Identification of Wild-Type CYP321A2 and Comparison of Allelochemical-Induced Expression Profiles of CYP321A2 with Its Paralog CYP321A1 in Helicoverpa zea

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Plant Xenobiotics Induction
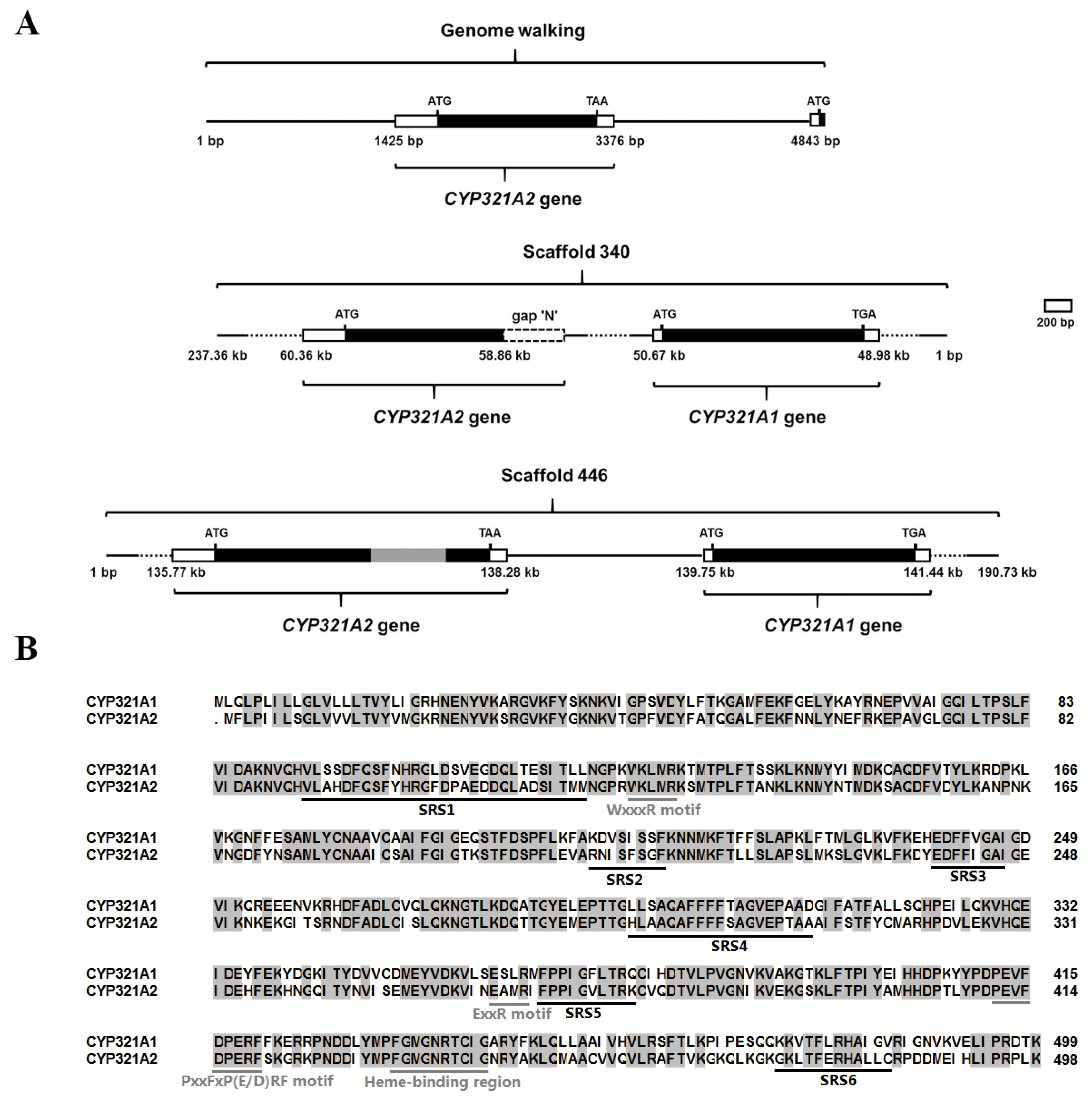
2.2. DNA Extraction and Cloning of Genomic Sequences
2.3. RNA Extraction and First Strand cDNA Synthesis
2.4. Identification of CYP321A2 Transcript by 5′- and 3′- RACE
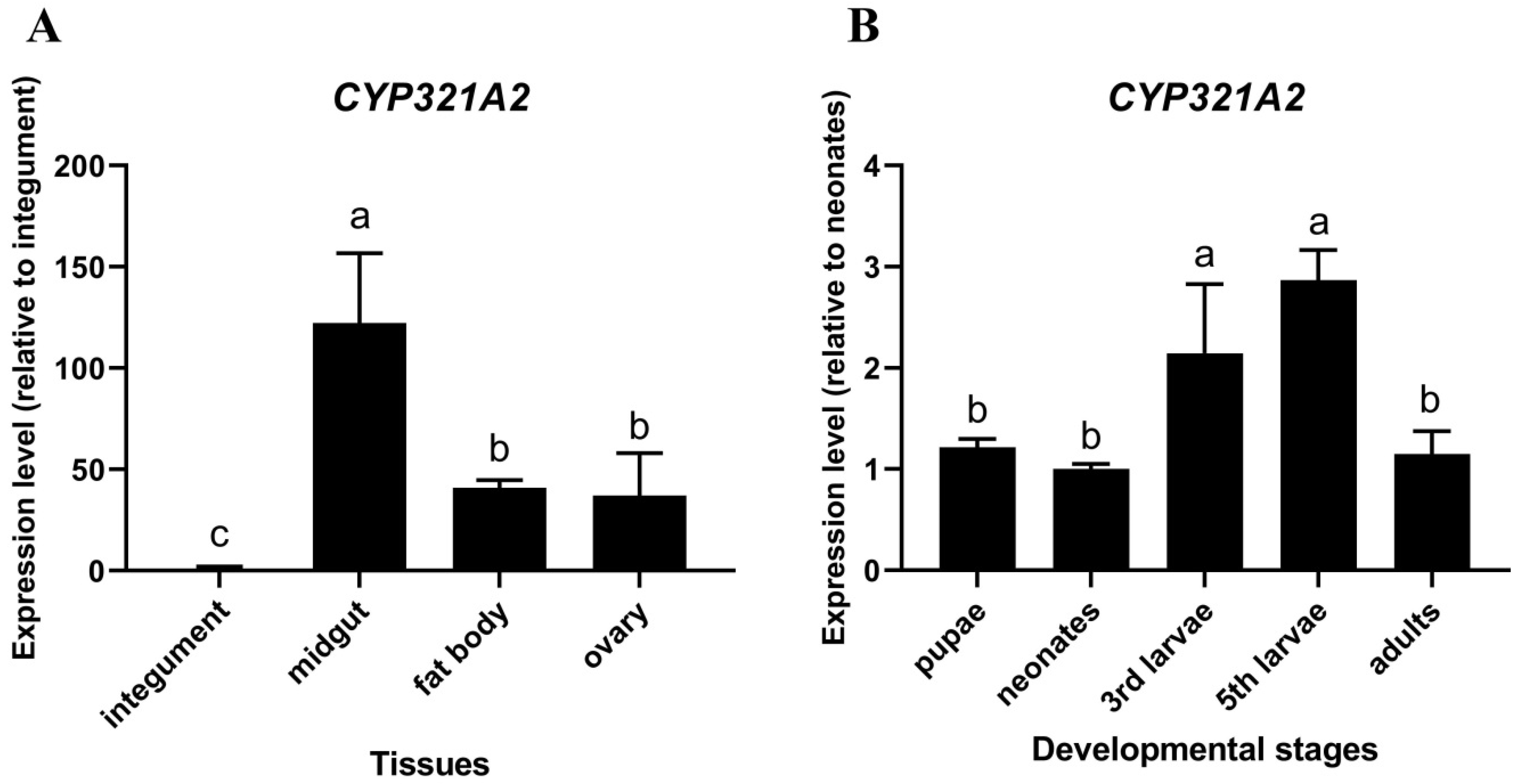
2.5. Analysis of CYP321A2 Expression Level in Different Tissues and Developmental Stages
2.6. Allelochemical and Plant Signal Molecule Induced Expression of CYP321A2 and CYP321A1 in Midgut and Fat Body
2.7. Sequence Analysis
2.8. Statistical Analysis
3. Results
3.1. Identification of the Wild-Type Intact CYP321A2
3.2. Tissue-Specific and Developmental Expressions of CYP321A2
3.3. Allelochemical and Plant Signal Xenobiotic Induced Expressions of CYP321A2 and CYP321A1
3.4. Sequence Comparison in the 5′ Flanking Regions of CYP321A2 and CYP321A1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berenbaum, M.R. Evolution of specialization in insect-umbellifer associations. Annu. Rev. Entomol. 1990, 35, 319–343. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. Host-Plant. Selection by Phytophagous Insects; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Baudry, J.; Berenbaum, M.R.; Schuler, M.A. Structural and functional divergence of insect CYP6B proteins: From specialist to generalist cytochrome P450. Proc. Natl. Acad. Sci. USA 2004, 101, 2939–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.B.; Schuler, M.A.; Berenbaum, M.R. A host-inducible cytochrome P450 from a host-specific caterpillar: Molecular cloning and evolution. Proc. Natl. Acad. Sci. USA 1992, 89, 10920–10924. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, R. Insect CYP Genes and P450 Enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Elsevier: San Diego, CA, USA, 2012; pp. 236–316. [Google Scholar]
- Schuler, M.A. The role of cytochrome P450 monooxygenases in plant-insect interactions. Plant Physiol. 1996, 112, 1411–1419. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, M.R. Postgenomic chemical ecology: From genetic code to ecological interactions. J. Chem. Ecol. 2002, 28, 873–896. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Plant allelochemicals differentially regulate Helicoverpa zea cytochrome P450 genes. Insect Mol. Biol. 2002, 11, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.F.; Carter, A.J.; Chiu, C.H. Gene conversion may aid adaptive peak shifts. J. Theor. Biol. 2000, 207, 495–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teshima, K.M.; Innan, H. The effect of gene conversion on the divergence between duplicated genes. Genetics 2004, 166, 1553–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osada, N.; Innan, H. Duplication and gene conversion in the Drosophila melanogaster genome. PLoS Genet. 2008, 4, e1000305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, T. Gene conversion and evolution of gene families: An overview. Genes 2010, 1, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, J.B. Sequence-dependent gene conversion: Can duplicated genes diverge fast enough to escape conversion? Genetics 1987, 117, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Berenbaum, M.R.; Schuler, M.A. Cytochrome P450 and actin genes expressed in Helicoverpa zea and Helicoverpa armigera: Paralogy/orthology identification, gene conversion, and evolution. Insect Biochem. Mol. Biol. 2002, 32, 311–320. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Jasmonate and salicylate induce expression of herbivore cytochrome P450 genes. Nature 2002, 419, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef]
- Tijet, N.; Helvig, C.; Feyereisen, R. The cytochrome P450 gene superfamily in Drosophila melanogaster: Annotation, intron–exon organization and phylogeny. Gene 2001, 262, 189–198. [Google Scholar] [CrossRef]
- Sasabe, M.; Wen, Z.; Berenbaum, M.R.; Schuler, M.A. Molecular analysis of CYP321A1, a novel cytochrome P450 involved in metabolism of plant allelochemicals (furanocoumarins) and insecticides (cypermethrin) in Helicoverpa zea. Gene 2004, 338, 163–175. [Google Scholar] [CrossRef]
- Zeng, R.; Wen, Z.; Niu, G.; Schuler, M.A.; Berenbaum, M.R. Allelochemical induction of cytochrome P450 monooxygenases and amelioration of xenobiotic toxicity in Helicoverpa zea. J. Chem. Ecol. 2007, 33, 449–461. [Google Scholar] [CrossRef]
- Zeng, R.; Wen, Z.; Niu, G.; Schuler, M.A.; Berenbaum, M.R. Enhanced toxicity and induction of cytochrome P450s suggest a cost of “eavesdropping” in a multitrophic interaction. J. Chem. Ecol. 2009, 35, 526–532. [Google Scholar] [CrossRef]
- Wen, Z.; Zeng, R.; Niu, G.; Berenbaum, M.R.; Schuler, M.A. Ecological significance of induction of broad-substrate cytochrome P450s by natural and synthetic inducers in Helicoverpa zea. J. Chem. Ecol. 2009, 35, 183–189. [Google Scholar] [CrossRef]
- Rupasinghe, S.G.; Wen, Z.; Chiu, T.; Schuler, M.A. Helicoverpa zea CYP6B8 and CYP321A1: Different molecular solutions to the problem of metabolizing plant toxins and insecticides. Protein Eng. Des. Sel. 2007, 20, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Wen, Z.; Rupasinghe, S.G.; Zeng, R.; Berenbaum, M.R.; Schuler, M.A. Aflatoxin B1 detoxification by CYP321A1 in Helicoverpa zea. Arch. Insect Biochem. 2008, 69, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P.; Cohen, R.W.; Friedman, S. An improved procedure for laboratory rearing of the corn earworm, Heliothis zea (Lepidoptera: Noctuidae). Great Lakes Entomol. 1984, 17, 113–118. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; pp. 7.23–7.26. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hasemann, C.A.; Kurumbail, R.G.; Boddupalli, S.S.; Peterson, J.A.; Deisenhofer, J. Structure and function of cytochromes P450: A comparative analysis of three crystal structures. Structure 1995, 3, 41–62. [Google Scholar] [CrossRef]
- Werck-Reichhart, D.; Feyereisen, R. Cytochromes P450: A success story. Genome Biol. 2000, 1, 3003.1–3003.9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Luo, X.; Ni, X.; Zhang, Y.; Li, X. Functional characterization of cis-acting elements mediating flavone-inducible expression of CYP321A1. Insect Biochem. Mol. Biol. 2010, 40, 898–908. [Google Scholar] [CrossRef]
- Zhang, C.; Wong, A.; Zhang, Y.; Ni, X.; Li, X. Common and unique cis-acting elements mediate xanthotoxin and flavone induction of the generalist P450 CYP321A1. Sci. Rep. 2014, 4, 6490. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Berenbaum, M.R.; Schuler, M.A. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Hodgson, E. Microsomal Monooxygenases. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon: Oxford, UK, 1985; pp. 225–331. [Google Scholar]
- Lynch, M. The Origins of Genome Architecture; Sinauer Associates: Sunderland, MA, USA, 2007. [Google Scholar]
- Ohno, S. Evolution by Gene Duplication; Springer-Verlag: Berlin/Heidelberg, Germany, 1970. [Google Scholar]
- Wang, H.; Shi, Y.; Wang, L.; Liu, S.; Wu, S.; Yang, Y.; Feyereisen, R.; Wu, Y. CYP6AE gene cluster knockout in Helicoverpa armigera reveals role in detoxification of phytochemicals and insecticides. Nat. Commun. 2018, 9, 4820. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [CrossRef]
- Seigler, D.S. Plant Secondary Metabolism; Kluwer Academic: Boston, MA, USA, 1998; pp. 306–309. [Google Scholar]
- Kogan, J.D.; Sell, K.; Stinner, R.E.; Bradley, J.R.; Kogan, M. A Bibliography of Heliothis zea (Boddie) and H. virescens (F.) (Lepidoptera: Noctuidae); INTSOY Series Number 17; International Agricultural Publication: Urbana, IL, USA, 1978. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Plant-Derived Compounds | CYP321A2 | CYP321A1 | |||
|---|---|---|---|---|---|
| Midgut | Fat Body | Midgut | Fat Body | ||
| plant allelochemicals | Xanthotoxin | 60.42 ± 14.70 | 20.62 ± 4.37 | 4.55 ± 0.74 | 7.94 ± 2.42 |
| chlorogenic acid | 12.86 ± 8.05 | 10.58 ± 2.21 | 7.05 ± 0.87 | 4.60 ± 0.73 | |
| indole-3-carbinol | 7.13 ± 1.64 | 2.60 ± 0.30 | 33.74 ± 4.01 | 16.59 ± 3.17 | |
| Flavone | 850.76 ± 389.11 | 16.99 ± 3.18 | 306.82 ± 15.38 | 10.36 ± 1.92 | |
| Rutin | 12.58 ± 2.62 | 1.06 ± 0.37 | 24.55 ± 1.21 | 3.54 ± 0.90 | |
| Gossypol | 1.19 ± 0.66 | 1.23 ± 0.25 | 1.68 ± 0.47 | 1.37 ± 0.07 | |
| 2-tridecanone | 18.25 ± 2.64 | 0.91 ± 0.40 | 20.34 ± 1.40 | 2.24 ± 0.28 | |
| Quercetin | 11.88 ± 3.69 | 2.13 ± 1.44 | 40.44 ± 2.66 | 4.25 ± 0.25 | |
| Coumarin | 61.18 ± 26.38 | 12.37 ± 2.16 | 102.36 ± 4.46 | 6.04 ± 0.99 | |
| plant signaling molecules | Jasmonate | 3.60 ± 1.34 | 0.45 ± 0.18 | 0.81 ± 0.13 | 2.65 ± 0.08 |
| Salicylate | 2.06 ± 0.20 | 0.61 ± 0.40 | 1.80 ± 0.42 | 1.55 ± 0.11 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Chen, S.; Xie, X.; Dong, S.; Li, X. Identification of Wild-Type CYP321A2 and Comparison of Allelochemical-Induced Expression Profiles of CYP321A2 with Its Paralog CYP321A1 in Helicoverpa zea. Insects 2021, 12, 75. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010075
Li S, Chen S, Xie X, Dong S, Li X. Identification of Wild-Type CYP321A2 and Comparison of Allelochemical-Induced Expression Profiles of CYP321A2 with Its Paralog CYP321A1 in Helicoverpa zea. Insects. 2021; 12(1):75. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010075
Chicago/Turabian StyleLi, Shengyun, Song Chen, Xingcheng Xie, Shuanglin Dong, and Xianchun Li. 2021. "Identification of Wild-Type CYP321A2 and Comparison of Allelochemical-Induced Expression Profiles of CYP321A2 with Its Paralog CYP321A1 in Helicoverpa zea" Insects 12, no. 1: 75. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010075