Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- The monophyly and delimitation of the families Keroplatidae and Lygistorrhinidae and their position within the infraorder Bibionomorpha.
- The monophyly, delimitation and interrelationships of the subfamilies and other higher taxa within Keroplatidae.
- The phylogenetic position of various problematic genera of Keroplatidae (e.g., Arachnocampa, Burmacrocera Cockerell, 1917, Chetoneura Colless, 1962, Kibaleana Kurina, Mantič and Ševčík, 2017, Paleoplatyura Meunier, 1899, Sciarokeroplatus Papp and Ševčík, 2005, Schizocyttara Matile, 1974).
- The inclusion of fossil taxa, either described or recently discovered undescribed ones, in the study of the phylogeny of Keroplatidae and Lygistorrhinidae.
2. Materials and Methods
3. Results and Discussion
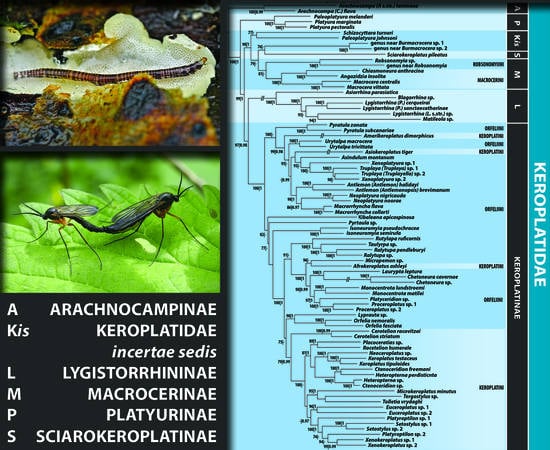
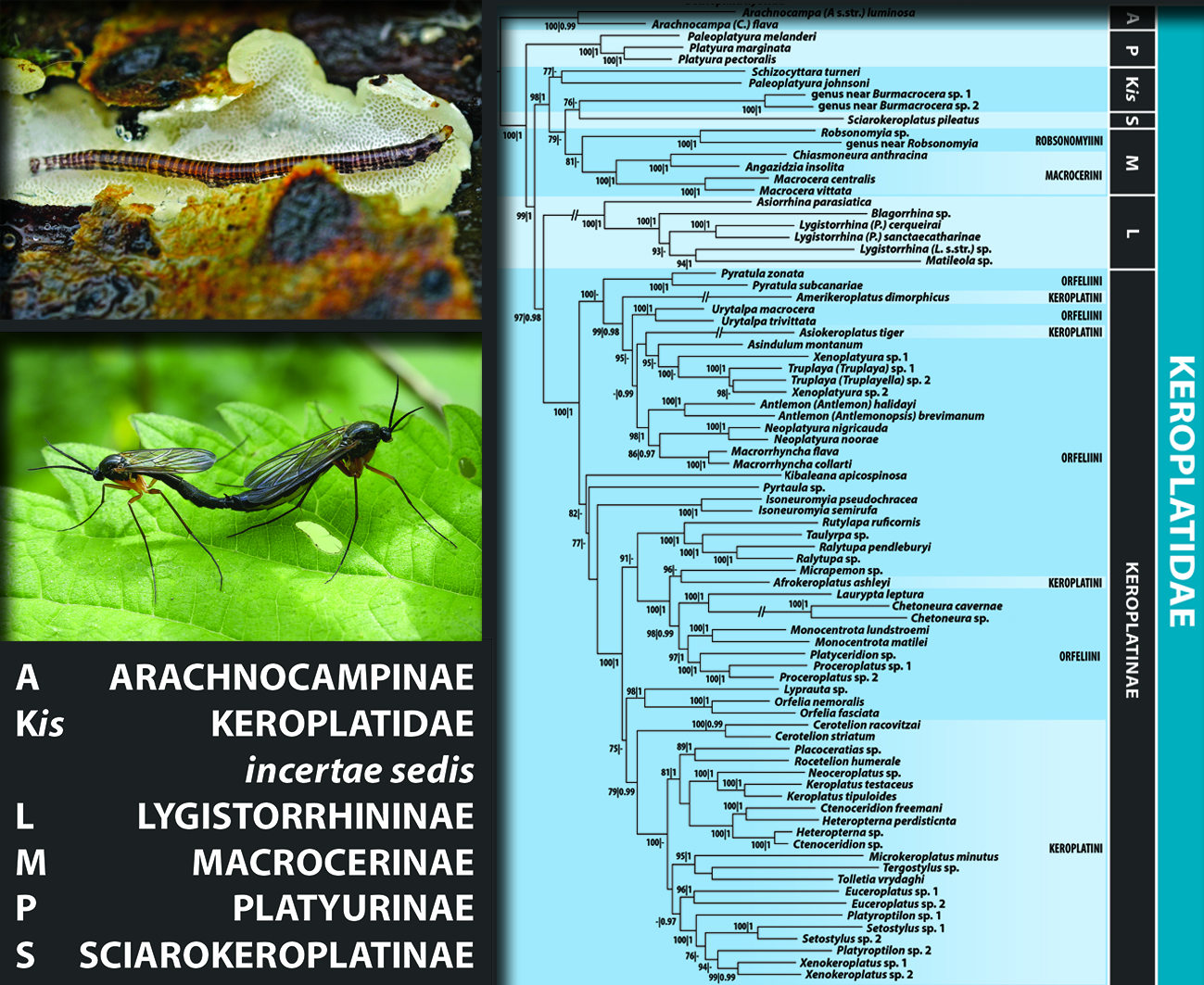
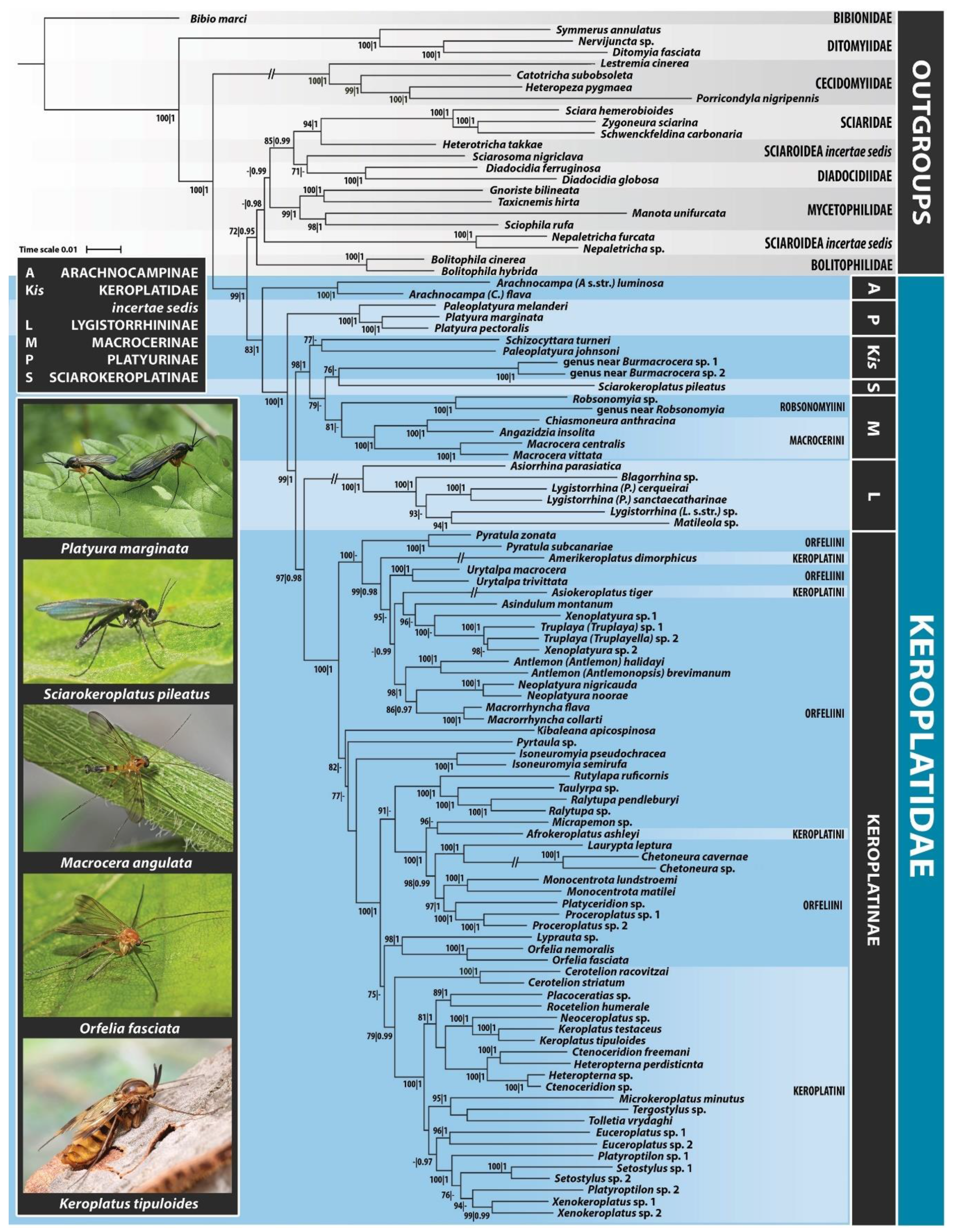
3.1. Delimitation and Monophyly of Keroplatidae
3.2. Subfamily Platyurinae Loew, 1850, Stat. Nov.
3.3. Delimitation and Monophyly of Macrocerinae and Sciarokeroplatinae
3.4. Subfamily Lygistorrhininae Edwards, 1925, Stat. Nov.
3.5. Monophyly and Interrelationships of Keroplatinae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ševčík, J.; Kaspřák, D.; Mantič, M.; Fitzgerald, S.; Ševčíková, T.; Tóthová, A.; Jaschhof, M. Molecular phylogeny of the megadiverse insect infraorder Bibionomorpha sensu lato (Diptera). PeerJ 2016, 4, e2563. [Google Scholar] [CrossRef] [Green Version]
- Matile, L. Recherches sur la systématique et lévolution des Keroplatidae: Diptera, Mycetophiloidea. Mem. Du Mus. Natl. D’histoire Nat. Ser. A Zool. 1990, 148, 1–682. [Google Scholar]
- Evenhuis, N. Catalog of The Keroplatidae of The World (Insecta: Diptera); Bishop Museum Press: Honolulu, HI, USA, 2006. [Google Scholar]
- Ševčík, J.; Mantič, M.; Blagoderov, V. Two new genera of Keroplatidae (Diptera), with an updated key to the World genera of Keroplatini. Acta Entomol. Musei Natl. Pragae 2015, 55, 387–399. [Google Scholar]
- Kurina, O.; Mantič, M.; Ševčík, J. A Remarkable New Genus of Keroplatidae (Insecta, Diptera) from the Afrotropical Region, with DNA Sequence Data. Afr. Invertebr. 2017, 58, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Fungus Gnats Online. Available online: http://sciaroidea.info/ (accessed on 22 April 2020).
- Evenhuis, N. Catalogue of The Fossil Flies of the World (Insecta: Diptera); Backhuys: Leiden, The Netherlands, 1994. [Google Scholar]
- Papp, L.; Ševčík, J. Sciarokeroplatinae, a new subfamily of Keroplatidae (Diptera). Acta Zool. Acad. Sci. Hung. 2005, 51, 113–123. [Google Scholar] [CrossRef]
- Blagoderov, V.; Grimaldi, D. Fossil Sciaroidea (Diptera) In Cretaceous Ambers, Exclusive of Cecidomyiidae, Sciaridae, and Keroplatidae. Am. Mus. Novit. 2004, 3433. [Google Scholar] [CrossRef]
- Grimaldi, D.; Amorim, D.D.S.; Blagoderov, V. The Mesozoic Family Archizelmiridae (Diptera: Insecta). J. Paleontol. 2003, 77, 368–381. [Google Scholar] [CrossRef]
- Baker, C.; Graham, G.; Scott, K.; Cameron, S.; Yeates, D.; Merritt, D. Distribution and Phylogenetic Relationships of Australian Glow-Worms Arachnocampa (Diptera, Keroplatidae). Mol. Phylogenet. Evol. 2008, 48, 506–514. [Google Scholar] [CrossRef]
- Saigusa, T. Rare Sciaroid Genera from the Temperate east Palaearctic Region (Diptera: Sciaroidea). Jpn. J. Syst. Entomol. 2018, 24, 163–173. [Google Scholar]
- Bechev, D. Flower visitation of fungus gnats from the genera Antlemon, Asindulum and Macrorrhyncha (Diptera: Keroplatidae): Published data and a new record. ZooNotes 2010, 7, 1–3. [Google Scholar]
- Mantič, M.; Ševčík, J. Macrocera rohaceki sp. nov. and other interesting records of Keroplatidae (Diptera) from southern and central Europe, with DNA sequence data. Acta Entomol. Musei Natl. Pragae 2017, 57, 751–764. [Google Scholar] [CrossRef] [Green Version]
- Reiter, N.; Freestone, M.; Brown, G.; Peakall, R. Pollination by sexual deception of fungus gnats (Keroplatidae and Mycetophilidae) in two clades of Pterostylis (Orchidaceae). Bot. J. Linn. Soc. 2019, 190, 101–116. [Google Scholar] [CrossRef]
- Matile, L. Phylogeny and evolution of the larval diet in the Sciaroidea (Diptera, Bibionomorpha) since the Mesozoic. In The Origin of Biodiversity in Insects: Phylogenetic Tests of Evolutionary Scenarios, 2nd ed.; Memoires du Museum National d’Histoire Naturelle Seri, A Zoologie: Paris, France, 1997; Volume 173, pp. 273–303. [Google Scholar]
- Sharpe, M.L.; Dearden, P.K.; Gimenez, G.; Krause, K.L. Comparative RNA seq analysis of the New Zealand glowworm Arachnocampa luminosa reveals bioluminescence-related genes. BMC Genom. 2015, 16, 825. [Google Scholar] [CrossRef] [Green Version]
- Falaschi, R.; Amaral, D.; Santos, I.; Domingos, A.; Johnson, G.; Martins, A.; Viroomal, I.; Pompéia, S.; Mirza, J.; Oliveira, A.; et al. Neoceroplatus betaryiensis nov. sp. (Diptera: Keroplatidae) is the first record of a bioluminescent fungus-gnat in South America. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Aiello, A.; Jolivet, P. Myrmecophily in Keroplatidae (Diptera: Sciaroidea). J. N. Y. Entomol. Soc. 1996, 104, 226–230. [Google Scholar]
- Chandler, P.J.; Matile, L. A new species of Platyceridion Tollet (Diptera, Keroplatidae) with a larva predatory in ant infested internodes of Humboldtia laurifolia Vahl. Studia Dipterol. 1998, 5, 163–173. [Google Scholar]
- Chandler, P.J.; Pijnakker, J. Tropical fungus gnats established in nurseries in The Netherlands (Diptera: Keroplatidae) and Mycetophilidae. Br. J. Entomol. Nat. Hist. 2009, 22, 81. [Google Scholar]
- Hickman, V.V. On Planarivora insignis gen. et sp. n. (Diptera: Mycetophilidae), whose larval stages in land planarians are parasitic. Pap. Proc. R. Soc. Tasman. 1965, 99, 1–8. [Google Scholar]
- Bertone, M. Field Observations of Lygistorrhina sanctaecatharinae Thompson (Diptera: Sciaroidea). Proc. Entomol. Soc. Wash. 2018, 120, 448–451. [Google Scholar] [CrossRef]
- Edwards, F. British Fungus-Gnats (Diptera, Mycetophilidae). With A Revised Generic Classification of the Family. Trans. R. Entomol. Soc. Lond. 1925, 73, 505–670. [Google Scholar] [CrossRef]
- Tuomikoski, R. Systematic position of Lygistorrhina Skuse (Diptera, Mycetophiloidea). Ann. Entomol. Fenn. 1966, 32, 254–260. [Google Scholar]
- Hennig, W. Ordnung Diptera (Zweiflügler). Handb. Der Zool. 1973, 4, 1–337. [Google Scholar]
- Søli, G.E.E.; Vokeroth, J.R.; Matile, L. Families of Sciaroidea. In Contributions to A Manual of Palaeartic Diptera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 2000; pp. 49–92. [Google Scholar]
- Amorim, D.D.S.; Rindal, E. Phylogeny of the Mycetophiliformia, with proposal of the subfamilies Heterotrichinae, Ohakuneinae, and Chiletrichinae for the Rangomaramidae (Diptera, Bibionomorpha). Zootaxa 2007, 1535, 1–92. [Google Scholar] [CrossRef]
- Hippa, H.; Vilkamaa, P. Phylogeny of the Sciaroidea (Diptera): The Implication of additional taxa and character data. Zootaxa 2006, 1132, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic radiations in the fly tree of life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [Green Version]
- Vilkamaa, P.; Hippa, H. The genus Sciarotricha gen. n. (Sciaridae) and the phylogeny of recent and fossil Sciaroidea (Diptera). Insect Syst. Evol. 2005, 36, 121–143. [Google Scholar] [CrossRef] [Green Version]
- Bertone, M.; Courtney, G.; Wiegmann, B. Phylogenetics and temporal diversification of the earliest true flies (Insecta: Diptera) based on multiple nuclear genes. Syst. Entomol. 2008, 33, 668–687. [Google Scholar] [CrossRef]
- Rindal, E.; Søli, G.E.; Bachmann, L. Molecular phylogeny of the fungus gnat family Mycetophilidae (Diptera, Mycetophiliformia). Syst. Entomol. 2009, 34, 524–532. [Google Scholar] [CrossRef]
- Kallweit, U. Lygistorrhina Skuse has branched from the Keroplatidae (Diptera: Sciaroidea)—A new perspective on the phylogenetic relationships in the keroplatid group of fungus gnats. In Proceedings of the 8th International Congress of Dipterology, Potsdam, Germany, 10–15 August 2014; Volume xxvii, p. 156. [Google Scholar]
- Thompson, F.C. Notes on the genus Lygistorrhina Skuse with the description of the first Nearctic species (Diptera: Mycetophiloidea). Proc. Entomol. Soc. Wash. 1975, 77, 434–445. [Google Scholar]
- Kaspřák, D.; Kerr, P.; Sýkora, V.; Tóthová, A.; Ševčík, J. Molecular phylogeny of the fungus gnat subfamilies Gnoristinae and Mycomyinae, and their position within Mycetophilidae (Diptera). Syst. Entomol. 2019, 44, 128–138. [Google Scholar] [CrossRef]
- Sikora, T.; Jaschhof, M.; Mantič, M.; Kaspřák, D.; Ševčík, J. Considerable congruence, enlightening conflict: Molecular analysis largely supports morphology-based hypotheses on Cecidomyiidae (Diptera) phylogeny. Zool. J. Linn. Soc. 2019, 185, 98–110. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.; Lemey, P. Assessing substitution saturation with DAMBE. Phylogenet. Handb. Pract. Approach DNA Protein Phylogeny 2009, 2, 615–630. [Google Scholar]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Among-site rate variation and its impact on phylogenetic analyses. Trends Ecol. Evol. 1996, 11, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Haeseler, A.V.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 gateway computing environments workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, 475–478. [Google Scholar] [CrossRef]
- Edwards, F.W. A note on the “New Zealand glow-worm” (Diptera, Mycetophilidae). Ann. Mag. Nat. Hist. 1924, 14, 175–179. [Google Scholar] [CrossRef]
- Sabrosky, C.W. Family-group names in Diptera: An annotated catalog. Myia 1999, 10, 1–360. [Google Scholar]
- Soós, Á.; Papp, L. Catalogue of Palaearctic Diptera: Ceratopogonidae-Mycetophilidae; Elsevier: Amsterdam, The Netherlands, 1988. [Google Scholar]
- Kurina, O. Fungus Gnats in Estonia: (Diptera: Bolitophilidae, Keroplatidae, Macroceridae, Ditomyiidae, Diadocidiidae, Mycetophilidae); Tartu University Press: Tartu, Estonia, 1998. [Google Scholar]
- Polevoi, A.V. Fungus Gnats (Diptera: Bolitophilidae, Ditomyiidae, Keroplatidae, Diadocidiidae, Mycetophilidae) of Karelia; Karelian Research Centre, Russian Academy of Sciences: Petrozavodsk, Russia, 2000. [Google Scholar]
- Papp, L. New record of Diptera species from Hungary, with the list of the Hungarian Scathophagidae. Folia Entomol. Hung. 2006, 67, 121–128. [Google Scholar]
- Papp, L. Dixidae, Axymyiidae, Mycetobiidae, Keroplatidae, Macroceridae, and Ditomyiidae(Diptera) from Taiwan. Acta Zool. Acad. Sci. Hung. 2007, 53, 273–294. [Google Scholar]
- Ševčík, J. Langkawiana maculata gen. et sp. n. from Malaysia and its systematic position in the tribe Robsonomyiini (Diptera: Keroplatidae). Zootaxa 2009, 2221, 58–66. [Google Scholar] [CrossRef]
- Amorim, D.D.S.; Niu, C.; Li, X.; Lei, C.; Clarke, A. Chetoneura shennonggongensis, a new species of cave-dwelling Keroplatini from China (Diptera: Keroplatidae), with a discussion of the position of Chetoneura. Zootaxa 2008, 1716, 59–68. [Google Scholar] [CrossRef]
- Blagoderov, V. Lygistorrhinidae (Long-beaked Fungus Gnats). In Manual of Afrotropical Diptera. Nematocerous Diptera and lower Brachycera, 2nd ed.; Kirk–Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 2, pp. 527–531. [Google Scholar]
- Hippa, H.; Mattsson, I.; Vilkamaa, P. New Taxa of the Lygistorrhinidae (Diptera: Sciaroidea) and Their Implications for a Phylogenetic Analysis of the Family. Zootaxa 2005, 960, 1–34. [Google Scholar] [CrossRef]
- Blagoderov, V.; Hippa, H.; Ševčík, J. Asiorrhina, a new Oriental genus of Lygistorrhinidae (Diptera: Sciaroidea) and its phylogenetic position. Zootaxa 2009, 2295, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Matile, L. Revision des Truplaya Afrotropicaux (Diptera, Mycetophilidae). Ann. De La Société Entomol. De Fr. 1978, 14, 451–477. [Google Scholar]
- Blagoderov, V.; Ševčík, J. Keroplatidae (Predaceous Fungus Gnats). In Manual of Afrotropical Diptera. Nematocerous Diptera and lower Brachycera, 2nd ed.; Kirk–Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 2, pp. 527–531. [Google Scholar]
- Matile, L. Rocetelion, a new Holarctic genus of the Keroplatidae (Diptera, Mycetophiloidea): Description, phylogenetic and biogeographic notes. Ann. Entomol. Fenn. 1988, 54, 107–113. [Google Scholar]
- Uesugi, K. Fungus gnats of the genus Urytalpa Edwards (Diptera: Keroplatidae) in Japan. Entomol. Sci. 2004, 7, 369–376. [Google Scholar] [CrossRef]
- Kjærandsen, J.; Martinsson, S.; Hedmark, K.; Evenhuis, N.L. On the genus Urytalpa Edwards (Diptera: Keroplatidae) in the Nordic and Nearctic Regions, with fixation of a new type species and a key to World males. Zootaxa 2009, 2160, 29–50. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, S. A Curious new genus of Keroplatini (Diptera: Keroplatidae) from Guatemala. Zootaxa 2019, 4586, 383. [Google Scholar] [CrossRef]
- Chandler, P.J.; Blasco-Zumeta, J. The fungus gnats (Diptera, Bolitophilidae, Keroplatidae and Mycetophilidae) of the Monegros region (Zaragoza, Spain) and five other new European species of Pyratula Edwards and Sciophila Meigen. Zapateri. Rev. Aragonesa De Entomol. 2001, 9, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Kovac, D.; Matile, L. Truplaya ferox (Insecta: Diptera: Mycetophiloidea), a new Malaysian keroplatid from bamboo phytotelmata with larvae predaceous on ants. Raffles Bull. Zool. 1997, 45, 15–28. [Google Scholar]
- Matile, L. Diptera: Mycetophilidae Keroplatinae. S. Afr. Anim. Life 1974, 15, 1–532. [Google Scholar]
- Ševčík, J. Terocelion gen. nov., a new Oriental genus of Keroplatidae (Diptera) with pectinate antennae. Acta Entomol. Musei Natl. Pragae 2012, 52, 495–503. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantič, M.; Sikora, T.; Burdíková, N.; Blagoderov, V.; Kjærandsen, J.; Kurina, O.; Ševčík, J. Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification. Insects 2020, 11, 348. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060348
Mantič M, Sikora T, Burdíková N, Blagoderov V, Kjærandsen J, Kurina O, Ševčík J. Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification. Insects. 2020; 11(6):348. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060348
Chicago/Turabian StyleMantič, Michal, Tomáš Sikora, Nikola Burdíková, Vladimir Blagoderov, Jostein Kjærandsen, Olavi Kurina, and Jan Ševčík. 2020. "Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification" Insects 11, no. 6: 348. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11060348