Contrasting Behavioral and Electrophysiological Responses of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) to Volatiles Emitted from the Tree of Heaven, Ailanthus altissima



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Attractiveness of Bare Phloem of Healthy and Injured Trees to Adult E. scrobiculatus and E. brandti
2.2. Attractiveness of Various Parts of A. altissima to Adult E. scrobiculatus and E. brandti
2.2.1. Large Still-Air Arena Experiment
2.2.2. Y-Tube Bioassay
2.3. Characterization of Volatiles from Different Parts of A. altissima
2.3.1. Collection of Plant Volatiles by Headspace-Solid Phase Microextraction (HS-SPME)
2.3.2. Collection of Plant Volatiles by Dynamic Headspace Method
2.4. Electroantennogram Responses
2.5. Data Analyses
3. Results
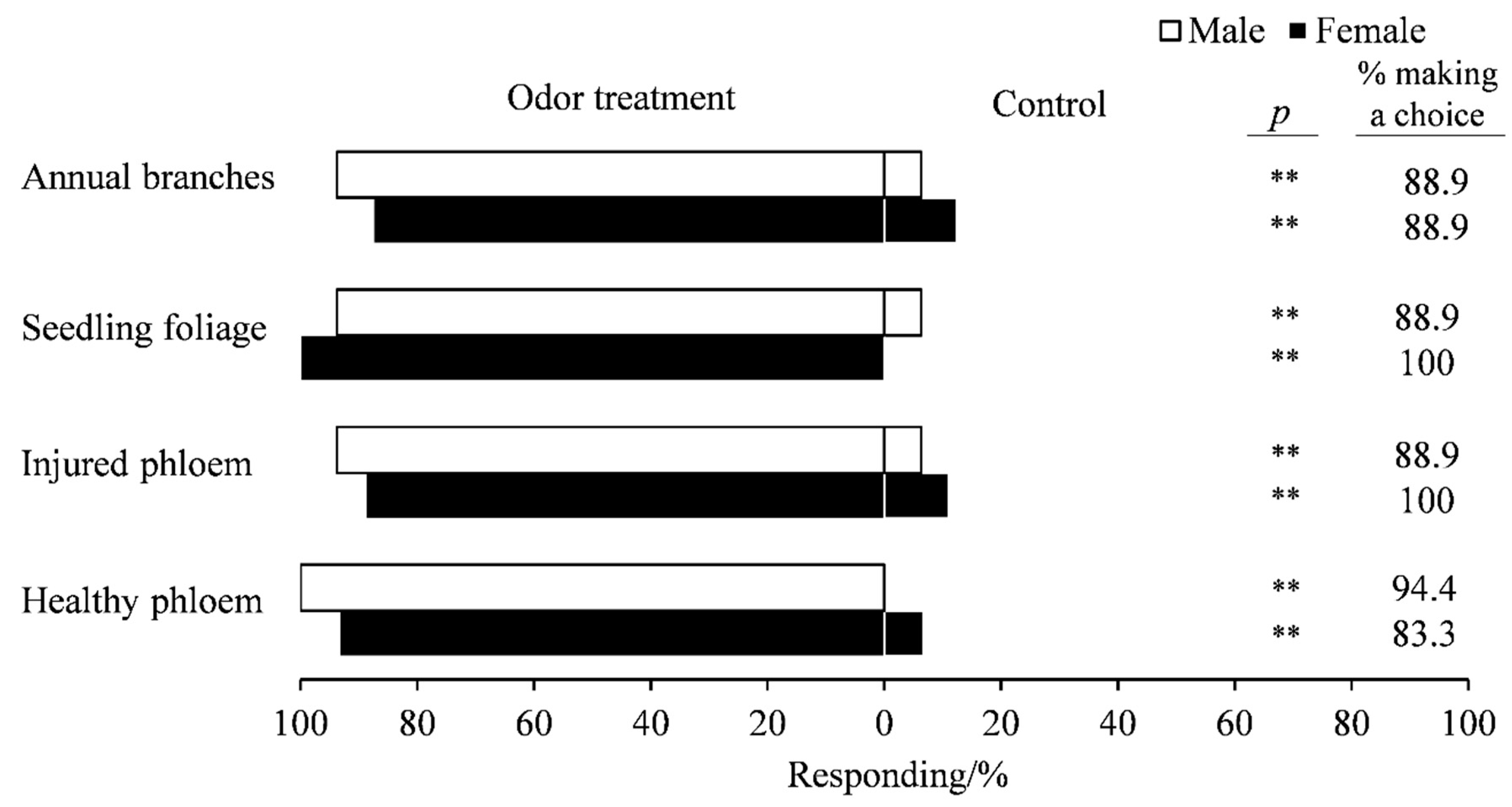
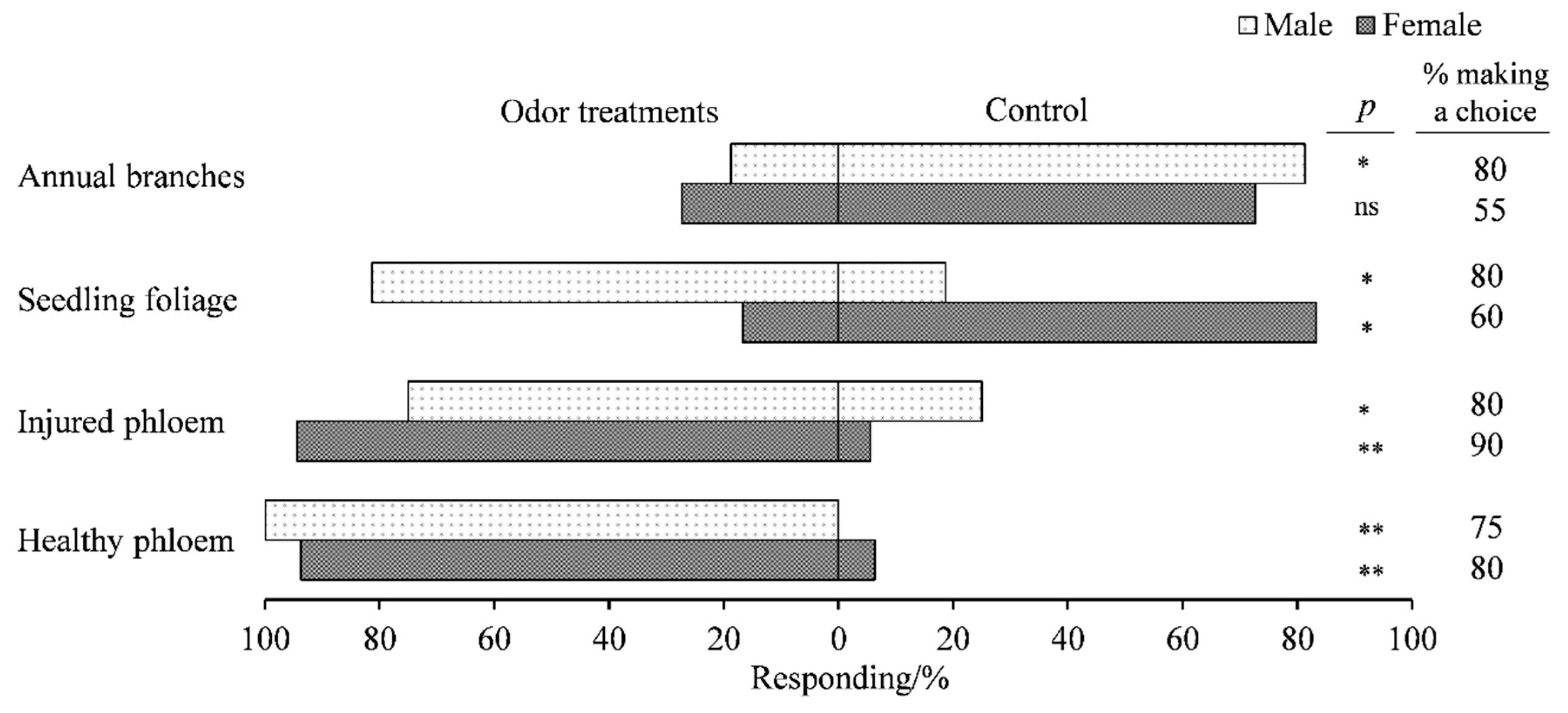
3.1. Attractiveness of Various Parts of A. altissima to Adult E. scrobiculatus and E. brandti
3.2. Characterization of Volatiles from Different Parts of A. altissima
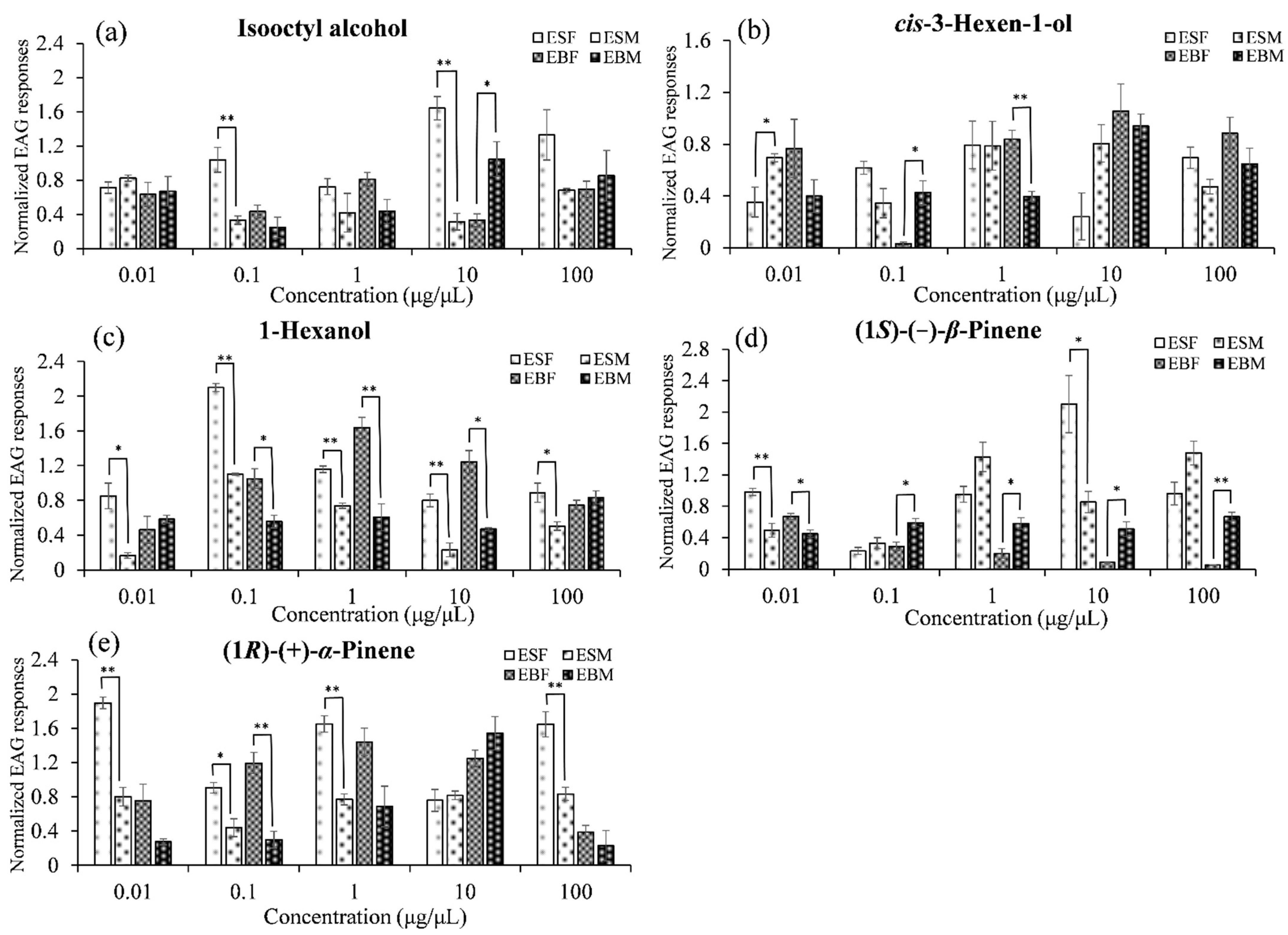
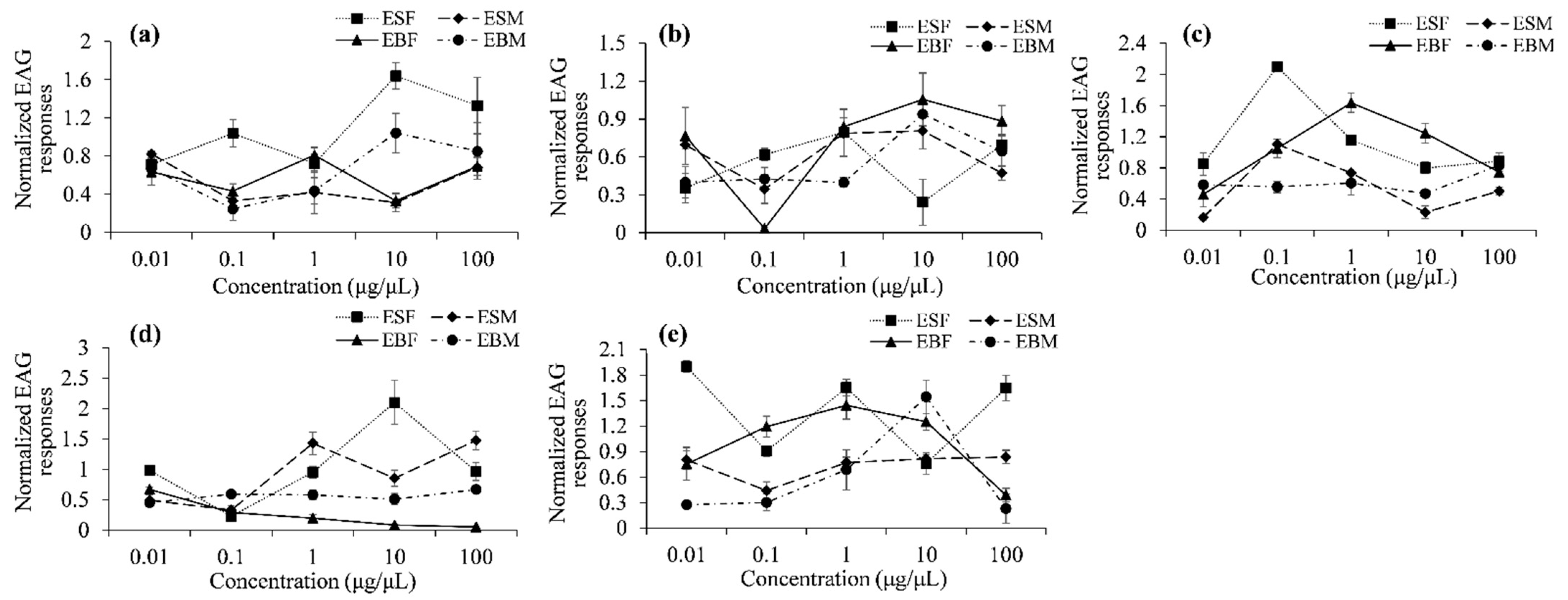
3.3. Electroantennogram Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Gause, G.F. The Struggle for Existence; The Williams & Wilkins Company: Baltimore, MD, USA, 1934; pp. 12–26. [Google Scholar]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Siemers, B.M.; Schnitzer, H.U. Echolocation signals reflect niche differentiation in five sympatric congeneric bat species. Nature 2004, 429, 657–661. [Google Scholar] [CrossRef]
- Jaenike, J. Host specialization in phytophagous insects. Annu. Rev. Ecol. Syst. 1990, 21, 243–273. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera of Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae); Entomopraxis: Barcelona, Spain, 1999; p. 315. [Google Scholar]
- Yang, G.J.; Yong, H.L.; Wang, X.P. The biological characters and behavior of Eucryptorrhynchus chinensis. Chin. J. Appl. Entomol. 2008, 45, 65–69. [Google Scholar]
- Herrick, N.J.; McAvoy, T.J.; Snyder, A.L.; Salom, S.M.; Kok, L.T. Host-range testing of Eucryptorrhynchus brandti (Coleoptera: Curculionidae), a candidate for biological control of tree-of-heaven, Ailanthus altissima. Environ. Entomol 2012, 41, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Qin, K.X. Damage of Eucryptorrhynchus brandti to tree of heaven and its control. Chin. J. Appl. Entomol. 1996, 33, 279. [Google Scholar]
- Yu, Q.Q.; Chen, C.; Liu, Z.K.; Sun, Y.W.; Cao, C.J.; Bao, S.; Wen, J.B. Occurrence and life-history of Eucryptorrhynchus chinensis in Lingwu, Ningxia. Chin. J. Appl. Entomol. 2012, 49, 1005–1009. [Google Scholar]
- McAvoy, T.J.; Salom, S.M.; Yu, B.; Ji, H.L.; Du, Y.Z.; Johnson, N.; Kok, L.T. Occurrence and development of Eucryptorrhynchus brandti and E. chinensis (Coleoptera: Curculionidae) on Ailanthus altissima trees subjected to different levels of mechanical damage. Biocontrol. Sci. Technol. 2014, 24, 65–79. [Google Scholar] [CrossRef]
- Ji, Y.C.; Gao, P.; Zhang, G.Y.; Wen, C.; Wen, J.B. Micro-habitat niche differentiation contributing to coexistence of Eucryptorrhynchus scrobiculatus Motschulsky and Eucryptorrhynchus brandti (Harold). Biocontrol. Sci. Technol. 2017, 27, 1180–1194. [Google Scholar] [CrossRef]
- Hu, D.W.; Qiu, L.F.; Wang, J.H. Occurrence and prevention of Eucryptorrhynchus brandti in Beijing. J. Gard. Sci. Technol. 2012, 3, 24–25. [Google Scholar]
- Yang, K.L.; Wen, X.J.; Ren, Y.; Wen, J.B. Novel trunk trap net designs for the control of Eucryptorrhynchus scrobiculatus (Coleoptera: Curculionidae). Pest Manag. Sci. 2019, 75, 2618–2626. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Wang, J.G.; Lang, X.R.; Sun, P.; Xing, L.R.; He, Z.Y. Preliminary study on integrated prevention and control technology of Eucryptorrhynchus chinensis and E. brandti in Ningxia, China. Chin. Agric. Technol. 2012, 32, 50–51. [Google Scholar]
- Yang, P. A Preliminary Study the Artificial Raising and Control of Eucryptorrhynchus Chinensis; Beijing Forestry University: Beijing, China, 2015. [Google Scholar]
- Yang, K.L.; Wen, X.J.; Zhang, G.Y.; Wen, J.B. Evaluation of trap designs and food attractants for trapping Eucryptorrhynchus scrobiculatus (Coleoptera: Curculionidae). Biocontrol. Sci. Technol. 2019, 29, 28–43. [Google Scholar] [CrossRef]
- Yang, K.L.; Wen, J.B. Efficacy of trunk trap nets and insecticides applied alone and in combination for control of tree-of-heaven root weevil Eucryptorrhynchus scrobiculatus in Ailanthus altissima plantations. Forests 2019, 10, 972. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.L.; Wen, X.J.; Guo, W.J.; Wen, J.B. A novel adhesive trunk trap net for trapping Eucryptorrhynchus brandti (Coleoptera: Curculionidae). Pest Manag. Sci. 2019, 75, 3218–3225. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.C.; Dorn, S. Synergism between aromatic compounds and green leaf volatiles derived from the host plant underlies female attraction in the oriental fruit moth. Entomol. Exp. Appl. 2010, 125, 185–194. [Google Scholar] [CrossRef]
- Ruther, J. Retention index database for identification of general green leaf volatiles in plants by coupled capillary gas chromatography-mass spectrometry. J. Chromatogr. A 2000, 890, 313–319. [Google Scholar] [CrossRef]
- Ren, L.L. Electrophysiological and Behavioral Responses of Monochamus Alternatus and Parasitoid Dastarcus Helophoroides to Semiochemicals of Several Tree Species; Beijing Forestry University: Beijing, China, 2014. [Google Scholar]
- Paré, P.W.; Tumlinson, J.H. Plant volatiles as a defense against insect herbivores. Plant Physiol. 1999, 121, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Mastelić, J.; Jerković, I. Volatile constituents from the leaves of young and old Alianthus altissima (Mill.) Swingle tree. Croat Chem. Acta 2002, 75, 189–197. [Google Scholar]
- Li, D.P.; Cao, Y.X.; Chen, N.Z.; Ma, F.; Yu, Y.X.; Li, Z.H.; Chen, H.J. Studies on volatiles compounds from leaf of Alianthus altissima extracts by simultaneous distillation extractor and solid phase micro-extraction. Plant Quar. 2013, 27, 1–5. [Google Scholar]
- Xie, X.Y. Research on Composition and Release Regularities of VOCs from Main Landscape Plants in Xi’an; Northwest A & F University: Yulin, China, 2016. [Google Scholar]
- Ji, X.Y.; Yan, J.; Wang, J. Identification and comparison of volatile components from Toona Sinensis and Alianthus altissima leaves. J. Anhui Agric. Sci. 2018, 46, 179–181. [Google Scholar]
- Wen, X.J.; Wang, Q.; Gao, P.; Wen, J.B. Identification and comparison of chemosensory genes in the antennal transcriptomes of Eucryptorrhynchus scrobiculatus and E. brandti fed on Ailanthus altissima. Front. Physiol. 2018, 9, 1652. [Google Scholar] [CrossRef]
- Birkett, M.A.; Bruce, T.J.A.; Martin, J.L.; Smart, L.E.; Oakley, J.; Wadhams, L.J. Responses of female orange wheat blossom midge, Sitodiplosis mosellana, to wheat panicle volatiles. J. Chem. Ecol. 2004, 30, 1319–1328. [Google Scholar] [CrossRef]
- Riffell, J.A.; Lei, H.; Christensen, T.A.; Hildebrand, J.G. Characterization and coding of behaviorally significant odor mixtures. Curr. Biol. 2009, 19, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, T.J.A.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects--finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Dimock, M.B.; Tingey, W.M. Host acceptance behaviour of Colorado potato beetle larvae influenced by potato glandular trichomes. Physiol. Entomol. 1988, 13, 399–406. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Wood, D.L. Feeding and boring behavior of the bark beetle Ips paraconfusus (Coleoptera: Scolytidae) on the bark of a host and non-host tree species. Can. Entomol. 1980, 112, 797–809. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | CAS | Area under Peak | |
|---|---|---|---|---|
| Healthy Phloem | Injured Phloem | |||
| 1 | (1R)-(+)-α-Pinene | 7785-70-8 | 599,387 | 302,479 |
| 2 | Camphene | 79-92-5 | 289,369 | |
| 3 | β-Phellandrene | 555-10-2 | 78,859 | |
| 4 | (1S)-(−)-β-Pinene | 18172-67-3 | 358,576 | |
| 5 | Myrcene | 123-35-3 | 1,360,591 | |
| 6 | (+)-(4R)-Limonene | 5989-27-5 | 1,898,791 | |
| 7 | β-Ocimene | 3338-55-4 | 4,851,574 | |
| 8 | (+)-2-Carene | 554-61-0 | 1,210,188 | |
| 9 | α-Cubebene | 17699-14-8 | 12,832,275 | |
| 10 | Copaene | 3856-25-5 | 10,567,644 | |
| 11 | β-Caryophyllene | 87-44-5 | 65,276 | 42,295,799 |
| 12 | α-Humulene | 6753-98-6 | 7,472,179 | |
| 13 | β-Copaene | 2,172,542 | ||
| 14 | (+)-δ-Cadinene | 483-76-1 | 1,005,043 | |
| 15 | l-Calamenene | 483-77-2 | 288,668 | |
| 16 | Caryophyllene oxide | 1139-30-6 | 616,535 | |
| 17 | 2-Xylene | 95-47-6 | 184,083 | 1,196,424 |
| 18 | p-Xylene | 106-42-3 | 332,852 | |
| 19 | 1-Ethyl-4-methylbenzene | 622-96-8 | 68,356 | |
| 20 | 3-Ethyltoluene | 620-14-4 | 972,029 | |
| 21 | 2-Ethyltoluene | 611-14-3 | 560,994 | |
| 22 | Mesitylen | 108-67-8 | 75,378 | 5,228,481 |
| 23 | 1,2,3-Trimethylbenzene | 526-73-8 | 69,816 | 566,908 |
| 24 | 1-Methyl-3-propylbenzene | 1074-43-7 | 445,924 | |
| 25 | 2-Ethyl-p-xylene | 1758-88-9 | 981,883 | |
| 26 | 4-Ethyl-m-xylene | 874-41-9 | 785,844 | |
| 27 | 1,2,3,4-Tetramethylbenzene | 488-23-3 | 520,840 | |
| 28 | 1,2,3,5-Tetramethylbenzene | 527-53-7 | 864,315 | |
| 29 | 1,2,4,5-Tetramethylbenzene | 95-93-2 | 303,672 | |
| 30 | 3-Methylcyclopentanol | 18729-48-1 | 159,700 | |
| 31 | cis-3-Hexen-1-ol | 928-96-1 | 863,559 | |
| 32 | Cyclohexanol | 108-93-0 | 333,474 | |
| 33 | 1-Hexanol | 111-27-3 | 4,902,291 | 7,042,158 |
| 34 | 2-Ethyl-1-hexanol | 104-76-7 | 515,699 | |
| 35 | 3,3-Dimethyl-1,2-epoxybutane | 2245-30-9 | 1,169,014 | |
| 36 | 2,4-Dimethylhexane | 589-43-5 | 110,544 | |
| 37 | Tetradecane | 629-59-4 | 57,964 | |
| 38 | d-Camphor | 464-49-3 | 59,699 | |
| 39 | 2-Hendecanone | 112-12-9 | 331,169 | |
| 40 | Tetradecanal | 124-25-4 | 644,070 | |
| 41 | Pentadecanal | 2765-11-9 | 286,561 | |
| 42 | Dibutyl ether | 142-96-1 | 1,178,162 | 1,392,083 |
| 43 | 1,2-Dimethoxybenzene | 91-16-7 | 540,650 | |
| 44 | Dibutyl phthalate | 84-74-2 | 272,435 | 554,530 |
| 45 | Oxetane, 3-(1-methylethyl)- | 10317-17-6 | 85,020 | |
| 46 | Di-tert-butyl peroxide | 110-05-4 | 119,142 | |
| 47 | 4-Hydroxy-3-methylbutanal | 56805-34-6 | 972,544 | |
| 48 | Aciphyllene | 87745-31-1 | 330,730 | |
| 49 | Cubebene | 13744-15-5 | 2,455,168 | |
| 50 | (+)-Epi-bicyclosesquiphellandrene | 54274-73-6 | 367,917 | |
| Number | Name | CAS | Relative Content (in %) | |||
|---|---|---|---|---|---|---|
| Healthy Phloem | Injured Phloem | Seedling Foliage | Annual Branches | |||
| 1 | (1R)-(+)-α-Pinene | 7785-70-8 | 2.92 | 0.53 | ||
| 2 | (−)-Camphene | 5794-04-7 | 8.22 | |||
| 3 | Camphene | 79-92-5 | 0.97 | |||
| 4 | β-Pinene | 127-91-3 | 5.49 | |||
| 5 | (1S)-(−)-β-Pinene | 18172-67-3 | 0.54 | |||
| 6 | 1-Decene | 872-05-9 | 1.7 | |||
| 7 | (+)-(4R)-Limonene | 5989-27-5 | 1.21 | |||
| 8 | 1-Undecene | 821-95-4 | 2.01 | |||
| 9 | 1-Tridecene | 2437-56-1 | 10.05 | |||
| 10 | α-Pinene | 3856-25-5 | 1.44 | 0.93 | 0.74 | |
| 11 | β-Bourbonene | 5208-59-3 | 4.33 | |||
| 12 | 1-Tetradecene | 1120-36-1 | 17.58 | |||
| 13 | β-Elemene | 515-13-9 | 8.24 | |||
| 14 | β-Caryophyllene | 87-44-5 | 33.88 | 26.94 | 21.63 | |
| 15 | 1-Pentadecene | 13360-61-7 | 2.54 | |||
| 16 | β-Copaene | 8.94 | 41.72 | 41.43 | ||
| 17 | α-Farnesene | 502-61-4 | 12.74 | 22.29 | ||
| 18 | 1-Heptadecene | 6765-39-5 | 2.91 | |||
| 19 | 2,4-Dimethylheptane | 2213-23-2 | 1.62 | |||
| 20 | n-Hendecane | 1120-21-4 | 6.91 | |||
| 21 | Dodecane | 112-40-3 | 1.2 | |||
| 22 | n-Tridecane | 629-50-5 | 2.88 | 5.45 | ||
| 23 | Tetradecane | 629-59-4 | 3.34 | 2.67 | ||
| 24 | n-Pentadecane | 629-62-9 | 3.13 | 6.01 | ||
| 25 | n-Heptadecane | 629-78-7 | 1.12 | |||
| 26 | 1-Dodecanol | 112-53-8 | 11 | |||
| 27 | 1-Tridecanol | 112-70-9 | 6.81 | |||
| 28 | 1-Pentadecanol | 629-76-5 | 1.21 | |||
| 29 | Ethyl 2-methylbutyrate | 7452-79-1 | 2.24 | |||
| 30 | Ethyl tiglate | 5837-78-5 | 0.67 | |||
| 31 | Leaf acetate | 3681-71-8 | 11.7 | 1.65 | ||
| 32 | Bis(2-ethylhexyl) adipate | 103-23-1 | 0.63 | |||
| 33 | Carvacrol | 499-75-2 | 3.39 | |||
| 34 | 2,4-Di-tert-butylphenol | 96-76-4 | 2.39 | |||
| 35 | 2-Phenylethyl-1,1,2,2-d4-amine | 876-20-0 | 17.52 | |||
| 36 | Cuminaldehyde | 122-03-2 | 6.1 | |||
| 37 | 4-Ethylbenzaldehyde | 4748-78-1 | 2.15 | |||
| 38 | Phenylethylene | 100-42-5 | 6.12 | |||
| 39 | Isobutylbenzene | 1.04 | ||||
| 40 | Hexane,2,2,3,3-tetramethyl- | 13475-81-5 | 1.59 | |||
| 41 | 2-Pentylfuran | 3777-69-3 | 2.82 | |||
| 42 | 4-Ethylcumen | 4218-48-8 | 2.11 | |||
| 43 | 5-Ethylundecane | 17453-94-0 | 2.63 | |||
| Compound | Insect | Mean EAG Responses (± SE) | ||||
|---|---|---|---|---|---|---|
| 0.01 μg/μL | 0.1 μg/μL | 1 μg/μL | 10 μg/μL | 100 μg/μL | ||
| Isooctyl alcohol | ESF | 0.71 ± 0.07c | 1.04 ± 0.14bc | 0.72 ± 0.1c | 1.64 ± 0.14a | 1.33 ± 0.29ab |
| ESM | 0.82 ± 0.04a | 0.33 ± 0.05b | 0.42 ± 0.22b | 0.31 ± 0.1b | 0.68 ± 0.02ab | |
| EBF | 0.63 ± 0.14abc | 0.43 ± 0.08bc | 0.81 ± 0.08a | 0.33 ± 0.08c | 0.69 ± 0.09ab | |
| EBM | 0.67 ± 0.17ab | 0.25 ± 0.12b | 0.43 ± 0.14ab | 1.04 ± 0.21a | 0.85 ± 0.3ab | |
| cis-3-Hexen-1-ol | ESF | 0.35 ± 0.12ab | 0.62 ± 0.05ab | 0.79 ± 0.18a | 0.24 ± 0.18b | 0.7 ± 0.08a |
| ESM | 0.7 ± 0.03ab | 0.34 ± 0.11b | 0.79 ± 0.19a | 0.81 ± 0.14a | 0.47 ± 0.06ab | |
| EBF | 0.77 ± 0.22a | 0.03 ± 0.01b | 0.84 ± 0.07a | 1.06 ± 0.21a | 0.88 ± 0.12a | |
| EBM | 0.4 ± 0.13b | 0.43 ± 0.09b | 0.4 ± 0.04b | 0.94 ± 0.09a | 0.65 ± 0.12ab | |
| 1-Hexanol | ESF | 0.85 ± 0.15c | 2.1 ± 0.05a | 1.16 ± 0.04b | 0.8 ± 0.07c | 0.89 ± 0.11bc |
| ESM | 0.16 ± 0.03d | 1.1 ± 0.01a | 0.74 ± 0.03b | 0.23 ± 0.08d | 0.5 ± 0.05c | |
| EBF | 0.46 ± 0.16d | 1.05 ± 0.11bc | 1.64 ± 0.12a | 1.25 ± 0.13b | 0.75 ± 0.05cd | |
| EBM | 0.58 ± 0.04ab | 0.56 ± 0.07ab | 0.61 ± 0.16ab | 0.47 ± 0.01b | 0.83 ± 0.08a | |
| (1S)-(−)-β-Pinene | ESF | 0.98 ± 0.05b | 0.23 ± 0.05c | 0.95 ± 0.1b | 2.1 ± 0.36a | 0.96 ± 0.15b |
| ESM | 0.49 ± 0.09bc | 0.33 ± 0.07c | 1.43 ± 0.19a | 0.85 ± 0.14b | 1.48 ± 0.15a | |
| EBF | 0.67 ± 0.04a | 0.29 ± 0.05b | 0.2 ± 0.06bc | 0.08 ± 0.0048cd | 0.05 ± 0.0043d | |
| EBM | 0.45 ± 0.05a | 0.59 ± 0.05a | 0.58 ± 0.07a | 0.51 ± 0.09a | 0.67 ± 0.05a | |
| (1R)-(+)-α-Pinene | ESF | 1.9 ± 0.07a | 0.91 ± 0.06b | 1.65 ± 0.1a | 0.76 ± 0.13b | 1.65 ± 0.15a |
| ESM | 0.8 ± 0.11a | 0.44 ± 0.1b | 0.77 ± 0.06b | 0.82 ± 0.05b | 0.83 ± 0.08b | |
| EBF | 0.75 ± 0.19b | 1.19 ± 0.12a | 1.44 ± 0.16a | 1.25 ± 0.09a | 0.39 ± 0.08b | |
| EBM | 0.28 ± 0.03b | 0.3 ± 0.1b | 0.69 ± 0.24b | 1.54 ± 0.19a | 0.23 ± 0.18b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, X.; Yang, K.; Piñero, J.C.; Wen, J. Contrasting Behavioral and Electrophysiological Responses of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) to Volatiles Emitted from the Tree of Heaven, Ailanthus altissima. Insects 2021, 12, 68. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010068
Wen X, Yang K, Piñero JC, Wen J. Contrasting Behavioral and Electrophysiological Responses of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) to Volatiles Emitted from the Tree of Heaven, Ailanthus altissima. Insects. 2021; 12(1):68. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010068
Chicago/Turabian StyleWen, Xiaojian, Kailang Yang, Jaime C. Piñero, and Junbao Wen. 2021. "Contrasting Behavioral and Electrophysiological Responses of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) to Volatiles Emitted from the Tree of Heaven, Ailanthus altissima" Insects 12, no. 1: 68. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010068