Determining Effects of Winter Weather Conditions on Adult Amblyomma americanum (Acari: Ixodidae) Survival in Connecticut and Maine, USA


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Treatment Assignments
2.3. Weather and Snow Data
2.4. Statistical Procedures
2.4.1. Three-Way ANOVA on Adult Survival
2.4.2. Three-Way ANOVA on Female and Male Survival
2.4.3. Weather Analyses
2.4.4. Spearman Rank Order Correlation
3. Results
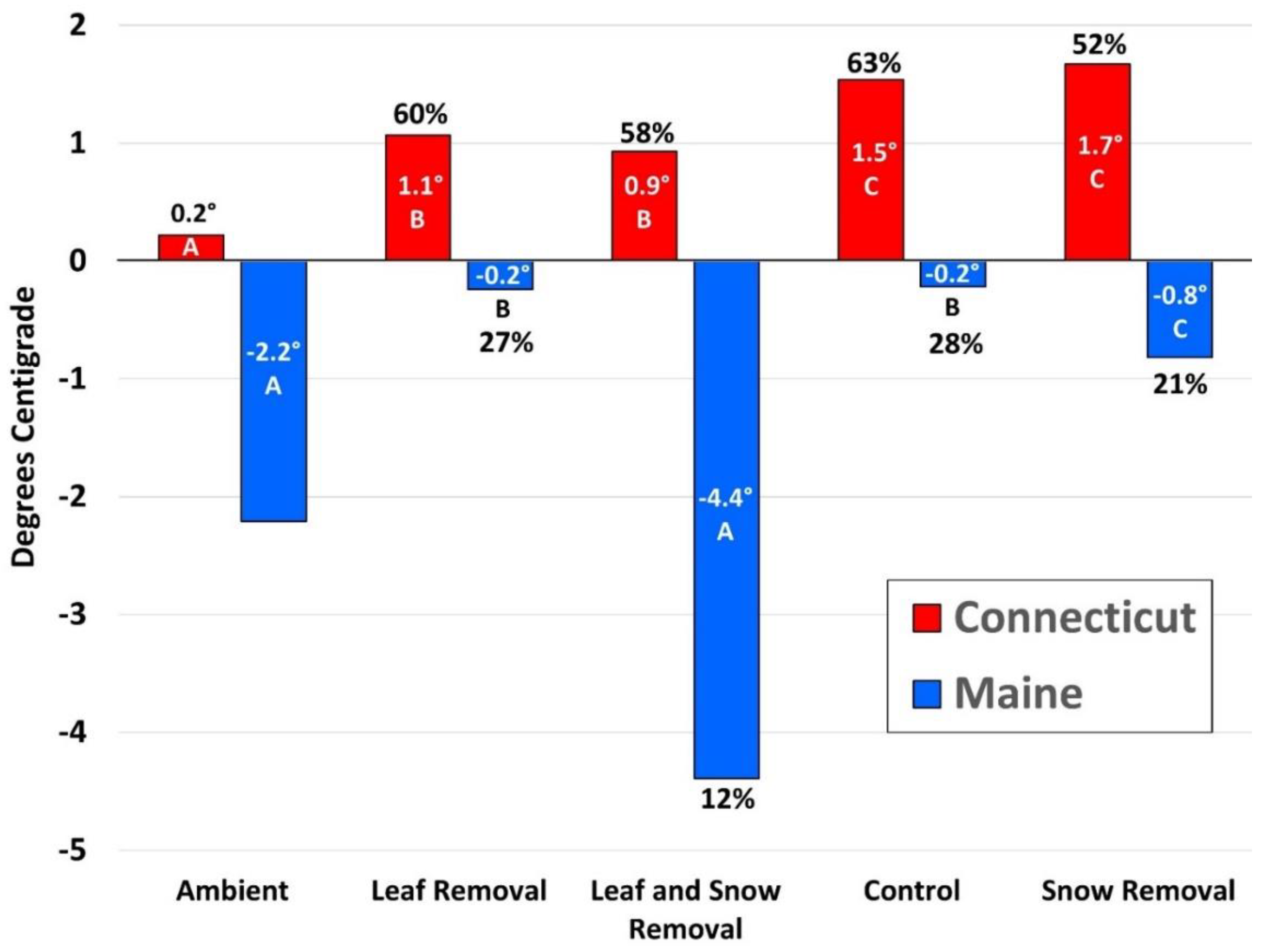
3.1. Three-Way ANOVA on Adult Survival in Connecticut
3.2. Three-Way ANOVA on Adult Survival in Maine
3.3. Three-Way ANOVA on Female Survival
3.4. Three-Way ANOVA on Male Survival
3.5. Weather Analyses
3.6. Survival and Mean Daily Temperature Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Randolph, S.E. The shifting landscape of tick-borne zoonoses: Tick-borne encephalitis and Lyme borreliosis in Europe. Philos. Trans. R. Soc. Lond. B 2001, 356, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.S.; Dautel, H.; Estrada-Peña, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Fish, D. The biological and social phenomenon of Lyme disease. Science 1993, 260, 1610–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stafford, K.C., III; DeNicola, A.J.; Kilpatrick, H.J. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) and the tick parasitoid Ixodiphagus hookeri (Hymenoptera: Encyrtidae) with reduction of white-tailed deer. J. Med. Entomol. 2003, 40, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogden, N.H.; Maarouf, A.; Barker, I.K.; Bigras-Poulin, M.; Lindsay, L.R.; Morshed, M.G.; O’Callaghan, C.J.; Ramay, F.; Waltner-Toews, D.; Charron, D.F. Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. Int. J. Parasitol. 2006, 36, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Ayllón, N.; De La Fuente, J. Impact of climate trends on tick-borne pathogen transmission. Front. Physiol. 2012, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Linske, M.A.; Stafford, K.C.; Williams, S.C.; Lubelczyk, C.B.; Welch, M.; Henderson, E.F. Impacts of deciduous leaf litter and snow presence on nymphal Ixodes scapularis (Acari: Ixodidae) overwintering survival in coastal New England, USA. Insects 2019, 10, 227. [Google Scholar] [CrossRef] [Green Version]
- Rainey, T.; Occi, J.L.; Robbins, R.G.; Egizi, A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) parasitizing a sheep in New Jersey, United States. J. Med. Entomol. 2018, 55, 757–759. [Google Scholar] [CrossRef]
- Neilson, F.J.A. An Investigation into the Ecology, Biology, Distribution and Control of Haemaphysalis longicornis Neumann, 1901. Master’s Thesis, Massey University, Palmerston North, New Zealand, 1980; p. 243. [Google Scholar]
- Heath, A. The temperature and humidity preferences of Haemaphysalis longicornis, Ixodes holocyclus and Rhipicephalus sanguineus (Ixodidae): Studies on eggs. Int. J. Parasitol. 1979, 9, 33–39. [Google Scholar] [CrossRef]
- Heath, A.C.; Palma, R.L.; Cane, R.P.; Hardwick, S. Checklist of New Zealand ticks (Acari: Ixodidae, Argasidae). Zootaxa 2011, 2995, 55–63. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, Z.; Chen, Z.; Zhou, L.; Zheng, B.; Ma, H.; Liu, J. Development and biological characteristics of Haemaphysalis longicornis (Acari: Ixodidae) under field conditions. Exp. Appl. Acarol. 2011, 53, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H.; Roberts, F.H.S.; Kohls, G.M.; Vernon, J.T. Review of Haemaphysalis (Kaiseriana) longicornis Neumann (resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and northeastern China and USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.H.; Tanaka, K.; Sawada, M. Cytogenetics of ticks (Acari: Ixodoidea). 12. Chromosome and hybridization studies of bisexual and parthenogenetic Haemaphysalis longicornis races from Japan and Korea. Chromosoma 1973, 42, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Hair, J.A.; Bowman, J.L. Behavorial ecology of Amblyomma americanum (L.). In Morpholgy, Physiology, and Behavorial Biology of Ticks; Sauer, J.R., Hair, J.A., Eds.; Wiley: New York, NY, USA, 1986; pp. 406–427. [Google Scholar]
- Ginsberg, H.S.; Ewing, C.P.; O’Connell, A.F., Jr.; Bosler, E.M.; Daley, J.G.; Sayre, M.W. Increased population densities of Amblyomma americanum (Acari: Ixodidae) on Long Island, New York. J. Parasitol. 1991, 77, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Springer, Y.P.; Eisen, L.; Beati, L.; James, A.M.; Eisen, R.J. Spatial distribution of counties in the continental United States with records of occurrence of Amblyomma americanum (Ixodida: Ixodidae). J. Med. Entomol. 2014, 51, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mather, T.N.; Mather, M.E. Intrinsic competence of three ixodid ticks (Acari) as vectors of the Lyme disease spirochetes. J. Med. Entomol. 1990, 27, 646–650. [Google Scholar] [CrossRef]
- Stafford, K.C., III.; Molaei, G.; Little, E.A.H.; Paddock, C.D.; Karpathy, S.E.; Labonte, A.M. Distribution and establishment of the lone star tick in Connecticut and implications for range expansion and public health. J. Med. Entomol. 2018, 55, 1561–1568. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Tick Borne Diseases in the United States: Tick ID. 2019. Available online: https://www.cdc.gov/ticks/tickbornediseases/tickID.html (accessed on 26 November 2019).
- Commins, S.P.; Platts-Mills, T.A. Tick bites and red meat allergy. Curr. Opin. Allergy Clin. 2013, 13, 354. [Google Scholar] [CrossRef] [Green Version]
- Van Nunen, S. Tick-induced allergies: Mammalian meat allergy, tick anaphylaxis and their significance. Asia Pac. Allergy 2015, 5, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Semtner, P.J.; Howell, D.E.; Hair, J.A. The ecology and behavior of the lone star tick (Acarina: Ixodidae) I. The relationship between vegetative habitat type and tick abundance and distribution in Cherokee Co., Oklahoma. J. Med. Entomol. 1971, 8, 329–335. [Google Scholar] [CrossRef]
- Ludwig, A.; Ginsberg, H.S.; Hickling, G.J.; Ogden, N.H. A dynamic population model to investigate effects of climate and climate-independent factors on the lifecycle of Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 99–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springer, Y.P.; Jarnevich, C.S.; Barnett, D.T.; Monaghan, A.J.; Eisen, R.J. Modeling the present and future geographic distribution of the lone star tick, Amblyomma americanum (Ixodida: Ixodidae), in the continental United States. Am. J. Trop. Med. Hyg. 2015, 93, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Needham, G.R.; Jaworski, D.C.; Chen, C.-P.; Lee, R.E., Jr. Cold-hardiness of a laboratory colony of lone star ticks (Acari: Ixodidae). J. Med. Entomol. 1966, 33, 706–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grove, D.; Fryxell, T.R.; Hickling, G.J.; Vail, K.; Ivey, J. Asian Longhorned Tick; University of Tennessee Extension Publication W826; University of Tennessee: Knoxville, TN, USA, 2019. [Google Scholar]
- Gladney, W.J.; Drummond, R. Mating behavior and reproduction of the lone star tick, Amblyomma americanum. Ann. Entomol. Soc. Am. 1970, 63, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Mufson, S.; Mooney, C.; Eilperin, J.; Muyskens, J. 2 °C: Beyond the Limit. Extreme Climate Change has Arrived in America. The Washington Post. 2019. Available online: https://www.washingtonpost.com/graphics/2019/national/climate-environment/climate-change-america/?wpisrc=nl_most&wpmm=1 (accessed on 3 October 2019).
- Molaei, G.; Little, E.A.H.; Williams, S.C.; Stafford, K.C., III. Bracing for the worst—Range expansion of the lone star tick in the northeastern United States. N. Eng. J. Med. 2019, 381, 2189–2192. [Google Scholar] [CrossRef] [PubMed]
- Haile, D.G.; Mount, G.A. Computer simulation of population dynamics of the lone star tick, Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 1987, 24, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.E.; Sims, K.G.; Olson, J.G.; Childs, J.E.; Piesman, J.F.; Harp, C.M.; Maupin, G.O.; Johnson, B.J.B. Amblyomma americanum: A potential vector of human ehrlichiosis. Am. Soc. Trop. Med. Hyg. 1993, 49, 239–244. [Google Scholar] [CrossRef]
- Goddard, J.; Varela-Stokes, A.S. Role of the lone star tick, Amblyomma americanum (L.) in human and animal diseases. Vet. Parasitol. 2009, 160, 1–12. [Google Scholar] [CrossRef]
- Pastula, D.M.; Turabelidze, G.; Yates, K.F.; Jones, T.F.; Lambert, A.J.; Panella, A.J.; Kosoy, O.I.; Velez, J.O.; Fisher, M.; Staples, E. Notes from the field: Heartland virus disease-United States, 2012–2013. MMWR 2014, 63, 270–271. [Google Scholar]


{kind=link}
| Treatment | Connecticut | Maine | |||
|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Year 1 | Year 2 | |
| Control | 56% | 69% | 78% | 7% | 73% |
| SR | 32% | 52% | 69% | 9% | 41% |
| LR | 83% | 63% | 65% | 9% | 62% |
| LRSR | 69% | 73% | 59% | 11% | 27% |
| Mean | 60% | 64% | 68% | 9% | 51% |
| Treatment | Connecticut | Maine | ||||
|---|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Year 1 | Year 2 | Year 3 | |
| Control | 46% | 69% | 61% | 13% | 40% | 18% |
| SR | 47% | 47% | 65% | 7% | 35% | 6% |
| LR | 56% | 59% | 37% | 2% | 56% | 4% |
| LRSR | 48% | 49% | 50% | 2% | 11% | 7% |
| Mean | 49% | 56% | 53% | 6% | 36% | 9% |
| Treatment | Connecticut | Maine | ||||
|---|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Year 1 | Year 2 | Year 3 | |
| Ambient | 0.83 | −1.17 | −0.40 | −1.70 | −4.19 | −3.34 |
| Control | 2.71 | 0.96 | 1.79 | −0.36 | −0.15 | −0.86 |
| SR | 2.82 | 1.10 | 1.98 | −0.65 | −1.55 | −1.35 |
| LR | 2.22 | 0.73 | 1.30 | −0.48 | −0.04 | −0.99 |
| LRSR | 2.15 | 0.59 | 1.45 | −0.86 | −1.85 | −1.54 |
| Mean | 2.14 | 0.44 | 1.22 | −0.81 | −1.56 | −1.62 |
| Location | Year | Month | # of No-Snow Days | # of Snow Days | Avg. Monthly Snow Accumulation (cm) |
|---|---|---|---|---|---|
| CT | 1 | Dec | 15 | 2 | 1.0 |
| CT | 1 | Jan | 26 | 5 | 9.2 |
| CT | 1 | Feb | 16 | 12 | 18.2 |
| CT | 2 | Dec | 12 | 5 | 2.0 |
| CT | 2 | Jan | 16 | 15 | 11.4 |
| CT | 2 | Feb | 25 | 3 | 7.2 |
| CT | 3 | Dec | 16 | 1 | 1.0 |
| CT | 3 | Jan | 25 | 6 | 2.0 |
| CT | 3 | Feb | 21 | 7 | 1.0 |
| Total | 172 | 56 | Mean = 5.9 | ||
| ME | 1 | Dec | 0 | 17 | 10.7 |
| ME | 1 | Jan | 6 | 25 | 26.4 |
| ME | 1 | Feb | 0 | 28 | 45.6 |
| ME | 2 | Dec | 0 | 17 | 20.2 |
| ME | 2 | Jan | 0 | 31 | 57.4 |
| ME | 2 | Feb | 0 | 28 | 16.1 |
| ME | 3 | Dec | 13 | 4 | 5.1 |
| ME | 3 | Jan | 5 | 26 | 10.1 |
| ME | 3 | Feb | 0 | 28 | 22.0 |
| Total | 24 | 204 | Mean = 23.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linske, M.A.; Williams, S.C.; Stafford, K.C.; Lubelczyk, C.B.; Henderson, E.F.; Welch, M.; Teel, P.D. Determining Effects of Winter Weather Conditions on Adult Amblyomma americanum (Acari: Ixodidae) Survival in Connecticut and Maine, USA. Insects 2020, 11, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010013
Linske MA, Williams SC, Stafford KC, Lubelczyk CB, Henderson EF, Welch M, Teel PD. Determining Effects of Winter Weather Conditions on Adult Amblyomma americanum (Acari: Ixodidae) Survival in Connecticut and Maine, USA. Insects. 2020; 11(1):13. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010013
Chicago/Turabian StyleLinske, Megan A., Scott C. Williams, Kirby C. Stafford, Charles B. Lubelczyk, Elizabeth F. Henderson, Margret Welch, and Peter D. Teel. 2020. "Determining Effects of Winter Weather Conditions on Adult Amblyomma americanum (Acari: Ixodidae) Survival in Connecticut and Maine, USA" Insects 11, no. 1: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010013