is unclear. Therefore, the detailed molecular mechanism of the anti-tumor effect of omega-3 PUFAs on melanoma should be investigated.
8. Adhesion Molecule and Omega-3 PUFAs
Cell–cell adhesion is important in maintaining the physiology of cell function. Its abnormality changes the structure and enhances the dissemination and metastatic spread of tumor cells. A component of the adhesion molecule is E-cadherin, and cell adhesion molecule 1 (CADM1) plays a role in cell–cell adhesion, and low expression of these adhesion molecules is related to lower survival in solid tumors. Cell adhesion is essential to the tumor environment. Reduced expression of the cell adhesion molecules plays a pivotal role in the metastatic ability of the tumor [96,97,98]. Although there is no clear evidence demonstrating that omega-3 PUFAs modify the expression of these cell adhesion molecules in skin cancers, EPA increases the expression of E-cadherin in colon cancer and breast cancer [99].
In addition to cell–cell adhesion, vascular cell adhesion molecules also play an important role in the distant metastasis of the tumor. High expression of vascular cell adhesion molecules becomes a scaffold for cell invasion into the blood vessels and enhances the migration of tumor cells. There is no report regarding the effect of omega-3 PUFA on the vascular cell adhesion molecules focused on the skin cancers-bearing host body. However, vascular cell adhesion molecule 1 expression on ACL-15 cells is downregulated by EPA–ethyl ester treatment [100], suggesting that a decreased ability of tumor cell adhesion to the capillary bed is expected during the treatment of omega-3 PUFA in patients with skin cancers.
9. Anti-Tumor Immunity and Omega-3 PUFAs
Immune checkpoint inhibitor treatment currently improves clinical outcomes in patients with unresectable skin cancers with distant metastasis [101,102,103]. Although there are several adverse reactions [104], some additive therapeutic potential is desired with the combination of immune checkpoint inhibitor treatment [105]. Although omega-3 PUFAs show an anti-tumor effect in skin cancers, it may be disadvantageous to use omega-3 PUFAs in combination therapy with immune checkpoint inhibitor treatment.
PUFAs regulate T-cell activation [106]. After culture with DHA, DCs expressed higher levels of CD40, CD80, CD83, CD86, and PDL-1. However, fewer T cells co-cultured with these DCs proliferated, while an increased Treg phenotype was observed in co-cultures with DHA-primed DCs.
10. Conclusions
Omega-3 PUFAs influence the proliferation and metastasis of various skin cancers Basically, omega-3 PUFAs show beneficial potency against skin cancers. Therefore, the intake of omega-3 PUFAs may be recommended. However, omega-3 PUFAs suppress the immune reaction, mediated mainly by Tregs, possibly leading to the impairment of anti-tumor immune reactions. Based on this mechanism, a combination of omega-3 PUFAs and an immune checkpoint inhibitor may not be recommended for use in the treatment of malignancies. However, specialized lipid mediators, such as resolvins and protectins, potently suppress tumor-derived dust-induced inflammatory responses in the induction of tumor development. Thus, omega-3 PUFAs may positively drive the anti-tumor effect in skin cancers.
Author Contributions
Y.M., Y.S. and M.N. wrote and revised manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Y.S. received basic research grant from Japanese Dermatology Association (Novartis Pharma donated) and research grant from Lydia O’Leary Memorial Pias Dermatological Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Egawa, G.; Kabashima, K. Skin as a peripheral lymphoid organ: Revisiting the concept of skin-associated lymphoid tissues. J. Investig. Dermatol. 2011, 131, 2178–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef]
- Vasseur, P.; Pohin, M.; Jégou, J.F.; Favot, L.; Venisse, N.; McHeik, J.; Morel, F.; Lecron, J.C.; Silvain, C. Liver fibrosis is associated with cutaneous inflammation in the imiquimod-induced murine model of psoriasiform dermatitis. Br. J. Dermatol. 2018, 179, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Zhang, M.; Zhang, C.; Sang, H. Psoriasis-Like Inflammation Induced Renal Dysfunction through the TLR/NF-κB Signal Pathway. Biomed. Res. Int. 2020, 2020, 3535264. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Nakamura, M.; Tokura, Y. Generalized fixed drug eruption caused by pazufloxacin. Acta Derm.-Venereol. 2011, 91, 600–601. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Nakajima, S.; Nonomura, Y.; Otsuka, A.; Egawa, G.; Yoshimoto, T.; Nakamura, M.; Narumiya, S.; et al. Prostaglandin E(2) (PGE(2))-EP2 signaling negatively regulates murine atopic dermatitis-like skin inflammation by suppressing thymic stromal lymphopoietin expression. J. Allergy Clin. Immunol. 2019, 144, 1265–1273.e1269. [Google Scholar] [CrossRef] [Green Version]
- Ueharaguchi, Y.; Honda, T.; Kusuba, N.; Hanakawa, S.; Adachi, A.; Sawada, Y.; Otsuka, A.; Kitoh, A.; Dainichi, T.; Egawa, G.; et al. Thromboxane A(2) facilitates IL-17A production from Vγ4(+) γδ T cells and promotes psoriatic dermatitis in mice. J. Allergy Clin. Immunol. 2018, 142, 680–683.e2. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Saito-Sasaki, N.; Mashima, E.; Nakamura, M. Daily Lifestyle and Inflammatory Skin Diseases. Int. J. Mol. Sci. 2021, 22, 5204. [Google Scholar] [CrossRef]
- Wada, A.; Sawada, Y.; Sugino, H.; Nakamura, M. Angioedema and Fatty Acids. Int. J. Mol. Sci. 2021, 22, 9000. [Google Scholar] [CrossRef]
- Sawada, Y.; Saito-Sasaki, N.; Nakamura, M. Omega 3 Fatty Acid and Skin Diseases. Front. Immunol. 2020, 11, 623052. [Google Scholar] [CrossRef]
- Saito-Sasaki, N.; Sawada, Y.; Mashima, E.; Yamaguchi, T.; Ohmori, S.; Yoshioka, H.; Haruyama, S.; Okada, E.; Nakamura, M. Maresin-1 suppresses imiquimod-induced skin inflammation by regulating IL-23 receptor expression. Sci. Rep. 2018, 8, 5522. [Google Scholar] [CrossRef]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Otsuka, A.; Ogawa, N.; Kobayashi, Y.; Nakamura, M.; Kabashima, K. Resolvin E1 attenuates murine psoriatic dermatitis. Sci. Rep. 2018, 8, 11873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, Y.; Honda, T.; Hanakawa, S.; Nakamizo, S.; Murata, T.; Ueharaguchi-Tanada, Y.; Ono, S.; Amano, W.; Nakajima, S.; Egawa, G.; et al. Resolvin E1 inhibits dendritic cell migration in the skin and attenuates contact hypersensitivity responses. J. Exp. Med. 2015, 212, 1921–1930. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Nakamura, M. Daily Lifestyle and Cutaneous Malignancies. Int. J. Mol. Sci. 2021, 22, 5227. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Malau-Aduli, A.E.O.; Nichols, P.D. Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, R.W.; Fisher, D.E. Treatment of Advanced Melanoma in 2020 and Beyond. J. Investig. Dermatol. 2020, 141, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Simiczyjew, A.; Dratkiewicz, E.; Mazurkiewicz, J.; Ziętek, M.; Matkowski, R.; Nowak, D. The Influence of Tumor Microenvironment on Immune Escape of Melanoma. Int. J. Mol. Sci. 2020, 21, 8359. [Google Scholar] [CrossRef]
- Chapman, P.B.; Hauschild, A.; Robert, C.; Haanen, J.B.; Ascierto, P.; Larkin, J.; Dummer, R.; Garbe, C.; Testori, A.; Maio, M.; et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 2011, 364, 2507–2516. [Google Scholar] [CrossRef] [Green Version]
- Peyssonnaux, C.; Eychène, A. The Raf/MEK/ERK pathway: New concepts of activation. Biol. Cell 2001, 93, 53–62. [Google Scholar] [CrossRef]
- Jung, T.; Haist, M.; Kuske, M.; Grabbe, S.; Bros, M. Immunomodulatory Properties of BRAF and MEK Inhibitors Used for Melanoma Therapy-Paradoxical ERK Activation and Beyond. Int. J. Mol. Sci. 2021, 22, 9890. [Google Scholar] [CrossRef] [PubMed]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Beadling, C.; Jacobson-Dunlop, E.; Hodi, F.S.; Le, C.; Warrick, A.; Patterson, J.; Town, A.; Harlow, A.; Cruz, F., 3rd; Azar, S.; et al. KIT gene mutations and copy number in melanoma subtypes. Clin. Cancer Res. 2008, 14, 6821–6828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires da Silva, I.; Ahmed, T.; Reijers, I.L.M.; Weppler, A.M.; Betof Warner, A.; Patrinely, J.R.; Serra-Bellver, P.; Allayous, C.; Mangana, J.; Nguyen, K.; et al. Ipilimumab alone or ipilimumab plus anti-PD-1 therapy in patients with metastatic melanoma resistant to anti-PD-(L)1 monotherapy: A multicentre, retrospective, cohort study. Lancet Oncol. 2021, 22, 836–847. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, G.; Pittelkow, M.R.; Ramoni, M.; Tsao, H. Expression profiling of UVB response in melanocytes identifies a set of p53-target genes. J. Investig. Dermatol. 2006, 126, 2490–2506. [Google Scholar] [CrossRef] [Green Version]
- Donat-Vargas, C.; Berglund, M.; Glynn, A.; Wolk, A.; Åkesson, A. Dietary polychlorinated biphenyls, long-chain n-3 polyunsaturated fatty acids and incidence of malignant melanoma. Eur. J. Cancer 2017, 72, 137–143. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Jiang, W.; Yang, Z.; Zhang, Z. Synthesis and preliminary antitumor activity evaluation of a DHA and doxorubicin conjugate. Bioorganic Med. Chem. Lett. 2006, 16, 2974–2977. [Google Scholar] [CrossRef]
- Homsi, J.; Bedikian, A.Y.; Kim, K.B.; Papadopoulos, N.E.; Hwu, W.J.; Mahoney, S.L.; Hwu, P. Phase 2 open-label study of weekly docosahexaenoic acid-paclitaxel in cutaneous and mucosal metastatic melanoma patients. Melanoma Res. 2009, 19, 238–242. [Google Scholar] [CrossRef]
- Homsi, J.; Bedikian, A.Y.; Papadopoulos, N.E.; Kim, K.B.; Hwu, W.J.; Mahoney, S.L.; Hwu, P. Phase 2 open-label study of weekly docosahexaenoic acid-paclitaxel in patients with metastatic uveal melanoma. Melanoma Res. 2010, 20, 507–510. [Google Scholar] [CrossRef]
- Bedikian, A.Y.; DeConti, R.C.; Conry, R.; Agarwala, S.; Papadopoulos, N.; Kim, K.B.; Ernstoff, M. Phase 3 study of docosahexaenoic acid-paclitaxel versus dacarbazine in patients with metastatic malignant melanoma. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2011, 22, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Tezabwala, B.U.; Bennett, M.; Grundy, S.M. Immunotoxicity of polyunsaturated fatty acids in serum-free medium. Immunopharmacol. Immunotoxicol. 1995, 17, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Reich, R.; Royce, L.; Martin, G.R. Eicosapentaenoic acid reduces the invasive and metastatic activities of malignant tumor cells. Biochem. Biophys. Res. Commun. 1989, 160, 559–564. [Google Scholar] [CrossRef]
- Denkins, Y.; Kempf, D.; Ferniz, M.; Nileshwar, S.; Marchetti, D. Role of omega-3 polyunsaturated fatty acids on cyclooxygenase-2 metabolism in brain-metastatic melanoma. J. Lipid Res. 2005, 46, 1278–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serini, S.; Fasano, E.; Piccioni, E.; Monego, G.; Cittadini, A.R.; Celleno, L.; Ranelletti, F.O.; Calviello, G. DHA induces apoptosis and differentiation in human melanoma cells in vitro: Involvement of HuR-mediated COX-2 mRNA stabilization and β-catenin nuclear translocation. Carcinogenesis 2012, 33, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, C.Y.; Arita, M.; Kim, K.; Li, X.; Zhang, H.; Kang, J.X. An omega-3 polyunsaturated fatty acid derivative, 18-HEPE, protects against CXCR4-associated melanoma metastasis. Carcinogenesis 2018, 39, 1380–1388. [Google Scholar] [CrossRef]
- Serini, S.; Zinzi, A.; Vasconcelos, R.O.; Fasano, E.; Riillo, M.G.; Celleno, L.; Trombino, S.; Cassano, R.; Calviello, G. Role of β-catenin signaling in the anti-invasive effect of the omega-3 fatty acid DHA in human melanoma cells. J. Dermatol. Sci. 2016, 84, 149–159. [Google Scholar] [CrossRef]
- Tan, R.H.; Wang, F.; Fan, C.L.; Zhang, X.H.; Zhao, J.S.; Zhang, J.J.; Yang, Y.; Xi, Y.; Zou, Z.Q.; Bu, S.Z. Algal oil rich in n-3 polyunsaturated fatty acids suppresses B16F10 melanoma lung metastasis by autophagy induction. Food Funct. 2018, 9, 6179–6186. [Google Scholar] [CrossRef]
- Bachi, A.L.; Kim, F.J.; Nonogaki, S.; Carneiro, C.R.; Lopes, J.D.; Jasiulionis, M.G.; Correa, M. Leukotriene B4 creates a favorable microenvironment for murine melanoma growth. Mol. Cancer Res. 2009, 7, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Puskás, L.G.; Fehér, L.Z.; Vizler, C.; Ayaydin, F.; Rásó, E.; Molnár, E.; Magyary, I.; Kanizsai, I.; Gyuris, M.; Madácsi, R.; et al. Polyunsaturated fatty acids synergize with lipid droplet binding thalidomide analogs to induce oxidative stress in cancer cells. Lipids Health Dis. 2010, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Zajdel, A.; Wilczok, A.; Chodurek, E.; Gruchlik, A.; Dzierzewicz, Z. Polyunsaturated fatty acids inhibit melanoma cell growth in vitro. Acta Pol. Pharm. 2013, 70, 365–369. [Google Scholar] [PubMed]
- Xia, S.; Lu, Y.; Wang, J.; He, C.; Hong, S.; Serhan, C.N.; Kang, J.X. Melanoma growth is reduced in fat-1 transgenic mice: Impact of omega-6/omega-3 essential fatty acids. Proc. Natl. Acad. Sci. USA 2006, 103, 12499–12504. [Google Scholar] [CrossRef] [Green Version]
- Almeida, E.B.; Silva, K.P.H.; Paixão, V.; Amaral, J.B.D.; Rossi, M.; Xavier-Navarro, R.A.; Barros, K.V.; Silveira, V.L.F.; Vieira, R.P.; Oliveira, L.V.F.; et al. A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment. Int. J. Mol. Sci. 2019, 20, 3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albino, A.P.; Juan, G.; Traganos, F.; Reinhart, L.; Connolly, J.; Rose, D.P.; Darzynkiewicz, Z. Cell cycle arrest and apoptosis of melanoma cells by docosahexaenoic acid: Association with decreased pRb phosphorylation. Cancer Res. 2000, 60, 4139–4145. [Google Scholar] [PubMed]
- Nehra, D.; Pan, A.H.; Le, H.D.; Fallon, E.M.; Carlson, S.J.; Kalish, B.T.; Puder, M. Docosahexaenoic acid, G protein-coupled receptors, and melanoma: Is G protein-coupled receptor 40 a potential therapeutic target? J. Surg. Res. 2014, 188, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, R.O.; Serini, S.; de Souza Votto, A.P.; Trindade, G.S.; Fanali, C.; Sgambato, A.; Calviello, G. Combination of ω-3 fatty acids and cisplatin as a potential alternative strategy for personalized therapy of metastatic melanoma: An in-vitro study. Melanoma Res. 2019, 29, 270–280. [Google Scholar] [CrossRef]
- Yang, C.J.; Kuo, C.T.; Wu, L.H.; Chen, M.C.; Pangilinan, C.R.; Phacharapiyangkul, N.; Liu, W.; Chen, Y.H.; Lee, C.H. Eicosapentaenoic acids enhance chemosensitivity through connexin 43 upregulation in murine melanoma models. Int. J. Med. Sci. 2019, 16, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, M.; Sturlan, S.; Roth, E.; Wessner, B.; Bachleitner-Hofmann, T. Enhancement of arsenic trioxide-mediated apoptosis using docosahexaenoic acid in arsenic trioxide-resistant solid tumor cells. Int. J. Cancer 2004, 112, 707–712. [Google Scholar] [CrossRef]
- Salem, M.L.; Kishihara, K.; Abe, K.; Matsuzaki, G.; Nomoto, K. N-3 polyunsaturated fatty acids accentuate B16 melanoma growth and metastasis through suppression of tumoricidal function of T cells and macrophages. Anticancer Res. 2000, 20, 3195–3203. [Google Scholar]
- Sulciner, M.L.; Serhan, C.N.; Gilligan, M.M.; Mudge, D.K.; Chang, J.; Gartung, A.; Lehner, K.A.; Bielenberg, D.R.; Schmidt, B.; Dalli, J.; et al. Resolvins suppress tumor growth and enhance cancer therapy. J. Exp. Med. 2018, 215, 115–140. [Google Scholar] [CrossRef]
- Sekulic, A.; Migden, M.R.; Oro, A.E.; Dirix, L.; Lewis, K.D.; Hainsworth, J.D.; Solomon, J.A.; Yoo, S.; Arron, S.T.; Friedlander, P.A.; et al. Efficacy and safety of vismodegib in advanced basal-cell carcinoma. N. Engl. J. Med. 2012, 366, 2171–2179. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ping, X.L.; Lee, P.K.; Wu, X.L.; Yao, Y.J.; Zhang, M.J.; Silvers, D.N.; Ratner, D.; Malhotra, R.; Peacocke, M.; et al. Role of P.PTCH and p53 genes in early-onset basal cell carcinoma. Am. J. Pathol. 2001, 158, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Ouhtit, A.; Nakazawa, H.; Armstrong, B.K.; Kricker, A.; Tan, E.; Yamasaki, H.; English, D.R. UV-radiation-specific p53 mutation frequency in normal skin as a predictor of risk of basal cell carcinoma. J. Natl. Cancer Inst. 1998, 90, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agar, N.S.; Halliday, G.M.; Barnetson, R.S.; Ananthaswamy, H.N.; Wheeler, M.; Jones, A.M. The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: A role for UVA in human skin carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4954–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallingford, S.C.; Hughes, M.C.; Green, A.C.; van der Pols, J.C. Plasma omega-3 and omega-6 concentrations and risk of cutaneous basal and squamous cell carcinomas in Australian adults. Cancer Epidemiol. Prev. Biomark. 2013, 22, 1900–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seviiri, M.; Law, M.H.; Ong, J.S.; Gharahkhani, P.; Nyholt, D.R.; Olsen, C.M.; Whiteman, D.C.; MacGregor, S. Polyunsaturated Fatty Acid Levels and the Risk of Keratinocyte Cancer: A Mendelian Randomization Analysis. Cancer Epidemiol. Prev. Biomark. 2021, 30, 1591–1598. [Google Scholar] [CrossRef]
- Willemze, R.; Jaffe, E.S.; Burg, G.; Cerroni, L.; Berti, E.; Swerdlow, S.H.; Ralfkiaer, E.; Chimenti, S.; Diaz-Perez, J.L.; Duncan, L.M.; et al. WHO-EORTC classification for cutaneous lymphomas. Blood 2005, 105, 3768–3785. [Google Scholar] [CrossRef] [Green Version]
- Cook, L.B.; Fuji, S.; Hermine, O.; Bazarbachi, A.; Ramos, J.C.; Ratner, L.; Horwitz, S.; Fields, P.; Tanase, A.; Bumbea, H.; et al. Revised Adult T-Cell Leukemia-Lymphoma International Consensus Meeting Report. J. Clin. Oncol. 2019, 37, 677–687. [Google Scholar] [CrossRef]
- Sawada, Y.; Hino, R.; Hama, K.; Ohmori, S.; Fueki, H.; Yamada, S.; Fukamachi, S.; Tajiri, M.; Kubo, R.; Yoshioka, M.; et al. Type of skin eruption is an independent prognostic indicator for adult T-cell leukemia/lymphoma. Blood 2011, 117, 3961–3967. [Google Scholar] [CrossRef]
- Sawada, Y.; Shimauchi, T.; Yamaguchi, T.; Okura, R.; Hama-Yamamoto, K.; Fueki-Yoshioka, H.; Ohmori, S.; Yamada, S.; Yoshizawa, M.; Hiromasa, K.; et al. Combination of skin-directed therapy and oral etoposide for smoldering adult T-cell leukemia/lymphoma with skin involvement. Leuk. Lymphoma 2013, 54, 520–527. [Google Scholar] [CrossRef]
- Tokura, Y.; Sawada, Y.; Shimauchi, T. Skin manifestations of adult T-cell leukemia/lymphoma: Clinical, cytological and immunological features. J. Dermatol. 2014, 41, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Tomonaga, M.; Fukuda, H.; Hanada, S.; Utsunomiya, A.; Tara, M.; Sano, M.; Ikeda, S.; Takatsuki, K.; Kozuru, M.; et al. A new G-CSF-supported combination chemotherapy, LSG15, for adult T-cell leukaemia-lymphoma: Japan Clinical Oncology Group Study 9303. Br. J. Haematol. 2001, 113, 375–382. [Google Scholar] [CrossRef]
- Tsukasaki, K.; Utsunomiya, A.; Fukuda, H.; Shibata, T.; Fukushima, T.; Takatsuka, Y.; Ikeda, S.; Masuda, M.; Nagoshi, H.; Ueda, R.; et al. VCAP-AMP-VECP compared with biweekly CHOP for adult T-cell leukemia-lymphoma: Japan Clinical Oncology Group Study JCOG9801. J. Clin. Oncol. 2007, 25, 5458–5464. [Google Scholar] [CrossRef] [PubMed]
- Stanchina, M.; Soong, D.; Zheng-Lin, B.; Watts, J.M.; Taylor, J. Advances in Acute Myeloid Leukemia: Recently Approved Therapies and Drugs in Development. Cancers 2020, 12, 3225. [Google Scholar] [CrossRef] [PubMed]
- Bladé, J.; Dimopoulos, M.; Rosiñol, L.; Rajkumar, S.V.; Kyle, R.A. Smoldering (asymptomatic) multiple myeloma: Current diagnostic criteria, new predictors of outcome, and follow-up recommendations. J. Clin. Oncol. 2010, 28, 690–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.; Bellon, M.; Nicot, C. Emodin and DHA potently increase arsenic trioxide interferon-alpha-induced cell death of HTLV-I-transformed cells by generation of reactive oxygen species and inhibition of Akt and AP-1. Blood 2007, 109, 1653–1659. [Google Scholar] [CrossRef]
- Crombie, J.L.; Armand, P. Diffuse Large B-Cell Lymphoma’s New Genomics: The Bridge and the Chasm. J. Clin. Oncol. 2020, 38, 3565–3574. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, B.; O’Connor, H.M.; Wang, A.H.; Liebow, M.; Thompson, C.A.; Fredericksen, Z.S.; Macon, W.R.; Slager, S.L.; Call, T.G.; Habermann, T.M.; et al. Trans fatty acid intake is associated with increased risk and n3 fatty acid intake with reduced risk of non-hodgkin lymphoma. J. Nutr. 2013, 143, 672–681. [Google Scholar] [CrossRef] [Green Version]
- Thanarajasingam, G.; Maurer, M.J.; Habermann, T.M.; Nowakowski, G.S.; Bennani, N.N.; Thompson, C.A.; Cerhan, J.R.; Witzig, T.E. Low Plasma Omega-3 Fatty Acid Levels May Predict Inferior Prognosis in Untreated Diffuse Large B-Cell Lymphoma: A New Modifiable Dietary Biomarker? Nutr. Cancer 2018, 70, 1088–1090. [Google Scholar] [CrossRef]
- Lomas, A.; Leonardi-Bee, J.; Bath-Hextall, F. A systematic review of worldwide incidence of nonmelanoma skin cancer. Br. J. Dermatol. 2012, 166, 1069–1080. [Google Scholar] [CrossRef]
- Corchado-Cobos, R.; García-Sancha, N.; González-Sarmiento, R.; Pérez-Losada, J.; Cañueto, J. Cutaneous Squamous Cell Carcinoma: From Biology to Therapy. Int. J. Mol. Sci. 2020, 21, 2956. [Google Scholar] [CrossRef] [PubMed]
- Burton, K.A.; Ashack, K.A.; Khachemoune, A. Cutaneous Squamous Cell Carcinoma: A Review of High-Risk and Metastatic Disease. Am. J. Clin. Dermatol. 2016, 17, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Claveau, J.; Archambault, J.; Ernst, D.S.; Giacomantonio, C.; Limacher, J.J.; Murray, C.; Parent, F.; Zloty, D. Multidisciplinary management of locally advanced and metastatic cutaneous squamous cell carcinoma. Curr. Oncol. 2020, 27, e399–e407. [Google Scholar] [CrossRef] [PubMed]
- Elattar, T.M.; Lin, H.S. Comparison of the inhibitory effect of polyunsaturated fatty acids on prostaglandin synthesis I oral squamous carcinoma cells. Prostaglandins Leukot. Essent. Fat. Acids 1989, 38, 119–125. [Google Scholar] [CrossRef]
- Nikolakopoulou, Z.; Nteliopoulos, G.; Michael-Titus, A.T.; Parkinson, E.K. Omega-3 polyunsaturated fatty acids selectively inhibit growth in neoplastic oral keratinocytes by differentially activating ERK1/2. Carcinogenesis 2013, 34, 2716–2725. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Scheff, N.N.; Bernabé, D.; Salvo, E.; Ono, K.; Liu, C.; Veeramachaneni, R.; Viet, C.T.; Viet, D.T.; Dolan, J.C.; et al. Anti-cancer and analgesic effects of resolvin D2 in oral squamous cell carcinoma. Neuropharmacology 2018, 139, 182–193. [Google Scholar] [CrossRef]
- Palakurthi, S.S.; Flückiger, R.; Aktas, H.; Changolkar, A.K.; Shahsafaei, A.; Harneit, S.; Kilic, E.; Halperin, J.A. Inhibition of translation initiation mediates the anticancer effect of the n-3 polyunsaturated fatty acid eicosapentaenoic acid. Cancer Res. 2000, 60, 2919–2925. [Google Scholar]
- Wen, B.; Deutsch, E.; Opolon, P.; Auperin, A.; Frascogna, V.; Connault, E.; Bourhis, J. n-3 polyunsaturated fatty acids decrease mucosal/epidermal reactions and enhance antitumour effect of ionising radiation with inhibition of tumour angiogenesis. Br. J. Cancer 2003, 89, 1102–1107. [Google Scholar] [CrossRef] [Green Version]
- Weed, H.G.; Ferguson, M.L.; Gaff, R.L.; Hustead, D.S.; Nelson, J.L.; Voss, A.C. Lean body mass gain in patients with head and neck squamous cell cancer treated perioperatively with a protein- and energy-dense nutritional supplement containing eicosapentaenoic acid. Head Neck 2011, 33, 1027–1033. [Google Scholar] [CrossRef]
- Solís-Martínez, O.; Plasa-Carvalho, V.; Phillips-Sixtos, G.; Trujillo-Cabrera, Y.; Hernández-Cuellar, A.; Queipo-García, G.E.; Meaney-Mendiolea, E.; Ceballos-Reyes, G.M.; Fuchs-Tarlovsky, V. Effect of Eicosapentaenoic Acid on Body Composition and Inflammation Markers in Patients with Head and Neck Squamous Cell Cancer from a Public Hospital in Mexico. Nutr. Cancer 2018, 70, 663–670. [Google Scholar] [CrossRef]
- Kubota, H.; Hirai, T.; Yamauchi, A.; Ogo, A.; Matsumoto, H.; Ueno, T. Inhibitory Effect of Eicosapentaenoic Acid on the Migration of the Esophageal Squamous Cell Carcinoma Cell Line TE-1. Anticancer Res. 2020, 40, 5043–5048. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Gallo, R.L. Role of Epigenetics in the Regulation of Immune Functions of the Skin. J. Investig. Dermatol. 2021, 141, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Nakatsuji, T.; Dokoshi, T.; Kulkarni, N.N.; Liggins, M.C.; Sen, G.; Gallo, R.L. Cutaneous innate immune tolerance is mediated by epigenetic control of MAP2K3 by HDAC8/9. Sci. Immunol. 2021, 6, eabe1935. [Google Scholar] [CrossRef]
- Sugino, H.; Sawada, Y.; Nakamura, M. IgA Vasculitis: Etiology, Treatment, Biomarkers and Epigenetic Changes. Int. J. Mol. Sci. 2021, 22, 7538. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Mo, M.; Zhong, Y.; Yang, Q.; Zhang, J.; Ye, X.; Zhang, L.; Cai, C. The Anticancer Role of Omega-3 Polyunsaturated Fatty Acids was Closely Associated with the Increase in Genomic DNA Hydroxymethylation. Anticancer Agents Med. Chem. 2019, 19, 330–336. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, M.; Lee, Y.B.; Yu, D.S. Investigation of Genetic Mutations in High-risk and Low-risk Basal Cell Carcinoma in a Non-Caucasian Population by Whole Exome Sequencing. Acta Derm. Venereol. 2021, 101, adv00458. [Google Scholar] [CrossRef]
- Li, L.; Li, F.; Xia, Y.; Yang, X.; Lv, Q.; Fang, F.; Wang, Q.; Bu, W.; Wang, Y.; Zhang, K.; et al. UVB induces cutaneous squamous cell carcinoma progression by de novo ID4 methylation via methylation regulating enzymes. EBioMedicine 2020, 57, 102835. [Google Scholar] [CrossRef]
- Collignon, E.; Canale, A.; Al Wardi, C.; Bizet, M.; Calonne, E.; Dedeurwaerder, S.; Garaud, S.; Naveaux, C.; Barham, W.; Wilson, A.; et al. Immunity drives TET1 regulation in cancer through NF-κB. Sci. Adv. 2018, 4, eaap7309. [Google Scholar] [CrossRef] [Green Version]
- Woollard, W.J.; Pullabhatla, V.; Lorenc, A.; Patel, V.M.; Butler, R.M.; Bayega, A.; Begum, N.; Bakr, F.; Dedhia, K.; Fisher, J.; et al. Candidate driver genes involved in genome maintenance and DNA repair in Sézary syndrome. Blood 2016, 127, 3387–3397. [Google Scholar] [CrossRef] [Green Version]
- Tsuzuki, T.; Kambe, T.; Shibata, A.; Kawakami, Y.; Nakagawa, K.; Miyazawa, T. Conjugated EPA activates mutant p53 via lipid peroxidation and induces p53-dependent apoptosis in DLD-1 colorectal adenocarcinoma human cells. Biochim. Biophys. Acta 2007, 1771, 20–30. [Google Scholar] [CrossRef]
- Kato, T.; Kolenic, N.; Pardini, R.S. Docosahexaenoic acid (DHA), a primary tumor suppressive omega-3 fatty acid, inhibits growth of colorectal cancer independent of p53 mutational status. Nutr. Cancer 2007, 58, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Nishimura, K.; Aires, V.; Yamashita, T.; Oaxaca-Castillo, D.; Kashiwagi, K.; Igarashi, K. Docosahexaenoic acid inhibits cancer cell growth via p27Kip1, CDK2, ERK1/ERK2, and retinoblastoma phosphorylation. J. Lipid Res. 2006, 47, 2306–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, N.; Budhraja, A.; Cheng, S.; Liu, E.H.; Huang, C.; Chen, J.; Yang, Z.; Chen, D.; Zhang, Z.; Shi, X. Interruption of the MEK/ERK signaling cascade promotes dihydroartemisinin-induced apoptosis in vitro and in vivo. Apoptosis 2011, 16, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, Y.; Gu, Z.; Owens, R.T.; Chen, Y.Q.; Edwards, I.J. Omega-3 fatty acids induce apoptosis in human breast cancer cells and mouse mammary tissue through syndecan-1 inhibition of the MEK-Erk pathway. Carcinogenesis 2011, 32, 1518–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, J.I.; McCaskey, S.J. Curcumin and docosahexaenoic acid block insulin-induced colon carcinoma cell proliferation. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 219–226. [Google Scholar] [CrossRef]
- Mashima, E.; Sawada, Y.; Yamaguchi, T.; Yoshioka, H.; Ohmori, S.; Haruyama, S.; Yoshioka, M.; Okada, E.; Nakamura, M. A high expression of cell adhesion molecule 1 (CADM1) is an unfavorable prognostic factor in mycosis fungoides. Clin. Immunol. 2018, 193, 121–122. [Google Scholar] [CrossRef]
- Sawada, Y.; Mashima, E.; Saito-Sasaki, N.; Nakamura, M. The Role of Cell Adhesion Molecule 1 (CADM1) in Cutaneous Malignancies. Int. J. Mol. Sci. 2020, 21, 9732. [Google Scholar] [CrossRef]
- Saito-Sasaki, N.; Sawada, Y.; Okada, E.; Nakamura, M. Cell Adhesion Molecule 1 (CADM1) Is an Independent Prognostic Factor in Patients with Cutaneous Squamous Cell Carcinoma. Diagnostics 2021, 11, 830. [Google Scholar] [CrossRef]
- Pasqualini, M.E.; Heyd, V.L.; Manzo, P.; Eynard, A.R. Association between E-cadherin expression by human colon, bladder and breast cancer cells and the 13-HODE:15-HETE ratio. A possible role of their metastatic potential. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 9–16. [Google Scholar] [CrossRef]
- Iwamoto, S.; Senzaki, H.; Kiyozuka, Y.; Ogura, E.; Takada, H.; Hioki, K.; Tsubura, A. Effects of fatty acids on liver metastasis of ACL-15 rat colon cancer cells. Nutr. Cancer 1998, 31, 143–150. [Google Scholar] [CrossRef]
- Mashima, E.; Inoue, A.; Sakuragi, Y.; Yamaguchi, T.; Sasaki, N.; Hara, Y.; Omoto, D.; Ohmori, S.; Haruyama, S.; Sawada, Y.; et al. Nivolumab in the treatment of malignant melanoma: Review of the literature. Onco Targets Ther. 2015, 8, 2045–2051. [Google Scholar] [CrossRef] [Green Version]
- Saito, R.; Sawada, Y.; Saito-Sasaki, N.; Yamamoto, K.; Yoshioka, H.; Ohmori, S.; Yoshioka, M.; Okada, E.; Nakamura, M. Profile fluctuation of peripheral blood in advanced melanoma patients treated with nivolumab. J. Dermatol. 2018, 45, 1452–1455. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Sawada, Y.; Nakamura, M. Immune Profile Analysis in Peripheral Blood and Tumor in Patients with Malignant Melanoma. Int. J. Mol. Sci. 2021, 22, 1957. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Sawada, Y.; Okada, E.; Yamaguchi, T.; Ohmori, S.; Haruyama, S.; Yoshioka, M.; Nakamura, M. Hypopituitarism and hypothyroidism following atrioventricular block during nivolumab treatment. J. Dermatol. 2017, 44, e144–e145. [Google Scholar] [CrossRef]
- Sato, S.; Sawada, Y.; Nakamura, M. STING Signaling and Skin Cancers. Cancers 2021, 13, 5603. [Google Scholar] [CrossRef]
- Carlsson, J.A.; Wold, A.E.; Sandberg, A.S.; Östman, S.M. The Polyunsaturated Fatty Acids Arachidonic Acid and Docosahexaenoic Acid Induce Mouse Dendritic Cells Maturation but Reduce T-Cell Responses In Vitro. PLoS ONE 2015, 10, e0143741. [Google Scholar] [CrossRef]
Figure 1.
Oncogenesis signaling of malignant melanoma and possible roles of omega-3 PUFAs. BRAF plays a role in MAPK pathway activation, which regulates the development of tumor cells, such as apoptosis, cell growth, and proliferation. NRAS mutations are related to the regulation of PI3K. Ultraviolet light exposure dysregulates p53 expression, leading to the suppression of tumor cell apoptosis.
Figure 1.
Oncogenesis signaling of malignant melanoma and possible roles of omega-3 PUFAs. BRAF plays a role in MAPK pathway activation, which regulates the development of tumor cells, such as apoptosis, cell growth, and proliferation. NRAS mutations are related to the regulation of PI3K. Ultraviolet light exposure dysregulates p53 expression, leading to the suppression of tumor cell apoptosis.

Figure 2.
Oncogenesis signaling of basal cell carcinoma and possible roles of omega-3 PUFAs. As the detailed oncogenesis of basal cell carcinoma, hedgehog pathway gene mutation activates patched homolog and smoothened homolog, which are responsible for the development of basal cell carcinoma. P53 is suppressed by ultraviolet light exposure, leading to the suppression of apoptosis of basal cell carcinoma.
Figure 2.
Oncogenesis signaling of basal cell carcinoma and possible roles of omega-3 PUFAs. As the detailed oncogenesis of basal cell carcinoma, hedgehog pathway gene mutation activates patched homolog and smoothened homolog, which are responsible for the development of basal cell carcinoma. P53 is suppressed by ultraviolet light exposure, leading to the suppression of apoptosis of basal cell carcinoma.

Figure 3.
Oncogenesis signaling of squamous cell carcinoma and possible roles of omega-3 PUFAs. P53 gene mutation is a crucial step in the development of cutaneous squamous cell carcinoma. P53 is responsible for cell apoptosis, DNA repair, and cell cycle arrest in squamous cell carcinoma. In addition, EGFR-mediated PI3K/AKT signaling and NRAS/RAF/MEK/ERK signaling also contribute to the development of squamous cell carcinoma.
Figure 3.
Oncogenesis signaling of squamous cell carcinoma and possible roles of omega-3 PUFAs. P53 gene mutation is a crucial step in the development of cutaneous squamous cell carcinoma. P53 is responsible for cell apoptosis, DNA repair, and cell cycle arrest in squamous cell carcinoma. In addition, EGFR-mediated PI3K/AKT signaling and NRAS/RAF/MEK/ERK signaling also contribute to the development of squamous cell carcinoma.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of detailed action of omega-3 PUFAs against skin cancers.
| Skin Cancers | Detailed Action |
|---|---|
| Melanoma | Reduce the risk of melanoma by DHA/EPA intake [27] Enhance the sensitivity to chemotherapy by EPA and DHA [28,29,30,31,46,47,48]. Suppress tumor growth by linoleic acid, EPA, DHA [32,37,41,42,44,45]. Suppress tumor metastasis by EPA [33,37] Suppress tumor invasion by EPA and DHA [34,37] Enhance ROS production and P53 induction by DHA and EPA [40]. Suppress debris-stimulated cancer progression by RvD1, RvD2, or RvE1 [50] |
| BCC | Reduce the risk of BCC by α-linolenic acid and linoleic [56]. |
| ATLL | Synergistic effect on cell cycle arrest and cell death by the combination of DHA with chemotherapy [66]. |
| DLBCL | Low intake of omega-3 fatty acids is associated with unfavorable survival [69]. |
| SCC | Enhance the toxicity by DHA [75]. Reduce tumor size and cancer-derived cytokines/chemokines by RvD2 [76]. Suppress cell proliferation by EPA [77] Suppress tumor growth by the combination of Omega-3 PUFAs with ionizing radiation [78]. Suppress migration by EPA [81]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Minokawa, Y.; Sawada, Y.; Nakamura, M. The Influences of Omega-3 Polyunsaturated Fatty Acids on the Development of Skin Cancers. Diagnostics 2021, 11, 2149. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11112149
AMA Style
Minokawa Y, Sawada Y, Nakamura M. The Influences of Omega-3 Polyunsaturated Fatty Acids on the Development of Skin Cancers. Diagnostics. 2021; 11(11):2149. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11112149
Chicago/Turabian StyleMinokawa, Yoko, Yu Sawada, and Motonobu Nakamura. 2021. "The Influences of Omega-3 Polyunsaturated Fatty Acids on the Development of Skin Cancers" Diagnostics 11, no. 11: 2149. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11112149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.