Interspecific Competition between the House Fly, Musca domestica L. (Diptera: Muscidae) and Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae) When Reared on Poultry Manure


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
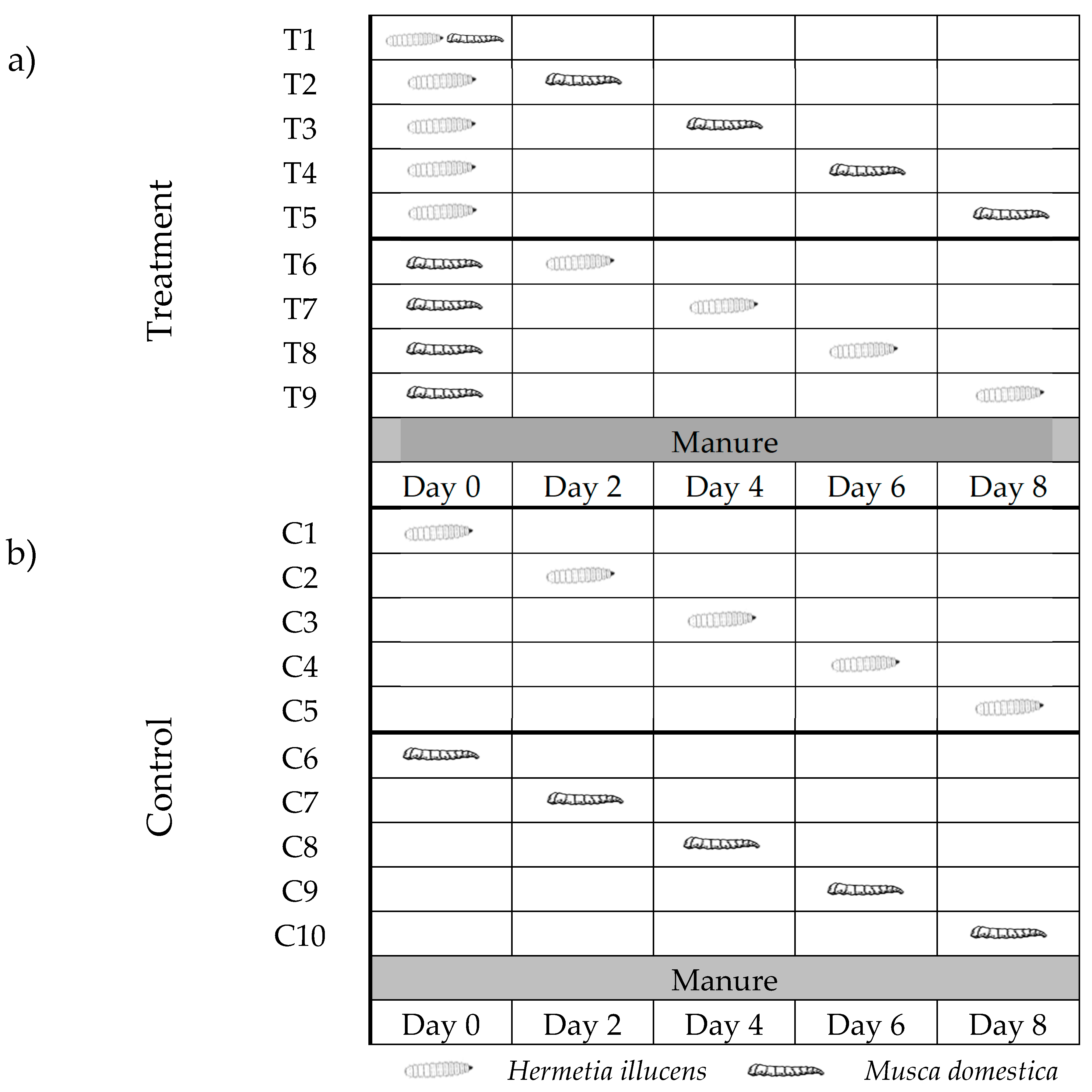
2. Materials and Methods
3. Results
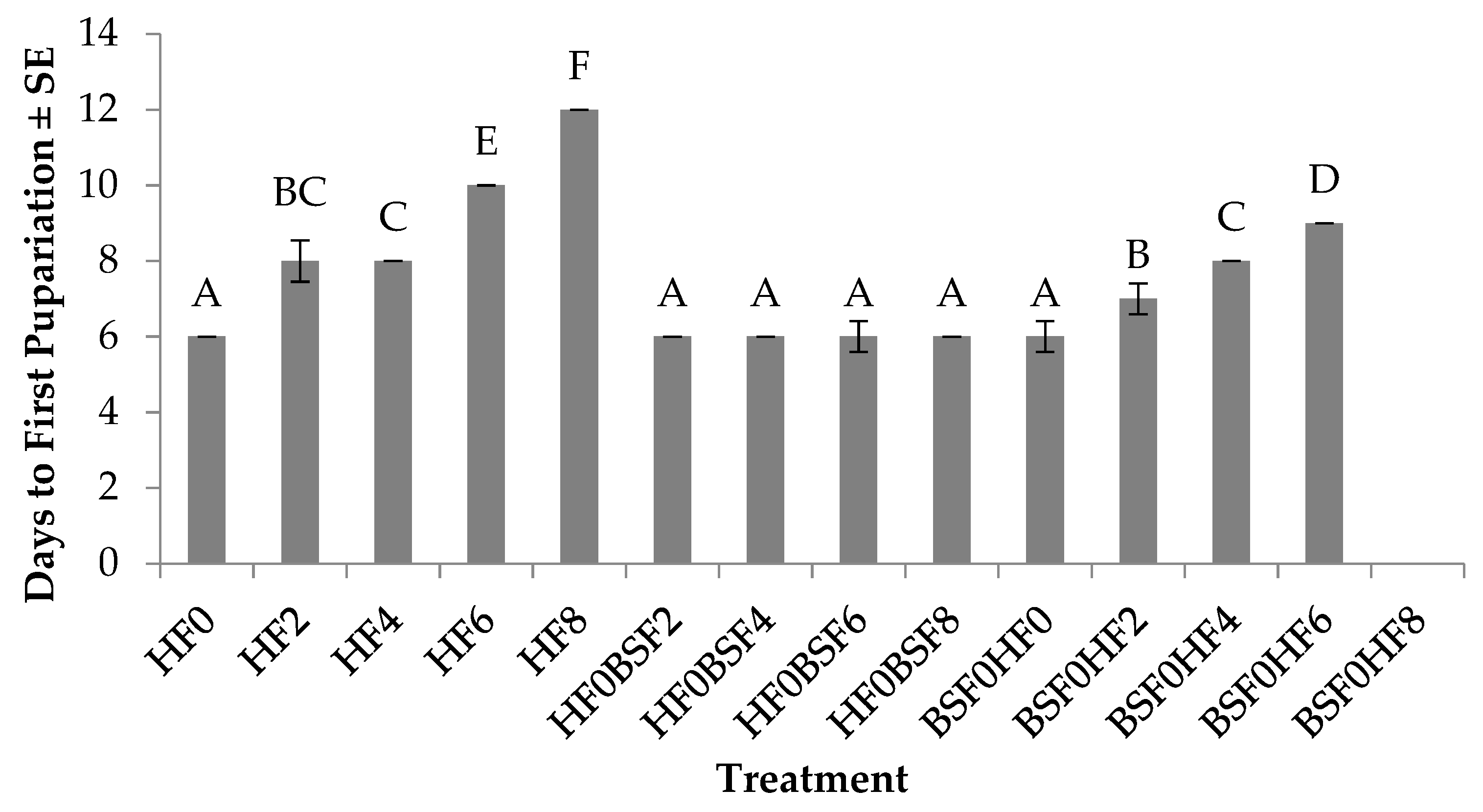
3.1. Time to First Pupariation (HF)
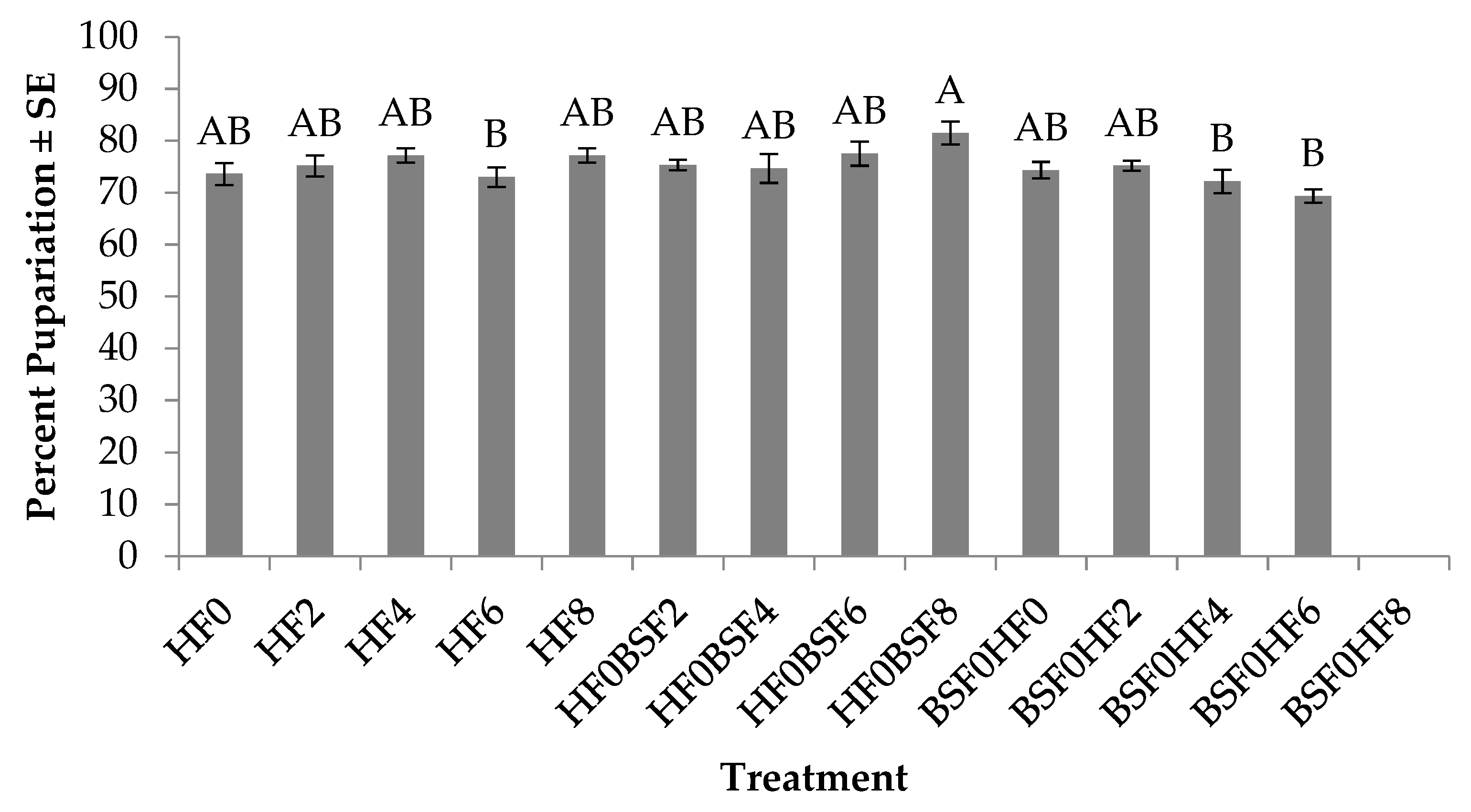
3.2. Percent Pupariation (HF)
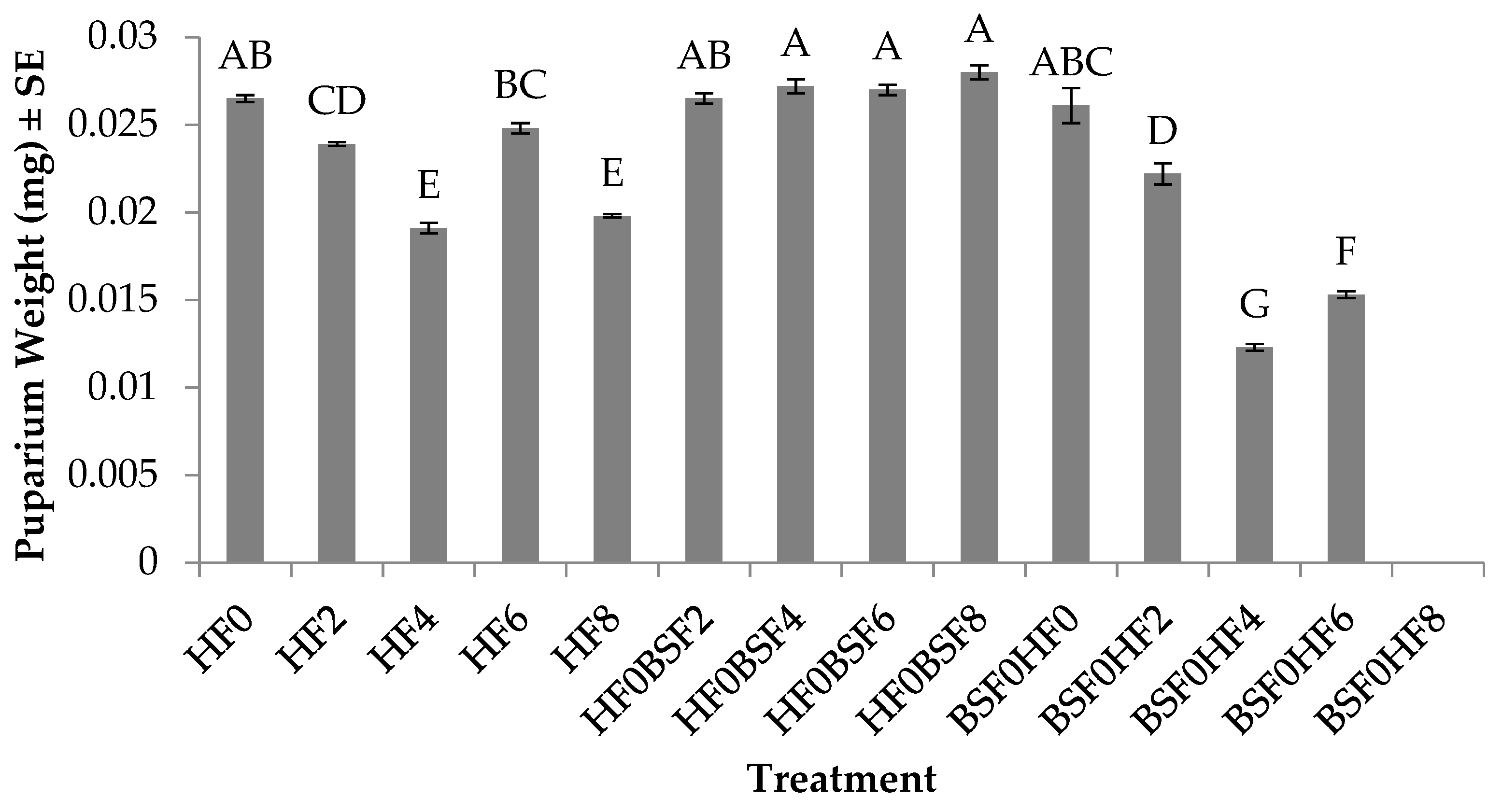
3.3. Puparium Weight (HF)
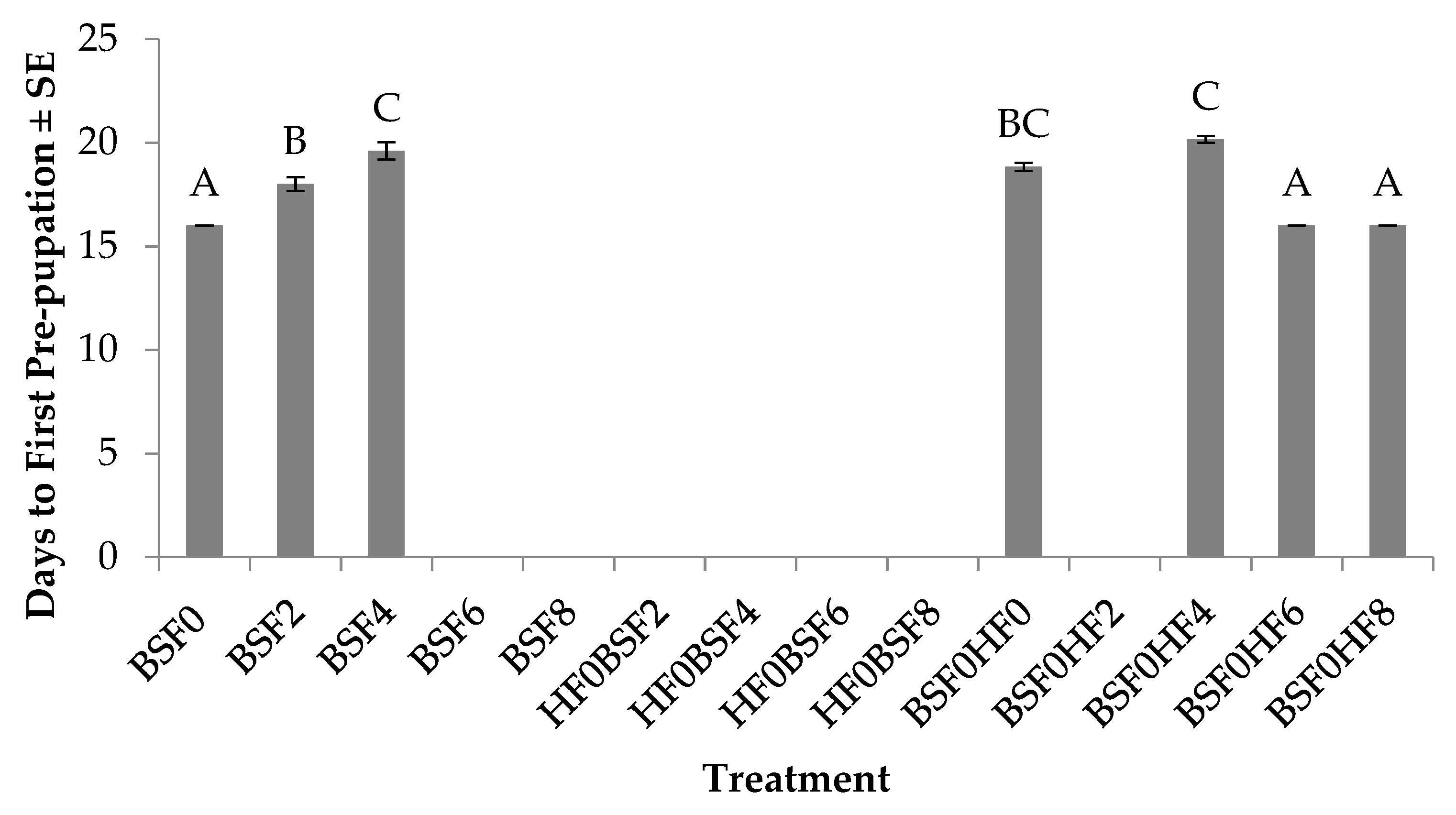
3.4. Time to First Pre-pupation (BSF)
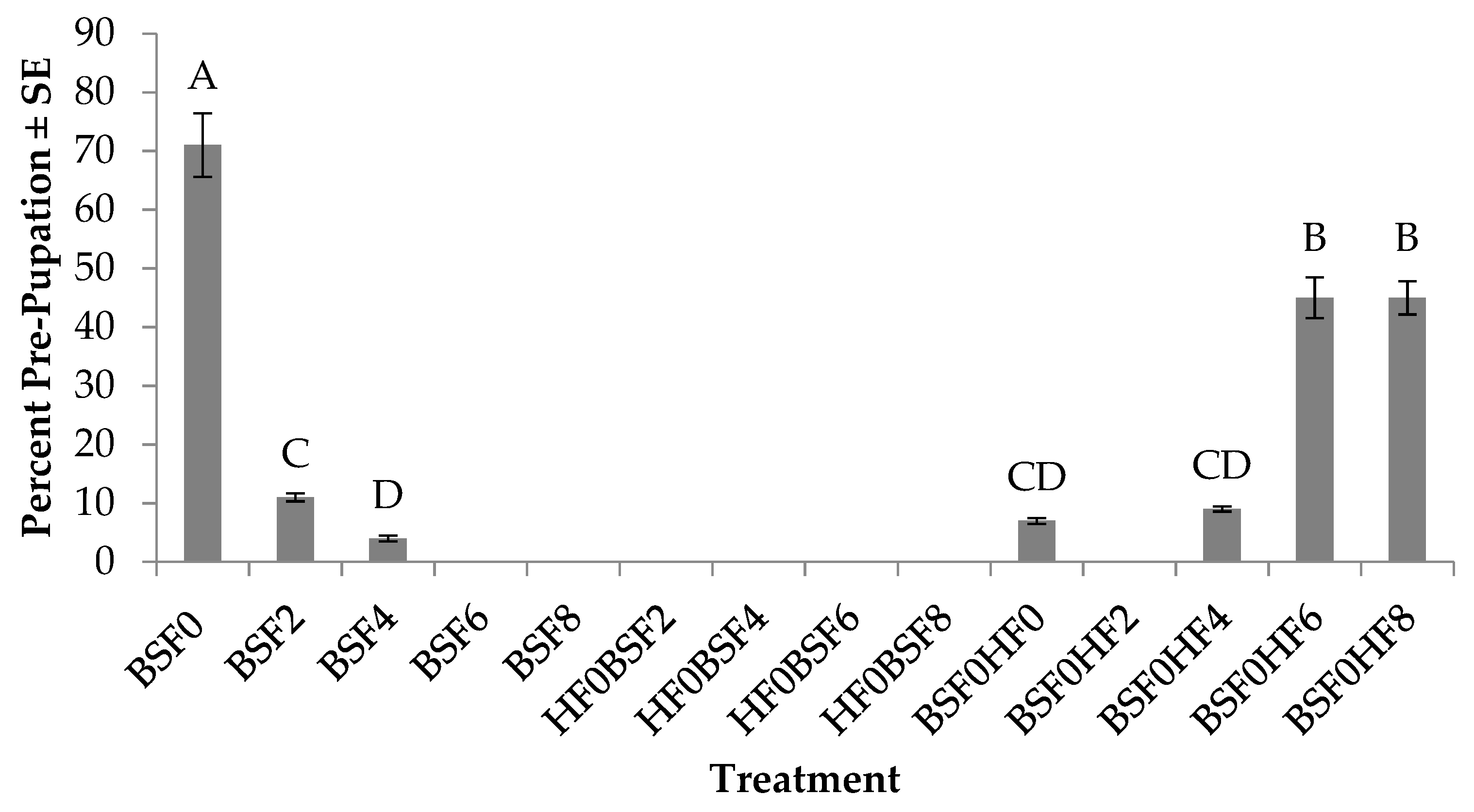
3.5. Percent Pre-pupation (BSF)
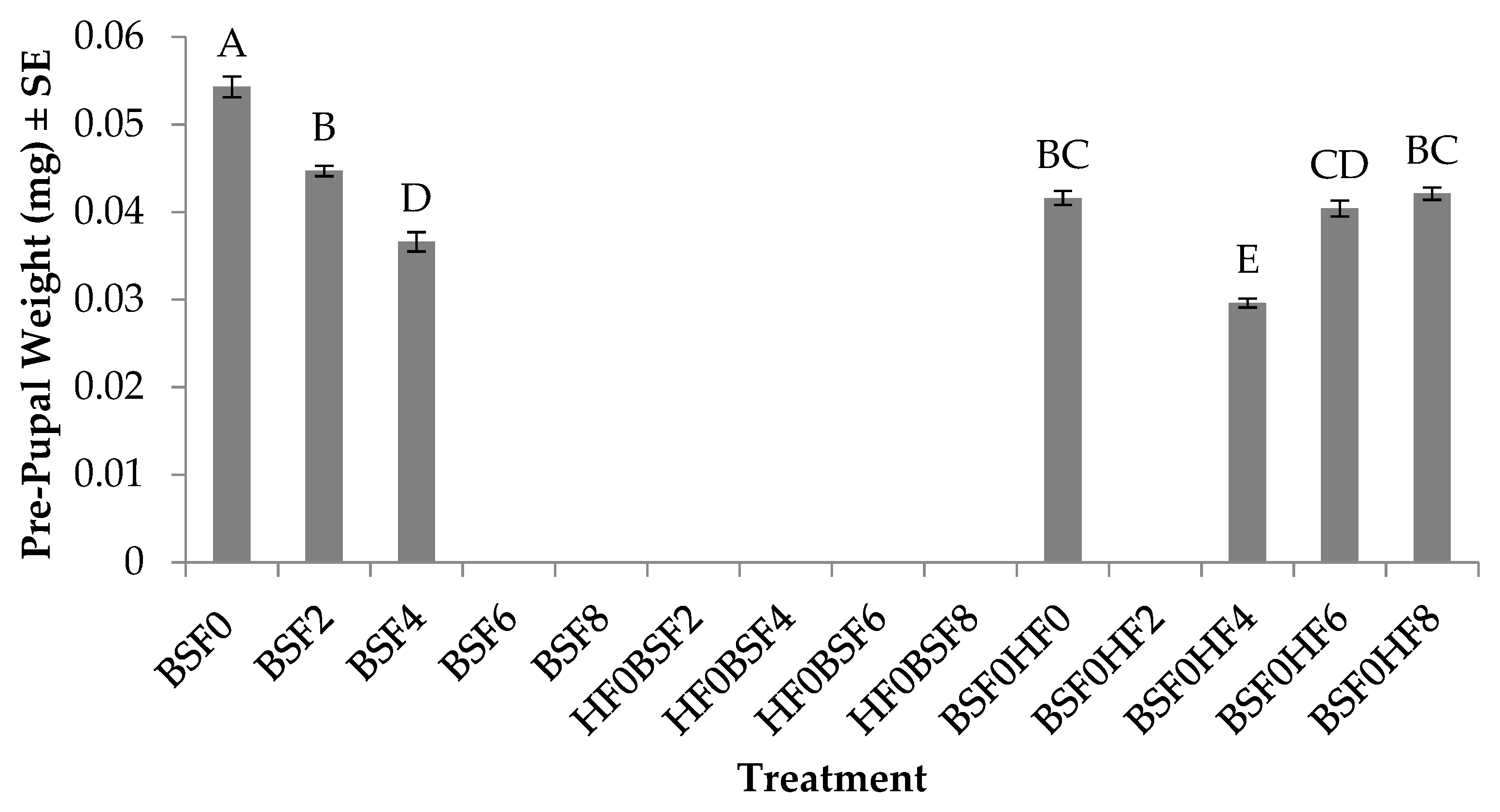
3.6. Pre-pupal Weight (BSF)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Begon, M.; Townsend, C.R.; Harper, J.L. Ecology: From Individuals to Ecosystems; Blackwell Publishing: Hoboken, NJ, USA, 2006. [Google Scholar]
- Taylor, D.B.; Moon, R.D.; Mark, D.R. Economic impact of stable flies (Diptera: Muscidae) on dairy and beef cattle production. J. Med. Entomol. 2012, 49, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Larrain, P.S.; Salas, C.F. House fly (Musca domestica L.) (Diptera: Muscidae) development in different types of manure. Chil. J. Agric. Res. 2008, 68, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-History traits of the house fly, Musca domestica L. (Diptera: Muscidae), reared on three manure types. J. Insects Food Feed 2019, 1–10. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Knight, R.; Gilman, R.H.; Cranfield, M.R. The role of non-biting flies in the epidemiology of human infectious diseases. Microbes Infect. 2001, 3, 231–235. [Google Scholar] [CrossRef]
- Thomas, G.D.; Skoda, S.R. Rural Flies in the Urban Environment; University of Nebraska-Lincoln, Institute of Agriculture and Natural Resources, Agriculture Experiment Station: Lincoln, Nebraska, 1993. [Google Scholar]
- Sawicki, R.; Lord, K. Some properties of a mechanism delaying penetration of insecticides into houseflies. J. Pestic. Sci. 1970, 1, 213–217. [Google Scholar] [CrossRef]
- Afifi, S.E.; Knutson, H. Reproductive potential, longevity, and weight of house flies which survived one insecticidal treatment. J. Econ. Entomol. 1956, 49, 310–313. [Google Scholar] [CrossRef]
- Georghiou, G. Distribution of insecticide-resistant house flies on neighboring farms. J. Econ. Entomol. 1966, 59, 341–346. [Google Scholar] [CrossRef]
- Busvine, J.R. Patterns of insecticide resistance to organo-phosphorus compounds in strains of houseflies from various sources. Entomol. Exp. Appl. 1959, 2, 58–67. [Google Scholar] [CrossRef]
- El Basheir, S. Causes of resistence to DDT in diazinon-selected and DDT-selected strain of house flies. Entomol. Exp. Appl. 1967, 10, 111–126. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Hawley, M.K. Insecticide resistance resulting from sequential selection of houseflies in the field by organophosphorus compounds. Bull. World Health Organ. 1971, 45, 43–51. [Google Scholar]
- Axtell, R.C. Poultry integrated pest management: Status and future. Integr. Pest Manag. Rev. 1999, 4, 53–73. [Google Scholar] [CrossRef]
- Axtell, R.; Edwards, T. Hermetia illucens control in poultry manure by larviciding. J. Econ. Entomol. 1970, 63, 1786–1787. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Influence of resources on Hermetia illucens (Diptera: Stratiomyidae) larval development. J. Med. Entomol. 2013, 50, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D.C.; Newton, G.L.; Thompson, S.A.; Savage, S. A value-added manure management-system using the black soldier fly. Bioresour. Technol. 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Sheppard, C. Housefly and lesser fly control utilizing the black soldier fly in manure management systems for caged laying hens. Environ. Entomol. 1983, 12, 1439–1442. [Google Scholar] [CrossRef]
- Myers, H.M.; Tomberlin, J.K.; Lambert, B.D.; Kattes, D. Development of black soldier fly (Diptera: Stratiomyidae) larvae fed dairy manure. Environ. Entomol. 2008, 37, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Beskin, K.V.; Holcomb, C.D.; Cammack, J.A.; Crippen, T.L.; Knap, A.H.; Sweet, S.T.; Tomberlin, J.K. Larval digestion of different manure types by the black soldier fly (Diptera: Stratiomyidae) impacts associated volatile emissions. Waste Manag. 2018, 74, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.P.; Young, R.D.; Catts, E.P. Hermetia illucens (Linnaeus) as a factor in the natural control of Musca domestica Linnaeus. J. Econ. Entomol. 1959, 52, 917–921. [Google Scholar] [CrossRef]
- Bradley, S.W.; Sheppard, D.C. Housefly oviposition inhibition by larvae of Hermetia illucens, the black soldier fly. J. Chem. Ecol. 1984, 10, 853–859. [Google Scholar] [CrossRef]
- Kilpatrick, J.W.; Schoof, H.F. Interrelationship of water and Hermetia illucens breeding to Musca domestica production in human excrement. Am. J. Trop. Med. Hyg. 1959, 8, 597–602. [Google Scholar] [CrossRef]
- Erickson, M.C.; Islam, M.; Sheppard, C.; Liao, J.; Doyle, M.P. Reduction of Escherichia coli O157:H7 and Salmonella enterica serovar enteritidis in chicken manure by larvae of the black soldier fly. J. Food Prot. 2004, 67, 685–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Tomberlin, J.K.; Brady, J.A.; Sanford, M.R.; Yu, Z. Black soldier fly (Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environ. Entomol. 2008, 37, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Schmidtmann, E.; Martin, P. Relationship between selected bacteria and the growth of immature house flies, Musca domestica, in an axenic test system. J. Med. Entomol. 1992, 29, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Rochon, K. Persistence and Significance of E. coli in House Flies (Musca Domestica) and Stable Flies (Stomoxys Calcitrans); University of Lethbridge, Faculty of Arts and Science: Lethbridge, AB, Canada, 2003. [Google Scholar]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-History traits of the black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), reared on three manure types. Animals 2019, 9, 281. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Eaton, A.; Clesceri, L.; Rice, E.; Greenberg, A.; Franson, M. Total suspended solids dried at 103–105 C. In Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Hogsette, J.A. New diets for production of house flies and stable flies (Diptera, Muscidae) in the laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef] [Green Version]
- El Boushy, A. House-fly pupae as poultry manure converters for animal feed: A review. Bioresour. Technol. 1991, 38, 45–49. [Google Scholar] [CrossRef]
- Fraenkel, G.; Bhaskaran, G. Pupariation and pupation in cyclorrhaphous flies (Diptera): Terminology and interpretation. Ann. Entomol. Soc. Am. 1973, 66, 418–422. [Google Scholar] [CrossRef]
- Martín-Vega, D.; Hall, M.J.; Simonsen, T.J. Resolving confusion in the use of concepts and terminology in intrapuparial development studies of cyclorrhaphous Diptera. J. Med. Entomol. 2016, 53, 1249–1251. [Google Scholar] [CrossRef] [Green Version]
- May, B. The occurrence in New Zealand and the life-history of the soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). N. Z. J. Sci. 1961, 4, 395–400. [Google Scholar]
- Gligorescu, A.; Toft, S.; Hauggaard-Nielsen, H.; Axelsen, J.A.; Nielsen, S.A. Development, growth and metabolic rate of Hermetia illucens larvae. J. Appl. Entomol. 2019, 143, 875–881. [Google Scholar] [CrossRef]
- Brundage, A.; Benbow, M.E.; Tomberlin, J.K. Priority effects on the life-history traits of two carrion blow fly (Diptera, Calliphoridae) species. Ecol. Entomol. 2014, 39, 539–547. [Google Scholar] [CrossRef]
- Blaustein, L.; Margalit, J. Priority effects in temporary pools: Nature and outcome of mosquito larva-toad tadpole interactions depend on order of entrance. J. Anim. Ecol. 1996, 65, 77–84. [Google Scholar] [CrossRef]
- Weslien, J.; Djupström, L.B.; Schroeder, M.; Widenfalk, O. Long-term priority effects among insects and fungi colonizing decaying wood. J. Anim. Ecol. 2011, 80, 1155–1162. [Google Scholar] [CrossRef] [Green Version]
- Palmer, T.M.; Young, T.P.; Stanton, M.L. Burning bridges: Priority effects and the persistence of a competitively subordinate acacia-ant in Laikipia, Kenya. Oecologia 2002, 133, 372–379. [Google Scholar] [CrossRef]
- Schremmer, F. The polymetabol development of the soldier fly larva Hermetia illucens—A contribution to the Metamorphosis of the Stratiomyidae. Ann. Nat. Mus. Wien. Ser. B Bot. Zool. 1984, 88, 405–429. [Google Scholar]
- Fatchurochim, S.; Geden, C.; Axtell, R. Filth fly (Diptera) oviposition and larval development in poultry manure of various moisture levels. J. Entomol. Sci. 1989, 24, 224–231. [Google Scholar] [CrossRef]
- Cammack, J.A.; Tomberlin, J.K. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Hanajima, D.; Toyoda, S.; Yoshida, N.; Morioka, R.; Osada, T. Microbiology of nitrogen cycle in animal manure compost. Microb. Biotechnol. 2011, 4, 700–709. [Google Scholar] [CrossRef]
- Lalander, C.H.; Fidjeland, J.; Diener, S.; Eriksson, S.; Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 2015, 35, 261–271. [Google Scholar] [CrossRef]
- Ong, S.-Q.; Lee, B.-B.; Tan, G.-P.; Saravanan, A.; Maniam, L. Capacity of black soldier fly and house fly larvae in treating the wasted rice in Malaysia. Malays. J. Sustain. Agric. 2017, 1, 8–10. [Google Scholar] [CrossRef]
- Kim, W.; Bae, S.; Park, K.; Lee, S.; Choi, Y.; Han, S.; Koh, Y. Biochemical characterization of digestive enzymes in the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). J. Asia Pac. Entomol. 2011, 14, 11–14. [Google Scholar] [CrossRef]
- Morrison, P.E.; Davies, D.M. Feeding of dry chemically defined diets + egg production in adult house-fly. Nature 1964, 201, 104–105. [Google Scholar] [CrossRef]
- Bertinetti, C.; Samayoa, A.C.; Hwang, S.-Y. Effects of feeding adults of Hermetia illucens (Diptera: Stratiomyidae) on longevity, oviposition, and egg hatchability: Insights into optimizing egg production. J. Insect Sci. 2019, 19, 19. [Google Scholar] [CrossRef] [Green Version]
- Bruno, D.; Bonelli, M.; Cadamuro, A.G.; Reguzzoni, M.; Grimaldi, A.; Casartelli, M.; Tettamanti, G. The digestive system of the adult Hermetia illucens (Diptera: Stratiomyidae): Morphological features and functional properties. Cell Tissue Res. 2019, 378, 221–238. [Google Scholar] [CrossRef]
- Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461. [Google Scholar] [CrossRef]
- Jiang, C.L.; Jin, W.Z.; Tao, X.H.; Zhang, Q.; Zhu, J.; Feng, S.Y.; Xu, X.H.; Li, H.Y.; Wang, Z.H.; Zhang, Z.J. Black soldier fly larvae (Hermetia illucens) strengthen the metabolic function of food waste biodegradation by gut microbiome. Microb. Biotechnol. 2019, 12, 528–543. [Google Scholar] [CrossRef] [Green Version]
- Park, S.I.; Chang, B.S.; Yoe, S.M. Detection of antimicrobial substances from larvae of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Entomol. Res. 2014, 44, 58–64. [Google Scholar] [CrossRef]
- Diener, S.; Zurbruegg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozanek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Interspecific Competition between the House Fly, Musca domestica L. (Diptera: Muscidae) and Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae) When Reared on Poultry Manure. Insects 2019, 10, 440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10120440
Miranda CD, Cammack JA, Tomberlin JK. Interspecific Competition between the House Fly, Musca domestica L. (Diptera: Muscidae) and Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae) When Reared on Poultry Manure. Insects. 2019; 10(12):440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10120440
Chicago/Turabian StyleMiranda, Chelsea D., Jonathan A. Cammack, and Jeffery K. Tomberlin. 2019. "Interspecific Competition between the House Fly, Musca domestica L. (Diptera: Muscidae) and Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae) When Reared on Poultry Manure" Insects 10, no. 12: 440. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10120440