Biological Notes and Distribution in Southern Europe of Aclees taiwanensis Kȏno, 1933 (Coleoptera: Curculionidae): A New Pest of the Fig Tree

 ,
,  , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
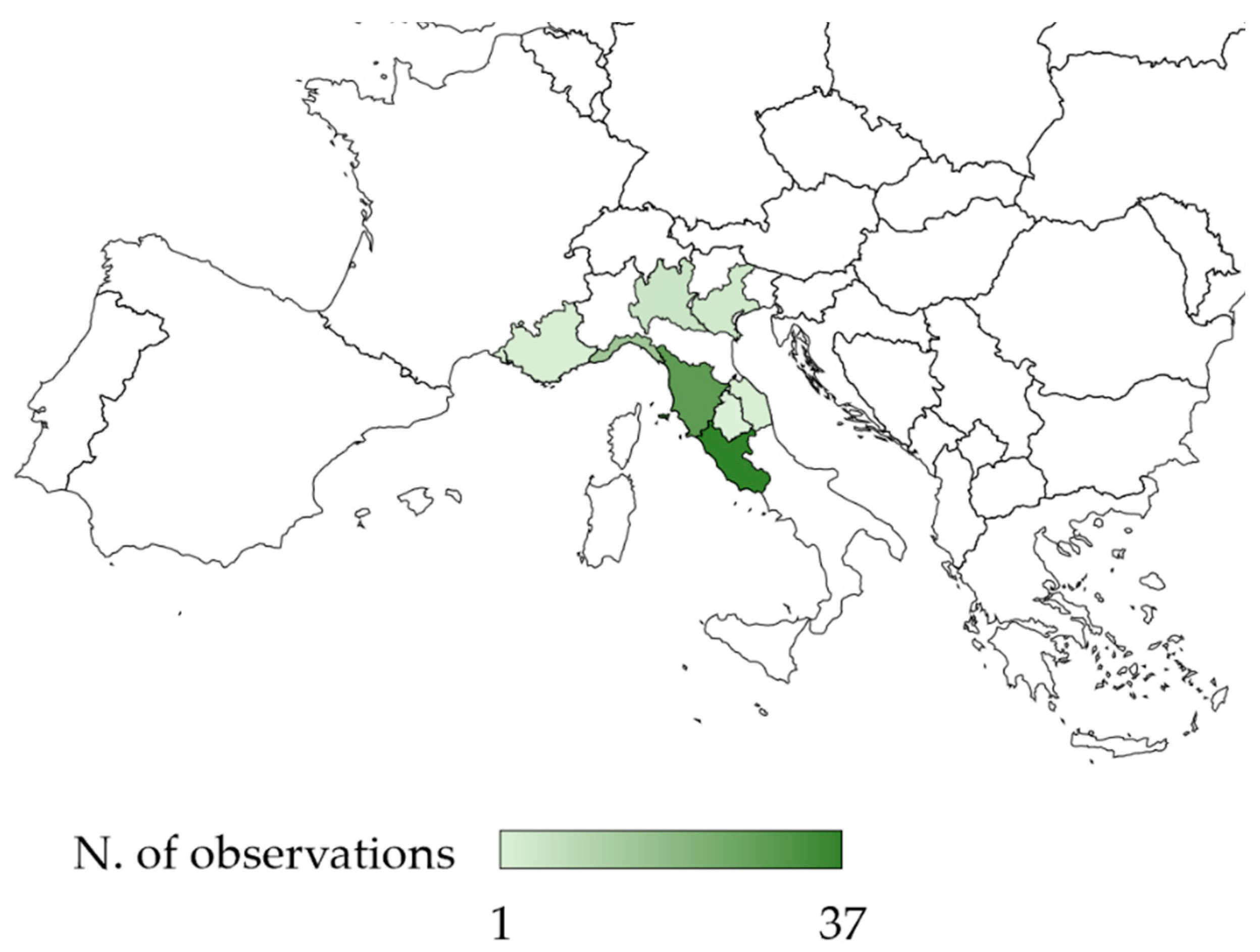
2.1. Distribution
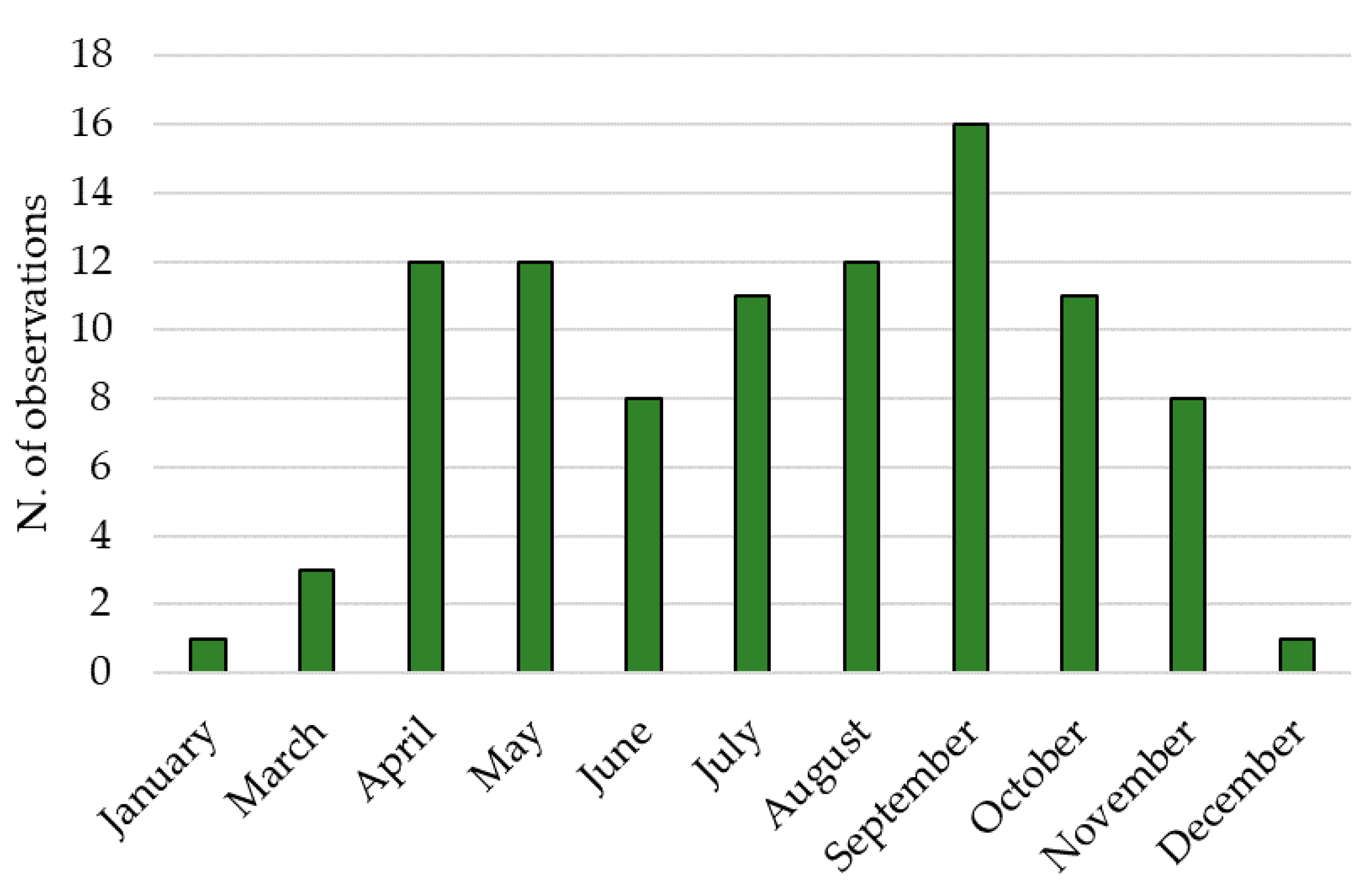
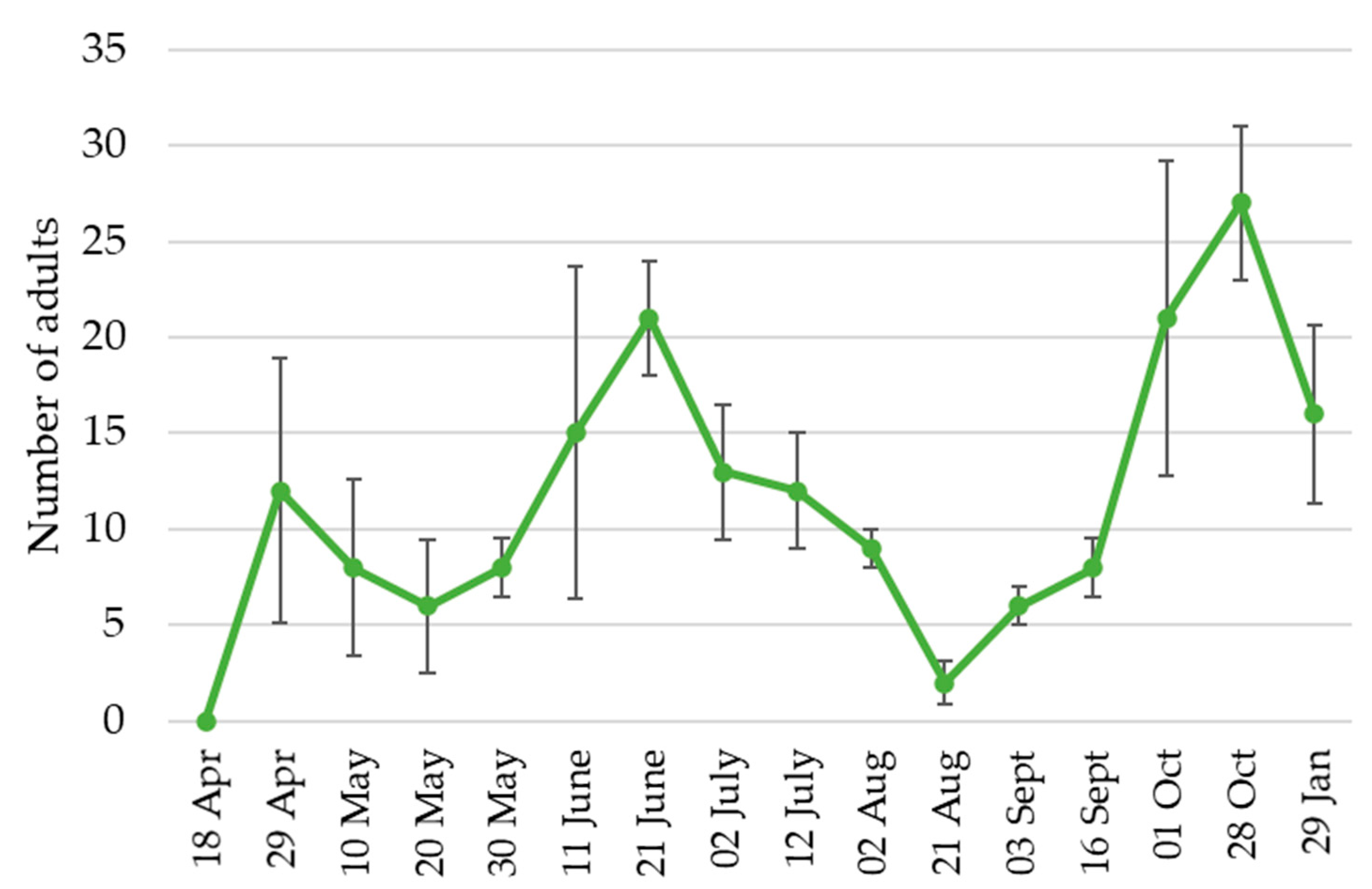
2.2. Population Dynamics in the Field
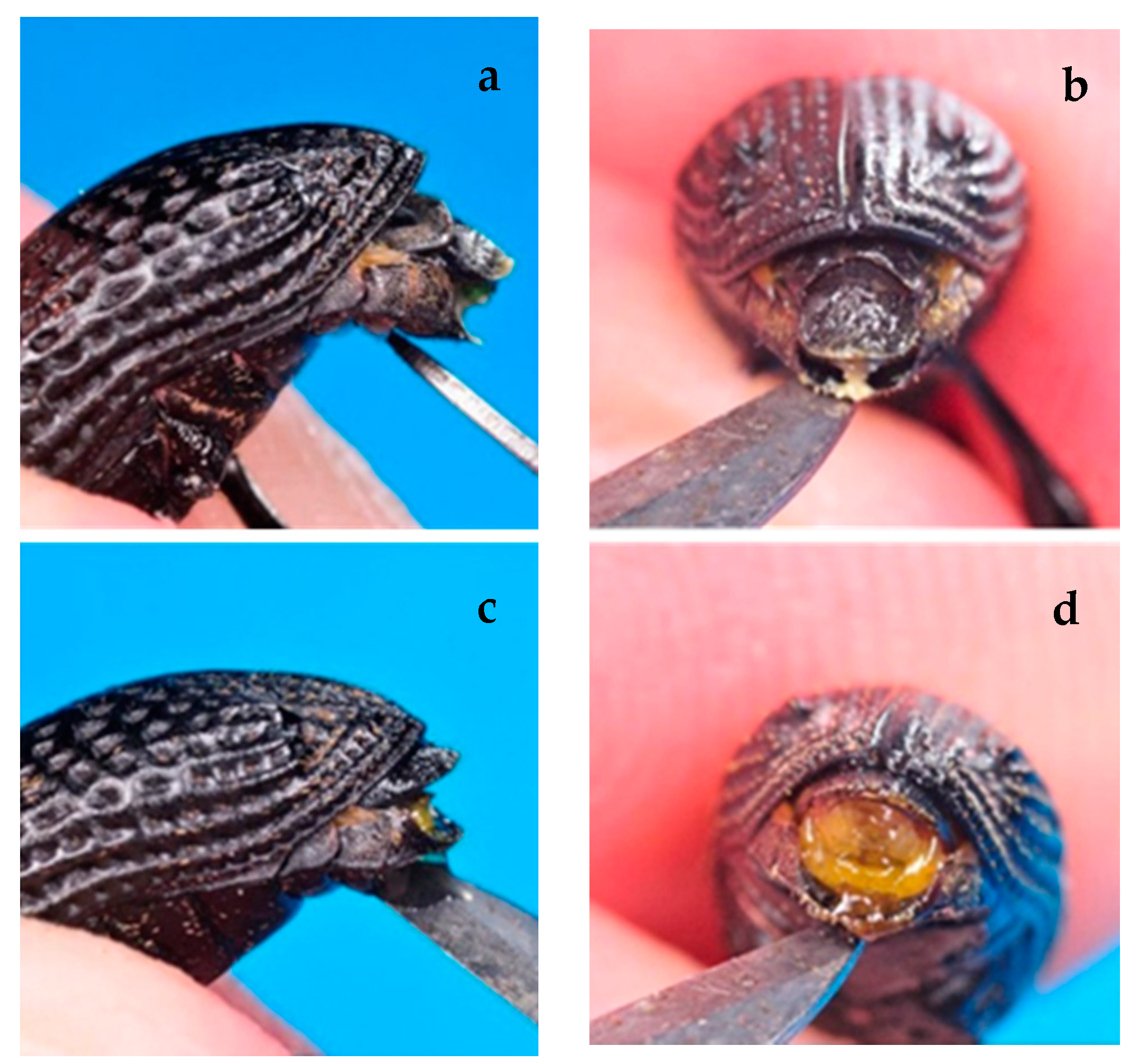
2.3. Sexual Dimorphism Determination
2.4. Insects Rearing
2.5. Female Fecundity and Fertility
2.6. Preimaginal Instars
2.7. Host Plant Species
2.8. Data Analysis
3. Results
3.1. Distribution
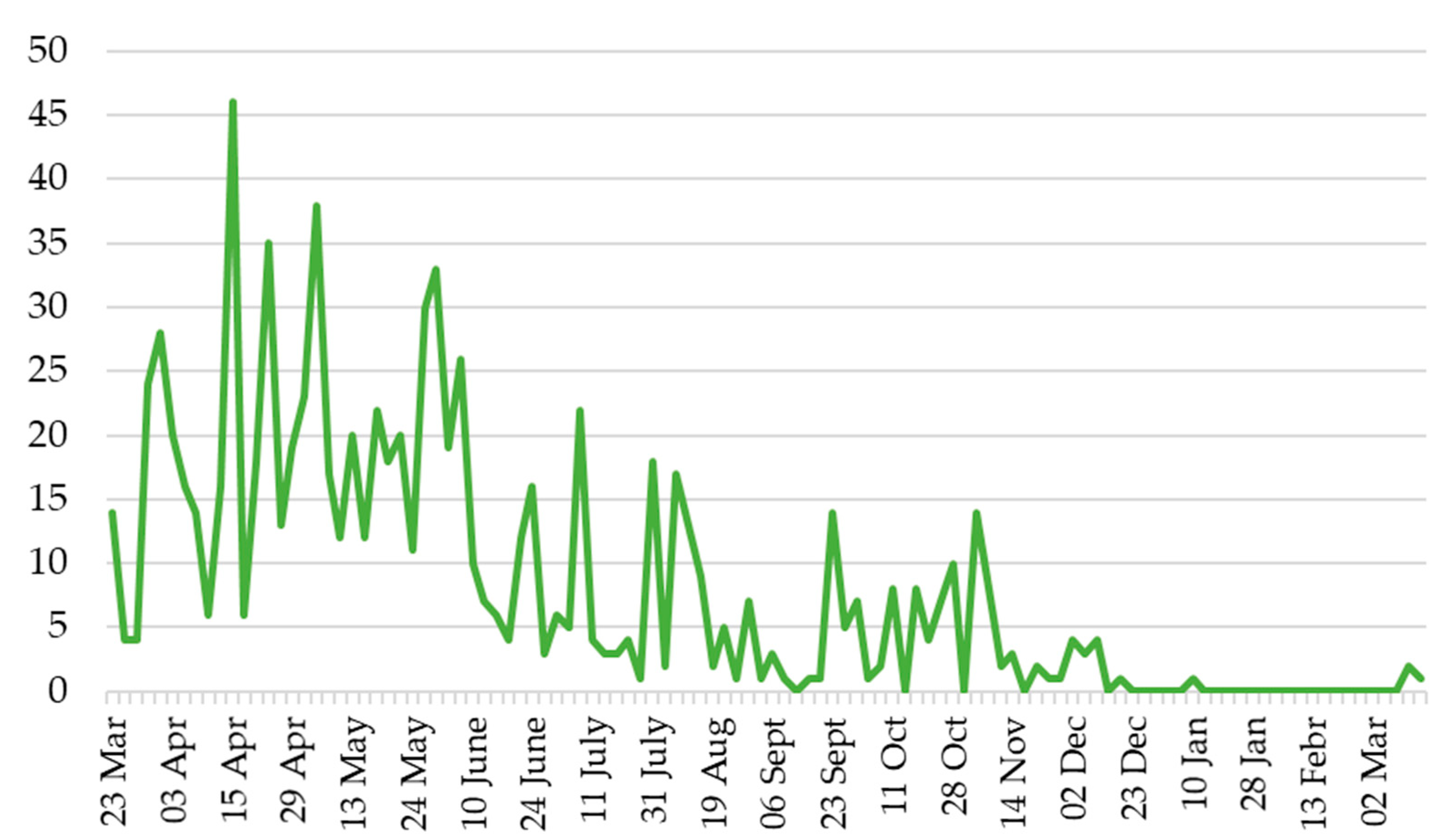
3.2. Population Dynamics in the Field
3.3. Sexual Dimorphism Determination
3.4. Female Fecundity and Fertility
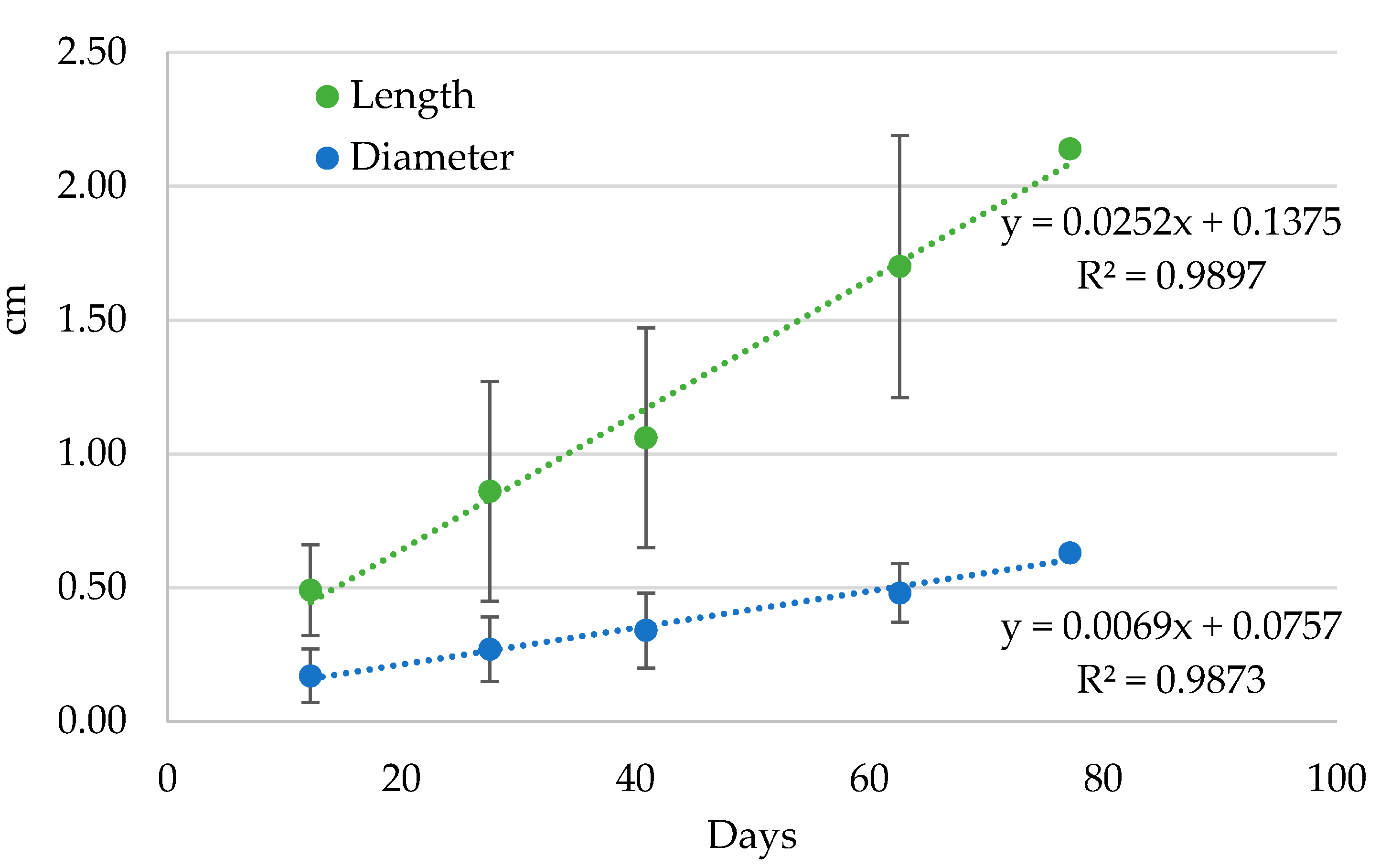
3.5. Preimaginal Instars
3.6. Host Plant Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diagne, C.; Leroy, B.; Gozlan, R.E.; Vaissière, A.C.; Assailly, C.; Nuninger, L.; Roiz, D.; Jourdain, F.; Jarić, I.; Courchamp, F. InvaCost, a public database of the economic costs of biological invasions worldwide. Sci. Data 2020, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H.; Mahajan, R.T. Traditional uses, phytochemistry and pharmacology of Ficus carica: A review. Pharm. Biol. 2014, 52, 1487–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, A.; Golubowicz, S.; Yablowicz, Z.; Grossman, S.; Bergman, M.; Gottlieb, H.E.; Altman, A.; Kerem, Z.; Flaishman, M.A. Antioxidant activities and anthocyanin content of fresh fruits of common fig (Ficus carica L.). J. Agric. Food Chem. 2006, 54, 7717–7723. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Soukara, S.; Vasilopoulou, E. Traditional foods: A science and society perspective. Trends Food Sci. Technol. 2007, 18, 420–427. [Google Scholar] [CrossRef]
- Guarrera, P.M. Traditional phytotherapy in Central Italy (Marche, Abruzzo, and Latium). Fitoterapia 2005, 76, 1–25. [Google Scholar] [CrossRef]
- Meregalli, M.; Boriani, M.; Taddei, A.; Hsu, C.F.; Tseng, W.Z.; Mouttet, R. A new species of Aclees from Taiwan with notes on other species of the genus (Coleoptera: Curculionidae: Molytinae). Zootaxa 2020, 4868, 1–26. [Google Scholar] [CrossRef]
- Perrin, H. Récoltes accidentelles de Curculionidae tropicaux en France métropolitaine (Coleoptera). Entomologiste 1997, 53, 155–158. [Google Scholar]
- Ciampolini, M.; Perrin, H.; Regalin, R. Aclees cribratus, nuovo per l’Italia nocivo al fico allevato in vivaio. Inf. Agrar. 2005, 47, 69–72. [Google Scholar]
- Benelli, G.; Meregalli, M.; Canale, A. Field observations on the mating behavior of Aclees sp. cf. foveatus Voss (Coleoptera: Curculionidae), an exotic pest noxious to fig orchards. J. Insect Behav. 2014, 27, 419–427. [Google Scholar] [CrossRef]
- Gargani, E.; Mazza, G.; Benvenuti, C.; Torrini, G.; Strangi, A.; Pennacchio, F.; Roversi, P.F. Biological control of Aclees sp. cf. foveatus and first recovery of an associate Beauveria bassiana strain. J. Zool 2016, XCIX, 29–33. [Google Scholar] [CrossRef]
- Gargani, E.; Simoni, S.; Benvenuti, C.; Frosinini, R.; Barzanti, G.P.; Roversi, P.R.; Caselli, A.; Guidotti, M. Aclees cf. sp. foveatus (Coleoptera Curculionidae), an exotic pest of Ficus carica in Italy: A sustainable approach to defence based on aluminosilicate minerals as host plant masking solids. J. Zool 2018, 101, 201–205. [Google Scholar] [CrossRef]
- EPPO Insect Pests Recorded for the First Time in Italy. Reporting Service No. 04. Num. Article: 2009/071. Available online: https://gd.eppo.int/reporting/article-175 (accessed on 17 December 2020).
- Ciampolini, M.; Regalin, R.; Farnesi, I.; Lorenzi, C. Prime osservazioni sulla bio-etologia di Aclees sp. (Curculionidae, Molytinae) esiziale a Ficus carica L. in Italia. Boll. Zool. Agrar. Bachicolt. 2007, 39, 51–60. [Google Scholar]
- Ciampolini, M.; Farnesi, I.; Scarselli, F.; Lorenzi, C. Contro il curculionide del fico decisiva la lotta agli adulti. Inf. Agrar. 2008, 25, 57–60. [Google Scholar]
- Gargani, E.; Cutino, I.; Barzanti, G.P.; Benvenuti, C.; Lodolini, E.M.; Nolasco, A.; Caboni, E.; Macchioni, V.; Carbone, K. The black weevil (Aclees sp. cf. foveatus) of the fig tree: Control trials with plant extracts. In Proceedings of the Book of Abstract of the VI International Symposium on Fig, Rovinij, Croatia, 2–5 September 2019. [Google Scholar]
- Mouttet, R.; Haran, J.; Boriani, M.; Meregalli, M.; Taddei, A.; Panchaud, K.; Vernier, F.; Streito, J.C. Aclees sp. ravageur des Figuiers établi en France métropolitaine (Coleoptera Curculionidae). Entomologiste 2020, 76, 65–68. [Google Scholar]
- Iovinella, I.; Pierattini, E.C.; Bedini, S.; Dani, F.R.; Guarino, S.; Lucchi, A.; Giannotti, P.; Cuzzupoli, G.; Girardi, J.; Conti, B. Semiochemicals for intraspecific communication of the fig weevil Aclees sp. cf. foveatus (Coleoptera:Curculionidae): A first survey. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Forum Entomologi Italiani. Available online: http://www.entomologiitaliani.net (accessed on 30 September 2020).
- Forum Natura Mediterraneo. Available online: https://www.naturamediterraneo.com/forum/ (accessed on 30 September 2020).
- iNaturalist. Available online: http://www.inaturalist.org (accessed on 30 September 2020).
- Gargani, E.; Barzanti, G.P.; Strangi, A.; Mazza, G.; Benvenuti, C.; Frosinini, R.; Roversi, P.F.; Cutino, I. Aclees sp. cf. foveatus, a real threat to Ficus carica in the Mediterranean area. In Proceedings of the Book of Abstract of the VI International Symposium on Fig, Rovinij, Croatia, 2–5 September 2019. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Roques, A.; Rabitsch, W.; Rasplus, J.Y.; Lopez-Vaamonde, C.; Nentwig, W.; Kenis, M. Alien Terrestrial Invertebrates of Europe. In DAISIE, Handbook of Alien Species in Europe; Springer: Berlin/Heidelberg, Germany, 2009; pp. 63–81. [Google Scholar]
- Inghilesi, A.F.; Mazza, G.; Cervo, R.; Gherardi, F.; Sposimo, P.; Tricarico, E.; Zapparoli, M. Alien insects in Italy: Comparing patterns from the regional to European level. J. Insect Sci. 2013, 13, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAOSTAT. Timerange: 2015–2018; Groups: Production; Domains: Crops; Country: Italy; Element: Production Quantity and Area harvested; Item: Figs. Available online: http://www.fao.org/faostat/en/#compare (accessed on 17 December 2020).
- Leather, S.R.; Day, K.R.; Salisbury, A.N. The biology and ecology of the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae): A problem of dispersal? Bull. Entomol. Res. 1999, 89, 3–16. [Google Scholar] [CrossRef]
- Adams, D.C.; Funk, D.J. Morphometric inferences on sibling species and sexual dimorphism in Neochlamisus bebbianae leaf beetles multivariate applications of the thin-plate pline. Syst. Biol. 1997, 46, 180–194. [Google Scholar] [CrossRef]
- Starr, C.K.; Wilson, D.D.; Severson, R.F.; Kays, S.J. Sexual dimorphism in the sweet potato weevil, Clymas formicarius (Coleoptera, Brentidae). Can. Entomol. 1997, 129, 61–69. [Google Scholar] [CrossRef]
- Schöps, K.; Wratten, S.D.; Emberson, R.M. Life cycle, behaviour and conservation of the large endemic weevil, Hadramphus spinipennis on the Chatham Islands, New Zealand. N. Z. J. Zool. 1999, 26, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Mori, E.; Mazza, G.; Lovari, S. Sexual dimorphism. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shakelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–7. [Google Scholar]
- Morimoto, K. The family Curculionidae of Japan. I. Subfamily Hylobiinae. Esakia 1982, 19, 51–121. [Google Scholar]
- Thu, P.Q.; Quang, D.N.; Dell, B. Threat to cedar, Cedrela odorata, plantations in Vietnam by the weevil, Aclees sp. J. Insect Sci. 2010, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Gold, C.S.; Pena, J.E.; Karamura, E.B. Biology and integrated pest management for the banana weevil Cosmopolites sordidus (Germar) (Coleoptera: Curculionidae). Integr. Pest Manag. Rev. 2001, 6, 79–155. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Rostrum Length | Body Length | Body Width |
|---|---|---|---|
| Female | 6.7 ± 1.0 | 19.8 ± 1.0 | 8.3 ± 0.3 |
| Male | 5.6 ± 0. 5 | 18.0 ± 1.0 | 7.9 ± 0.5 |
| Couple | Eggs Laid | % in the Ground | % in the Branches | Hatched Eggs | % Hatched Eggs |
|---|---|---|---|---|---|
| 1 | 75 | 39 | 61 | 53 | 71 |
| 2 | 58 | 38 | 62 | 46 | 79 |
| 3 | 126 | 74 | 26 | 76 | 60 |
| 4 | 147 | 51 | 49 | 106 | 72 |
| 5 | 186 | 50 | 50 | 124 | 67 |
| 6 | 105 | 48 | 52 | 59 | 55 |
| Total | 697 | 52 | 48 | 464 | 66 |
| Average | 116.17 ± 47.17 | 49.84 ± 13.02 | 50.16 ± 13.02 | 77.33 ± 31.34 | 67.33 ± 8.69 |
| N. of Eggs/Specimens | Instar | Days a |
|---|---|---|
| 356 | Embryonic | 9.95 ± 1.71 |
| 37 | L1 | 12.22 ± 6.41 |
| 29 | L2 | 15.34 ± 7.72 |
| 17 | L3 | 13.35 ± 7.26 |
| 14 | L4 | 21.71 ± 7.24 |
| 10 | L5 | 14.56 ± 8.13 |
| 4 | Pupae | 23.25 ± 2.16 |
| Total | 110.38 |
| Instar | Dimensions | Average |
|---|---|---|
| Eggs | Length | 1.83 ± 0.09 |
| Diameter | 1.27 ± 0.07 | |
| L1 | Length | 0.49 ± 0.17 |
| Diameter | 0.17 ± 0.10 | |
| L2 | Length | 0.86 ± 0.41 |
| Diameter | 0.27 ± 0.12 | |
| L3 | Length | 1.06 ± 0.41 |
| Diameter | 0.34 ± 0.14 | |
| L4 | Length | 1.70 ± 0.49 |
| Diameter | 0.48 ± 0.11 |
| Plant Species | 7 Days | 21 Days | 90 Days | New Adults |
|---|---|---|---|---|
| Ficus pandurata | 51.7 | 80.0 | 100 | 0.0 |
| Ficus carica | 0.0 | 20.0 | 47.2 | 6.0 |
| Ficus benjamina | 0.0 | 20.0 | 48.0 | 0.0 |
| Ficus microcarpa | 20.7 | 46.6 | 66.6 | 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farina, P.; Mazza, G.; Benvenuti, C.; Cutino, I.; Giannotti, P.; Conti, B.; Bedini, S.; Gargani, E. Biological Notes and Distribution in Southern Europe of Aclees taiwanensis Kȏno, 1933 (Coleoptera: Curculionidae): A New Pest of the Fig Tree. Insects 2021, 12, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010005
Farina P, Mazza G, Benvenuti C, Cutino I, Giannotti P, Conti B, Bedini S, Gargani E. Biological Notes and Distribution in Southern Europe of Aclees taiwanensis Kȏno, 1933 (Coleoptera: Curculionidae): A New Pest of the Fig Tree. Insects. 2021; 12(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010005
Chicago/Turabian StyleFarina, Priscilla, Giuseppe Mazza, Claudia Benvenuti, Ilaria Cutino, Paolo Giannotti, Barbara Conti, Stefano Bedini, and Elisabetta Gargani. 2021. "Biological Notes and Distribution in Southern Europe of Aclees taiwanensis Kȏno, 1933 (Coleoptera: Curculionidae): A New Pest of the Fig Tree" Insects 12, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010005