
Genetic Association of the Functional WDR4 Gene in Male Fertility

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Recruitment and Sample Collection
2.2. Semen Analysis
2.3. Genomic DNA Extraction
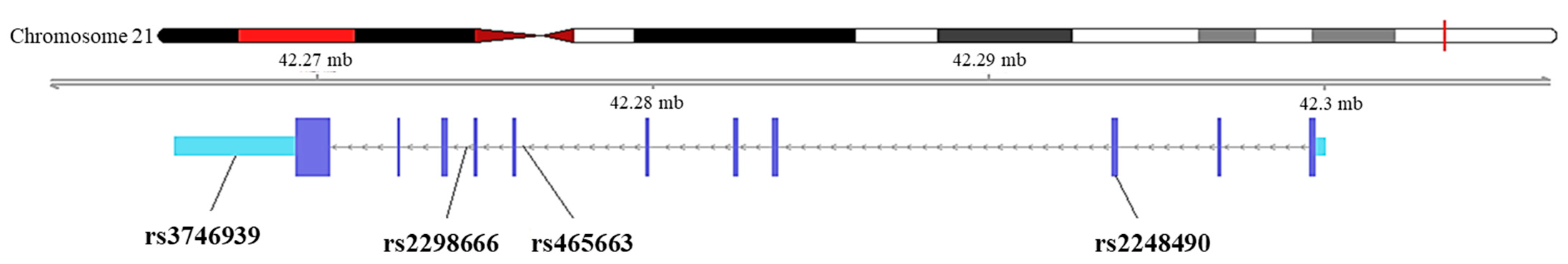
2.4. Genotyping of SNPs in the WDR4 Gene
2.5. Functional Annotation Data Query
2.6. Phenome-Wide Association Study (PheWAS)
2.7. Cell Culture
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. PheWAS for WDR4 Variants in A Taiwanese Population
3.2. Participant Characteristics
3.3. Associations between WDR4 Variants and Oligozoospermia
3.4. Associations between WDR4 Variants and Asthenozoospermia
3.5. Associations between WDR4 Variants and Teratozoospermia
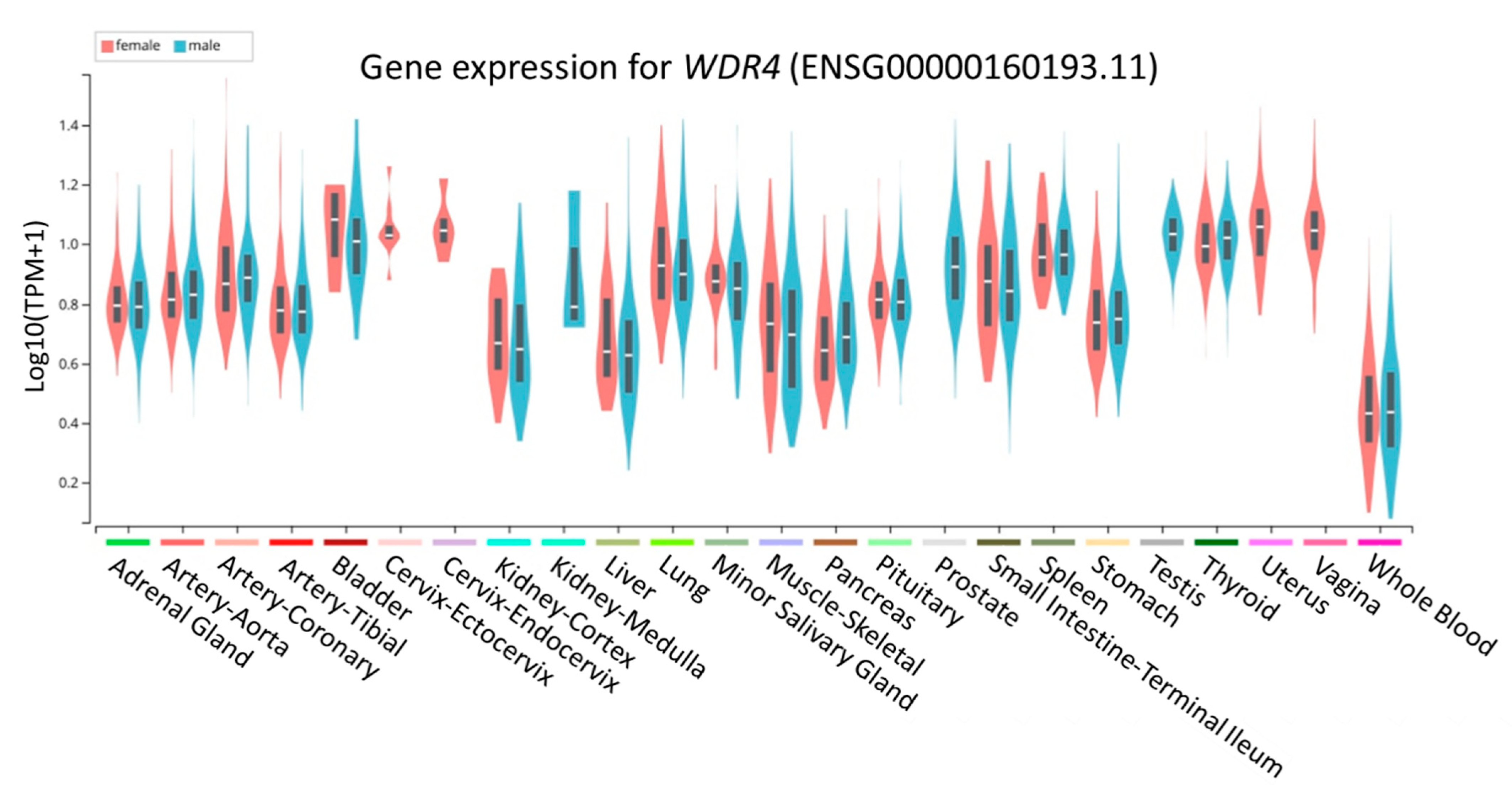
3.6. Functional Annotation from GTEx Portal for rs465663
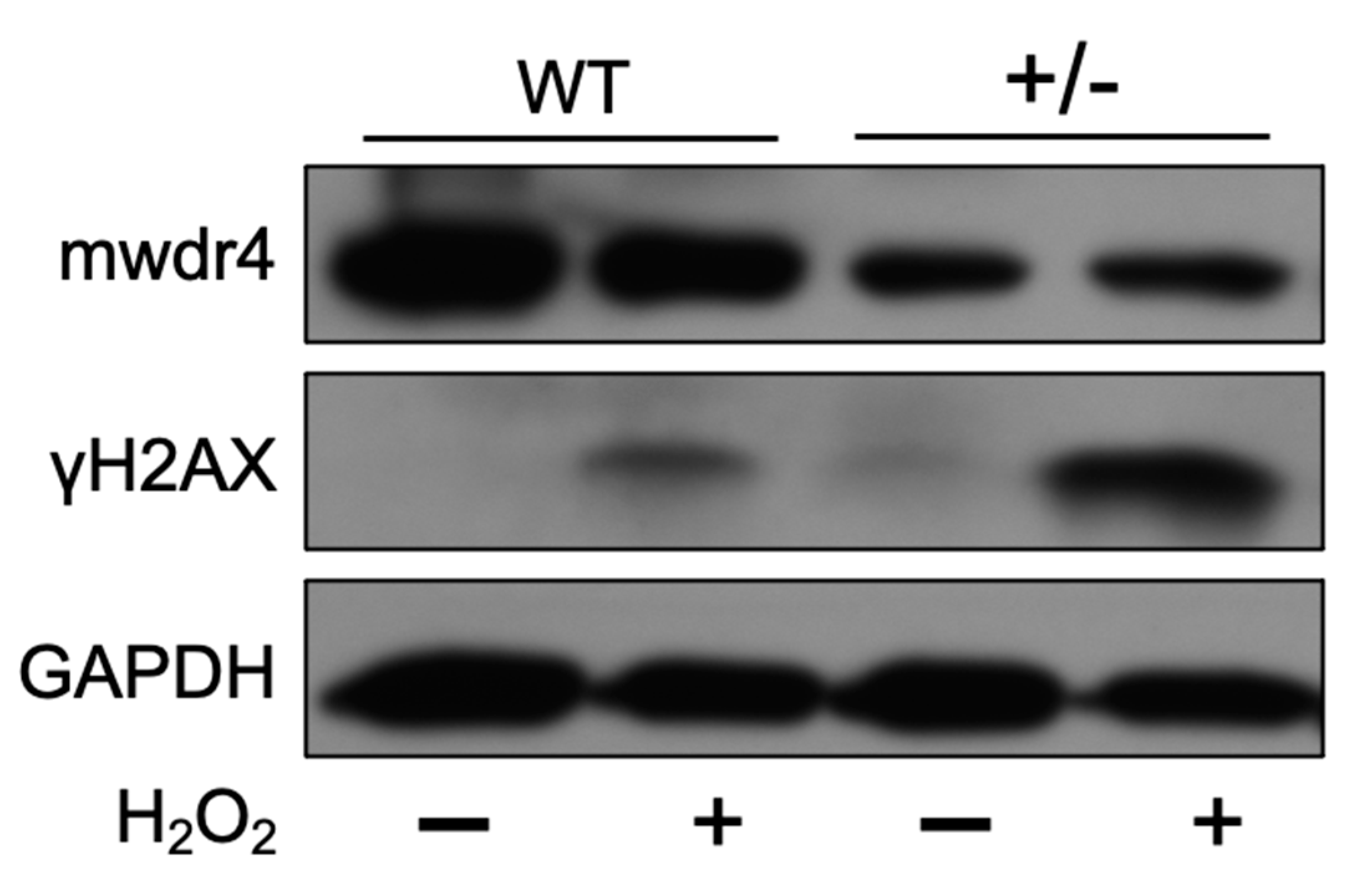
3.7. Effects of WDR4 in DNA Fragmentation through γH2AX Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zegers-Hochschild, F.; Adamson, G.; de Mouzon, J.; Ishihara, O.; Mansour, R.; Nygren, K.; Sullivan, E.; Vanderpoel, S. International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology, 2009. Fertil. Steril. 2009, 92, 1520–1524. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.C. Clinical relevance of routine semen analysis and controversies surrounding the 2010 World Health Organization criteria for semen examination. Int. Braz. J. Urol. 2014, 40, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Giwercman, A.; Tournaye, H.; Diemer, T.; Kopa, Z.; Dohle, G.; Krausz, C.G. European Association of Urology Guidelines on Male Infertility: The 2012 Update. Eur. Urol. 2012, 62, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.H.; Sharma, R.; Thornton, J.; Mascha, E.; Abdel-Hafez, M.A.; Thomas, A.J.; Agarwal, A. Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Hum. Reprod. 2004, 19, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbashir, S.; Magdi, Y.; Rashed, A.; Ibrahim, M.A.; Edris, Y.; Abdelaziz, A.M. Relationship between sperm progressive motility and DNA integrity in fertile and infertile men. Middle East Fertil. Soc. J. 2018, 23, 195–198. [Google Scholar] [CrossRef]
- Hopps, C.V.; Mielnik, A.; Goldstein, M.; Palermo, G.D.; Rosenwaks, Z.; Schlegel, P.N. Detection of sperm in men with Y chromosome microdeletions of the AZFa, AZFb and AZFc regions. Hum. Reprod. 2003, 18, 1660–1665. [Google Scholar] [CrossRef] [Green Version]
- Ozdemir, O.; Gül, E.; Kilicarslan, H.; Gokce, G.; Beyaztas, F.Y.; Ayan, S.; Sezgin, I. SRY and AZF gene variation in male infertility: A cytogenetic and molecular approach. Int. Urol. Nephrol. 2007, 39, 1183–1189. [Google Scholar] [CrossRef]
- Gentiluomo, M.; Crifasi, L.; Luddi, A.; Locci, D.; Barale, R.; Piomboni, P.; Campa, D. Taste receptor polymorphisms and male infertility. Hum. Reprod. 2017, 32, 2324–2331. [Google Scholar] [CrossRef]
- Aston, K.I.; Carrell, D.T. Genome-wide study of single-nucleotide polymorphisms associated with azoospermia and severe oligozoospermia. J. Androl. 2009, 30, 711–725. [Google Scholar] [CrossRef]
- Hu, Z.; Xia, Y.; Guo, X.; Dai, J.; Li, H.; Hu, H.; Jiang, Y.; Lu, F.; Wu, Y.; Yang, X.; et al. A genome-wide association study in Chinese men identifies three risk loci for non-obstructive azoospermia. Nat. Genet. 2011, 44, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Tajima, A.; Shichiri, K.; Tanaka, A.; Tanaka, K.; Inoue, I. Genome-Wide Expression of Azoospermia Testes Demonstrates a Specific Profile and Implicates ART3 in Genetic Susceptibility. PLoS Genet. 2008, 4, e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Hou, J.H.; Hsieh, T.-S. A new Drosophila gene wh (wuho) with WD40 repeats is essential for spermatogenesis and has maximal expression in hub cells. Dev. Biol. 2006, 296, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Purcell, S.; Chang, C. Plink v1.9. Available online: https://www.cog-genomics.org/plink/ (accessed on 18 May 2021).
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Cheng, I.-C.; Chen, B.C.; Shuai, H.-H.; Chien, F.-C.; Chen, P.; Hsieh, T.-S. Wuho Is a New Member in Maintaining Genome Stability through its Interaction with Flap Endonuclease 1. PLoS Biol. 2016, 14, e1002349. [Google Scholar] [CrossRef]
- Gao, X.; Starmer, J.; Martin, E.R. A multiple testing correction method for genetic association studies using correlated single nucleotide polymorphisms. Genet. Epidemiol. 2008, 32, 361–369. [Google Scholar] [CrossRef]
- Pasqualotto, F.F.; Sobreiro, B.P.; Hallak, J.; Pasqualotto, E.B.; Lucon, A.M. Sperm concentration and normal sperm morphology decrease and follicle-stimulating hormone level increases with age. BJU Int. 2005, 96, 1087–1091. [Google Scholar] [CrossRef]
- Zhong, H.-Z.; Lv, F.-T.; Deng, X.-L.; Hu, Y.; Xie, D.-N.; Lin, B.; Mo, Z.-N.; Lin, F.-Q. Evaluating γH2AX in spermatozoa from male infertility patients. Fertil. Steril. 2015, 104, 574–581. [Google Scholar] [CrossRef]
- Riedl, S.J.; Salvesen, G.S. The apoptosome: Signalling platform of cell death. Nat. Rev. Mol. Cell Biol. 2007, 8, 405–413. [Google Scholar] [CrossRef]
- Tahmasebi, S.; Khoutorsky, A.; Mathews, M.B.; Sonenberg, N. Translation deregulation in human disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 791–807. [Google Scholar] [CrossRef]
- Ikechebelu, J.; Adinma, B.; Orie, E.; Ikegwuonu, S. High prevalence of male infertility in southeastern Nigeria. J. Obstet. Gynaecol. 2003, 23, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Nagy, R.; Boutin, T.S.; Marten, J.; Huffman, J.E.; Kerr, S.M.; Campbell, A.; Evenden, L.; Gibson, J.; Amador, C.; Howard, D.M.; et al. Exploration of haplotype research consortium imputation for genome-wide association studies in 20,032 Generation Scotland participants. Genome Med. 2017, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Rastegari, E.; Kajal, K.; Tan, B.-S.; Huang, F.; Chen, R.-H.; Hsieh, T.-S.; Hsu, H.-J. WD40 protein Wuho controls germline homeostasis via TRIM-NHL tumor suppressor Mei-p26 in Drosophila. Development 2020, 147, dev182063. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Jeon, S.; Choi, M.; Lee, M.-H.; Park, M.; Lee, D.R.; Jun, K.-Y.; Kwon, Y.; Lee, O.-H.; Song, S.-H.; et al. Mutations in SOHLH1 gene associate with nonobstructive Azoospermia. Hum. Mutat. 2010, 31, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Hasuike, S.; Yogev, L.; Maduro, M.R.; Ishikawa, M.; Westphal, H.; Lamb, D.J. Azoospermia in patients heterozygous for a mutation in SYCP3. Lancet 2003, 362, 1714–1719. [Google Scholar] [CrossRef]
- Yatsenko, A.N.; Georgiadis, A.P.; Röpke, A.; Berman, A.J.; Jaffe, T.; Olszewska, M.; Westernströer, B.; Sanfilippo, J.; Kurpisz, M.; Rajkovic, A.; et al. X-Linked TEX11 Mutations, Meiotic Arrest, and Azoospermia in Infertile Men. N. Engl. J. Med. 2015, 372, 2097–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaheen, R.; Abdel-Salam, G.; Guy, M.P.; AlOmar, R.; Abdel-Hamid, M.S.; Afifi, H.H.; Ismail, S.; Emam, B.A.; Phizicky, E.M.; Alkuraya, F.S. Mutation in WDR4 impairs tRNA m7G46 methylation and causes a distinct form of microcephalic primordial dwarfism. Genome Biol. 2015, 16, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadoune, J.-P. Spermatozoal RNAs: What about their functions? Microsc. Res. Tech. 2009, 72, 536–551. [Google Scholar] [CrossRef]
- Oscanoa, J.; Sivapalan, L.; Gadaleta, E.; Ullah, A.Z.D.; Lemoine, N.R.; Chelala, C. SNPnexus: A web server for functional annotation of human genome sequence variation (2020 update). Nucleic Acids Res. 2020, 48, W185–W192. [Google Scholar] [CrossRef]
- Torres, A.G.; Batlle, E.; de Pouplana, L.R. Role of tRNA modifications in human diseases. Trends Mol. Med. 2014, 20, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.; Francisco, S.; Varanda, A.S.; Santos, M.; Santos, M.A.S.; Soares, A.R. Impact of tRNA Modifications and tRNA-Modifying Enzymes on Proteostasis and Human Disease. Int. J. Mol. Sci. 2018, 19, 3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.-J.; Shen, L.; Jang, C.-W.; Falnes, P.Ø.; Zhang, Y. Ikbkap/Elp1 Deficiency Causes Male Infertility by Disrupting Meiotic Progression. PLoS Genet. 2013, 9, e1003516. [Google Scholar] [CrossRef] [PubMed]
- Giacone, F.; Cannarella, R.; Mongioì, L.M.; Alamo, A.; Condorelli, R.A.; Calogero, A.E.; La Vignera, S. Epigenetics of Male Fertility: Effects on Assisted Reproductive Techniques. World J. Men’s Health 2019, 37, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, A.; Zhang, Z.; Wang, P.; Qian, Y.; He, L.; Shi, H.; Xing, Q.; Du, J. DNA methylation levels of imprinted and nonimprinted genes DMRs associated with defective human spermatozoa. Andrologia 2016, 48, 1027–1035. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Participants | Normal Range |
|---|---|---|
| Age (years) a | 36.21 ± 4.58 | |
| Range (years) | 27~51 | |
| Semen analysis, no. (%) | ||
| Oligozoospermia (total sperm number < 39 × 106) | ||
| Oligozoospermia | 12 (15) | |
| Isolated | 9 | |
| Combined | 3 | |
| Non-oligozoospermia | 69 (85) | |
| Asthenozoospermia (PR < 32%) | ||
| Asthenozoospermia | 22 (27) | |
| Isolated | 10 | |
| Combined | 12 | |
| Non-asthenozoospermia | 59 (73) | |
| Teratozoospermia (sperm with normal morphology <4%) | ||
| Teratozoospermia | 6 (7) | |
| Isolated | 0 | |
| Combined | 6 | |
| Non-teratozoospermia | 75 (93) | |
| Serum hormone analysis a | ||
| Total testosterone (ng/mL) | 4.87 ± 1.59 | 2.51~10.60 |
| FSH (mIU/mL) | 5.42 ± 4.24 | 1~14 |
| LH (mIU/mL) | 2.40 ± 1.59 | 1.5~9.2 |
| SHBG (nmol/L) | 28.88 ± 14.82 | 14.5~48.4 |
| Inhibin B (pg/mL) | 252.40 ± 162.44 | — |
| TSH (μIU/mL) | 1.63 ± 0.82 | 0.34~5.60 |
| T3 (ng/dL) | 112.20 ± 18.06 | 87~178 |
| T4 (μg/dL) | 8.15 ± 1.28 | 6.09~12.23 |
| Free T4 (ng/dL) | 0.90 ± 0.10 | 0.61~1.12 |
| AMH (ng/mL) | 12.51 ± 7.84 | — |
| Zinc (μg/L) | 1076.10 ± 217.26 | 800~1200 |
| SNP | Genotype | Cases | Non-Oligozoospermia | Recessive Model p-Value | ||
|---|---|---|---|---|---|---|
| No. | % | No. | % | |||
| rs2298666 | GG | 10 | 83.3 | 48 | 70.6 | 0.665 |
| GA | 2 | 16.7 | 18 | 26.5 | ||
| AA | 0 | 0 | 2 | 2.9 | ||
| rs465663 | TT | 9 | 75.0 | 33 | 49.3 | 0.157 |
| TC | 3 | 25.0 | 24 | 35.8 | ||
| CC | 0 | 0 | 10 | 14.9 | ||
| rs2248490 | CC | 8 | 66.7 | 30 | 43.5 | 0.095 |
| CG | 4 | 33.3 | 29 | 42.0 | ||
| GG | 0 | 0 | 10 | 14.5 | ||
| rs3746939 | AA | 8 | 66.7 | 37 | 53.6 | 0.165 |
| AC | 4 | 33.3 | 27 | 39.1 | ||
| CC | 0 | 0 | 5 | 7.2 | ||
| SNP | Genotype | Cases | Non-Asthenozoospermia | Recessive Model p-Value | ||
|---|---|---|---|---|---|---|
| No. | % | No. | % | |||
| rs2298666 | GG | 17 | 81 | 41 | 69.5 | 0.399 |
| GA | 4 | 19 | 16 | 27.1 | ||
| AA | 0 | 0 | 2 | 3.4 | ||
| rs465663 | TT | 15 | 71.4 | 27 | 46.6 | 0.025 * |
| TC | 6 | 28.6 | 21 | 36.2 | ||
| CC | 0 | 0 | 10 | 17.2 | ||
| rs2248490 | CC | 15 | 68.2 | 23 | 39 | 0.677 |
| CG | 5 | 22.7 | 28 | 47.5 | ||
| GG | 2 | 9.1 | 8 | 13.6 | ||
| rs3746939 | AA | 15 | 68.2 | 30 | 50.8 | 0.513 |
| AC | 6 | 27.3 | 25 | 42.4 | ||
| CC | 1 | 4.5 | 4 | 6.8 | ||
| SNP | Genotype | Cases | Non-Teratozoospermia | Recessive Model p-Value | ||
|---|---|---|---|---|---|---|
| No. | % | No. | % | |||
| rs2298666 | GG | 5 | 83.3 | 53 | 71.6 | 0.759 |
| GA | 1 | 16.7 | 19 | 25.7 | ||
| AA | 0 | 0 | 2 | 2.7 | ||
| rs465663 | TT | 5 | 83.3 | 37 | 50.7 | 0.395 |
| TC | 1 | 16.7 | 26 | 35.6 | ||
| CC | 0 | 0 | 10 | 13.7 | ||
| rs2248490 | CC | 5 | 83.3 | 33 | 44 | 0.154 |
| CG | 1 | 16.7 | 32 | 42.7 | ||
| GG | 0 | 0 | 10 | 13.3 | ||
| rs3746939 | AA | 4 | 66.7 | 41 | 54.7 | 0.186 |
| AC | 2 | 33.3 | 29 | 38.7 | ||
| CC | 0 | 0 | 5 | 6.7 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-J.; Mugiyanto, E.; Peng, Y.-T.; Huang, W.-C.; Chou, W.-H.; Lee, C.-C.; Wang, Y.-S.; Irham, L.M.; Perwitasari, D.A.; Hsu, M.-I.; et al. Genetic Association of the Functional WDR4 Gene in Male Fertility. J. Pers. Med. 2021, 11, 760. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11080760
Wang Y-J, Mugiyanto E, Peng Y-T, Huang W-C, Chou W-H, Lee C-C, Wang Y-S, Irham LM, Perwitasari DA, Hsu M-I, et al. Genetic Association of the Functional WDR4 Gene in Male Fertility. Journal of Personalized Medicine. 2021; 11(8):760. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11080760
Chicago/Turabian StyleWang, Yu-Jia, Eko Mugiyanto, Yun-Ting Peng, Wan-Chen Huang, Wan-Hsuan Chou, Chi-Chiu Lee, Yu-Shiuan Wang, Lalu Muhammad Irham, Dyah Aryani Perwitasari, Ming-I Hsu, and et al. 2021. "Genetic Association of the Functional WDR4 Gene in Male Fertility" Journal of Personalized Medicine 11, no. 8: 760. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11080760