
Condition-Specific Competitive Effects of the Invasive Mosquito Aedes albopictus on the Resident Culex pipiens among Different Urban Container Habitats May Explain Their Coexistence in the Field



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Surveys
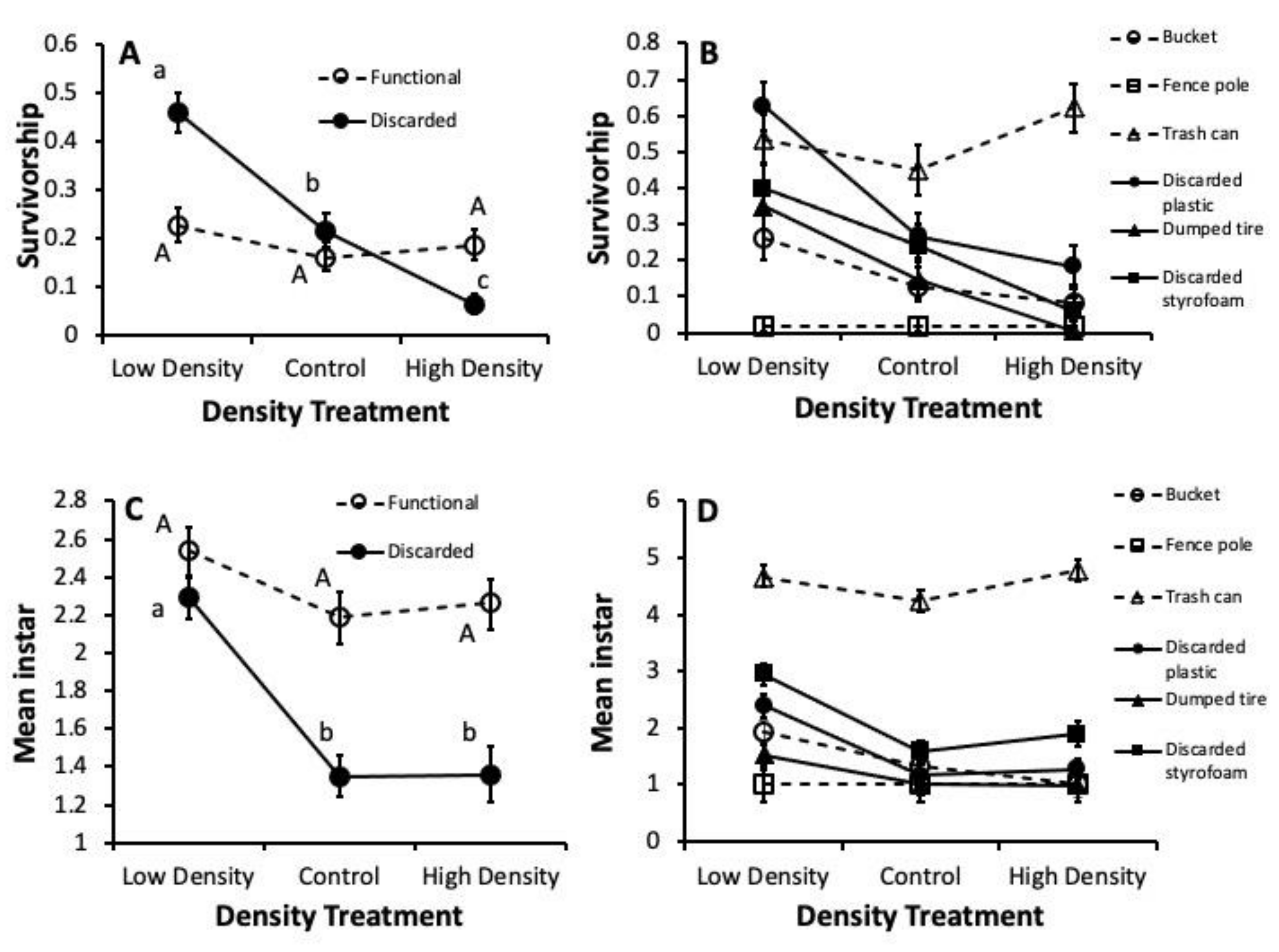
2.2. Competition Trial
2.3. Statistical Analyses
3. Results
3.1. Field Surveys
3.2. Competition Trial
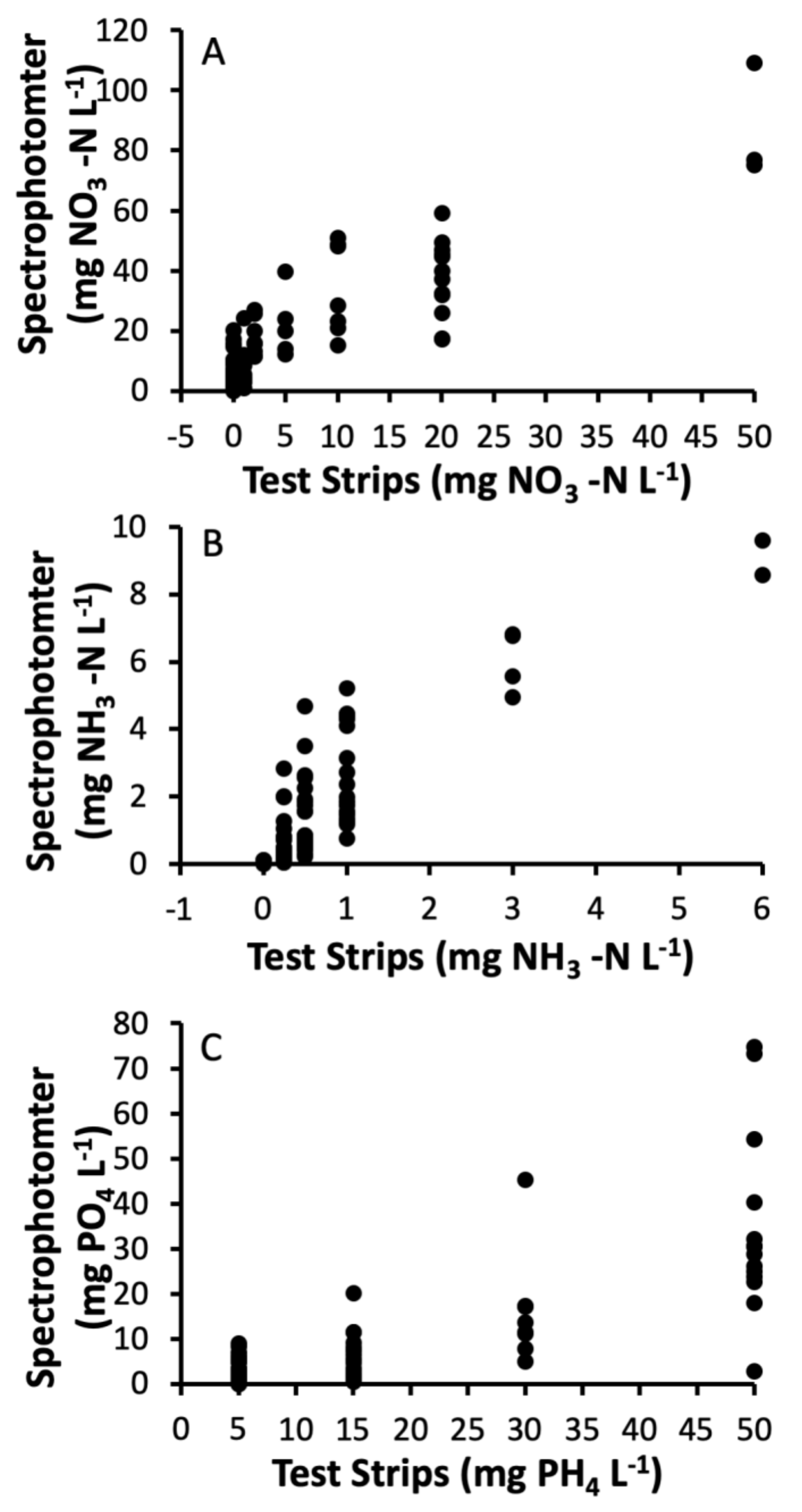
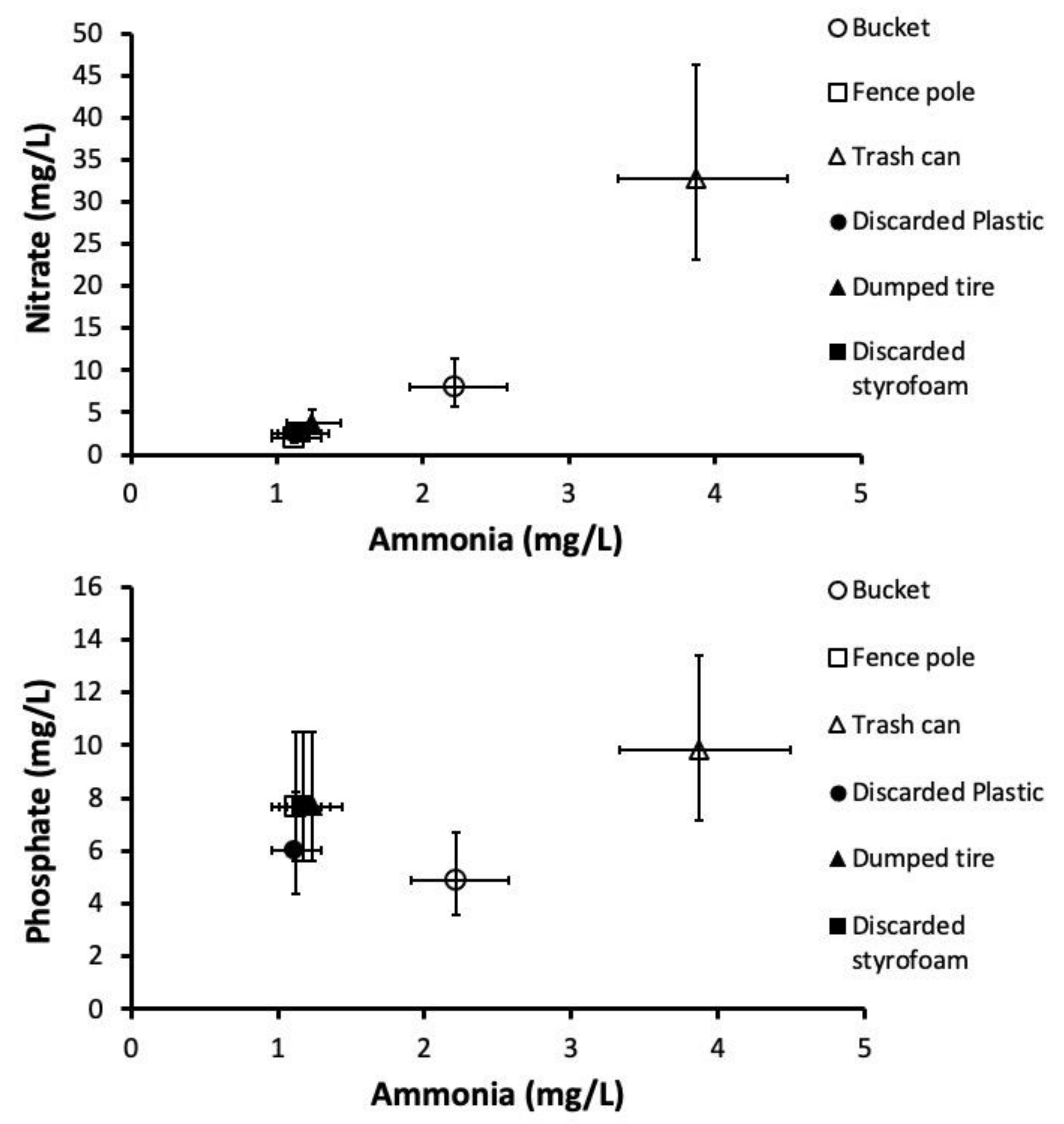
3.3. Nutrient Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D. Resource Competition and Community Structure; Princeton University Press Princeton: Princeton, NJ, USA, 1982. [Google Scholar]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contempory Approaches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Shorrocks, B.; Atkinson, W.; Charlesworth, P. Competition on a divided and ephemeral resource. J. Anim. Ecol. 1979, 48, 899–908. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Ann. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Chesson, P. Environmental variation and the coexistence of species. In Community Ecol; Diamond, J., Case, T.J., Eds.; Harper and Row: New York, NY, USA, 1986; pp. 240–256. [Google Scholar]
- Taniguchi, Y.; Nakano, S. Condition-dependent competition: Implications for the distributions of stream fishes. Ecology 2000, 81, 2027–2039. [Google Scholar] [CrossRef]
- Murrell, E.G.; Juliano, S.A. Detritus type alters the outcome of interspecific competition between Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Hui, C.; Richardson, D.M. Invasion Dynamics; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Chase, J.M.; Abrams, P.A.; Grover, J.P.; Diehl, S.; Chesson, P.; Holt, R.D.; Richards, S.A.; Nisbet, R.M.; Case, T.J. The interaction between predation and competition: A review and synthesis. Ecol. Lett. 2002, 5, 302–315. [Google Scholar]
- Blaustein, L.; Chase, J.M. Interactions between mosquito larvae and species that share the same trophic level. Ann. Rev. Entomol. 2007, 52, 489–507. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A. Species interactions among larval mosquitoes: Context dependence across habitat gradients. Ann. Rev. Entomol. 2009, 54, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Marini, G.; Guzzetta, G.; Baldacchino, F.; Arnoldi, D.; Montarsi, F.; Capelli, G. The effect of interspecific competition on the temporal dynamics of Aedes albopictus and Culex pipiens. Parasites Vectors 2017, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.B.; Eisen, L.; McAllister, J.; Savage, H.M.; Mutebi, J.P.; Eisen, R.J. Updated reported distribution of Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in the United States, 1995–2016. J. Med. Entomol. 2017, 54, 1420–1424. [Google Scholar] [CrossRef] [PubMed]
- Westby, K.M.; Adalsteinsson, S.A.; Biro, E.G.; Beckermann, A.J.; Medley, K.A. Aedes albopictus populations and larval habitat characteristics across the landscape: Significant differences exist between urban and rural land use types. Insects 2021, 161, 196. [Google Scholar] [CrossRef]
- Juliano, S.A.; Lounibos, P.L. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef] [Green Version]
- Little, E.; Biehler, D.; Leisnham, P.T.; Jordan, R.; Wilson, S.; LaDeau, S.L. Socio-ecological mechanisms supporting high densities of Aedes albopictus (Diptera: Culicidae) in Baltimore, MD. J. Med. Entomol. 2017, 54, 1183–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.P. Invasions by insect vectors of human disease. Ann. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Leisnham, P.T.; Biehler, D.; Bodner, D. Higher mosquito production in low-income neighborhoods of Baltimore and Washington, DC: Understanding ecological drivers and mosquito-borne disease risk in temperate Cities. Int. J. Environ. Res. Public Health 2013, 10, 1505–1526. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, K.S.; Mormann, K.; Juliano, S.A. Asymmetrical competition and patterns of abundance of Aedes albopictus and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2005, 42, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, K.S.; Muturi, E.J.; Lampman, H.L.; Alto, B.W. The effects of resource type and ratio on competition with Aedes albopictus and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, M.; Bacchi, M.; Bellini, R.; Maini, S. On the competition occurring between Aedes albopictus and Culex pipiens (Diptera: Culicidae) in Italy. Environ. Entomol. 2003, 32, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Müller, R.; Knautz, T.; Vollroth, S.; Berger, R.; Kreb, A.; Reuss, F.; Groneberg, D.A.; Kuch, U. Larval superiority of Culex pipiens to Aedes albopictus in a replacement series experiment: Prospects for coexistence in Germany. Parasites Vectors 2018, 11, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, Z.; Ladeau, S.L.; Armbruster, P.; Biehler, D.; Leisnham, P.T. Socioeconomic status affects mosquito (Diptera: Culicidae) larval habitat type availability and infestation level. J. Med. Entomol. 2013, 50, 764–772. [Google Scholar] [CrossRef]
- Facon, B.; Genton, B.J.; Shykoff, J.A.; Jarne, P.; Estoup, A.; David, P. A general eco-evolutionary framework for understanding bioinvasions. Trends Ecol. Evol. 2006, 21, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.L.; Holway, D.A. Condition-specific competition between invasive Argentine ants and Australian Iridomyrmex. J. Anim. Ecol. 2005, 74, 532–542. [Google Scholar] [CrossRef]
- Costanzo, K.S.; Kesavaraju, B.; Juliano, S.A. Condition specific competition in container mosquitoes: The role of non-competing life-history stages. Ecology 2005, 86, 3289–3295. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.J.; Robson, M.G.; Fonseca, D.M. Unexpected spatiotemporal abundance of infected Culex restuans suggest a greater role as a West Nile virus vector for this native species. Infect. Genet. Evol. 2015, 31, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Kilpatrick, M.A. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, A.M.; Fonseca, D.M.; Ebel, G.D.; Reddy, M.R.; Kramer, L.D. Spatial and temporal variation in vector competence of Culex pipiens and Cx. restuans mosquitoes for West Nile virus. Am. J. Trop. Med. Hyg. 2010, 83, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Sardelis, M.R.; Turell, M.J.; O’Guinn, M.L.; Andre, R.G.; Roberts, D.R. Vector competence of three North American strains of Aedes albopictus for West Nile virus. J. Am. Mosq. Contr. Assoc. 2002, 18, 284–289. [Google Scholar]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative host feeding patterns of the Asian tiger mosquito, Aedes albopictus, in urban and suburban northeastern USA and implications for disease transmission. PLoS Neglect. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [Green Version]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’Guinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Farajollahi, A.; Nelder, M.P. Changes in Aedes albopictus (Diptera: Culicidae) populations in New Jersey and implications for arbovirus transmission. J. Med. Entomol. 2009, 46, 1220–1224. [Google Scholar] [CrossRef] [Green Version]
- Mercer, D.R.; Sheeley, S.L.; Brown, E.J. Mosquito (Diptera : Culicidae) development within microhabitats of an Iowa wetland. J. Med. Entomol. 2005, 42, 685–693. [Google Scholar] [CrossRef]
- Isbell, F.I.; Mercer, D.R.; Mohammad, Z.I. Reliability of water quality test strips for field measurement of nutrients essential for developing mosquito larvae. Am. J. Undergrad. Res. 2006, 5, 1–8. [Google Scholar] [CrossRef]
- Johnson, B.J.; Sukhdeo, M. Successional mosquito dynamics in surrogate treehole and ground-container habitats in the northeastern United States: Where does Aedes albopictus fit in? J. Vect. Ecol. 2013, 38, 168–174. [Google Scholar] [CrossRef]
- Service, M.W. Population dynamics and mortalities of mosquito preadults. In Ecology of Mosquitoes, Proceedings of a Workshop; Lounibos, L.P., Rey, J.R., Frank, J.H., Eds.; Medical Entomology Laboratory: Vero Beach, FL, USA, 1985; pp. 185–201. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; Freeman and Company: New York, NY, USA, 1995; p. 850. [Google Scholar]
- Goldberg, D.E.; Scheiner, S.M. ANOVA and ANCOVA: Field competition experiments. In Design and Analysis of Ecological Experiments, 2nd ed.; Scheiner, S.M., Gurevitch, J., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 77–98. [Google Scholar]
- Scheiner, S.M. MANOVA: Multiple response variables and multispecies interactions. In Design and Analysis of Ecological Experiments; Scheiner, S.M., Gurevitch, J., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 99–115. [Google Scholar]
- Juliano, S.A. Species introduction and replacement among mosquitoes: Interspecific resource competition or apparent competition? Ecology 1998, 79, 255–268. [Google Scholar] [CrossRef]
- Yee, D.A.; Juliano, S.A. Consequences of detritus type in an aquatic microsystem: Effects on water quality, micro-organisms and performance of the dominant consumer. Fresh. Biol. 2006, 51, 448–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.; O’Meara, G.; Escher, R.; Nishimura, N.; Cutwa, M.; Nelson, T.; Campos, R.; Juliano, S. Testing predictions of displacement of native Aedes by the invasive Asian Tiger Mosquito Aedes albopictus in Florida, USA. Biol. Inv. 2001, 3, 151–166. [Google Scholar] [CrossRef]
- Kesavaraju, B.; Damal, K.; Juliano, S.A. Do natural container habitats impede invader dominance? Predator-mediated coexistence of invasive and native container-dwelling mosquitoes. Oecologia 2008, 155, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Freed, T.Z.; Kesavaraju, B.; Leisnham, P.T. Effects of competition and predation by native mosquitoes on the North American invasion of Aedes japonicus japonicus (Diptera: Culicidae). J. Med. Entomol. 2014, 51, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Freed, T.Z.; Leisnham, P.T. Roles of spatial partitioning, competition, and predation in the North American invasion of an exotic mosquito. Oecologia 2014, 175, 601–611. [Google Scholar] [CrossRef]
- Aliabadi, B.W.; Juliano, S.A. Escape from gregarine parasites affects the competitive interactions of an invasive mosquito. Biol. Inv. 2002, 4, 283–297. [Google Scholar] [CrossRef]
- Bodner, D.; LaDeau, S.L.; Leisnham, P.T. Relationships among immature-stage metrics and adult abundances of mosquito populations in Baltimore, MD. J. Med. Entomol. 2019, 59, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.M. Improving the Surveillance and Control of Vector Mosquitoes in Heterogenous Landscapes. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2020. [Google Scholar]
- Unlu, I.; Farajollahi, A.; Strickman, D.; Fonseca, D.M. Crouching tiger, hidden trouble: Urban sources of Aedes albopictus (Diptera: Culicidae) refractory to source-reduction. PLoS ONE 2013, 8, e77999. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Jöst, A.; Weitzel, T. The Culex pipiens complex in Europe. J. Am. Mosq. Control. Assoc. 2012, 28. [Google Scholar] [CrossRef] [PubMed]
- Dowling, Z.; Armbruster, P.; LaDeau, S.L.; DeCotiis, M.; Mottley, J.; Leisnham, P.T. Linking mosquito infestation to resident socioeconomic status, knowledge, and source reduction practices in suburban Washington, DC. EcoHealth 2013, 10, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E.B. Culex pipiens pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control; Pensoft: Sofia, Bulgaria, 2000. [Google Scholar]
- Ishii, T.; Sohn, S.R. Highly polluted larval habitats of the Culex pipiens complex in central Sweden. J. Am. Mosq. Control. Assoc. 1987, 3, 276–281. [Google Scholar] [PubMed]
- Allgood, D.W.; Yee, D.A. Oviposition preference and offspring performance in container breeding mosquitoes: Evaluating the effects of organic compounds and laboratory colonisation. Ecol. Entomol. 2017, 42, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, B.T.; Paulson, S.L. Seasonal abundance of Culex restuans and Culex pipiens in southwestern Virginia through ovitrapping. J. Am. Mosq. Control. Assoc. 2006, 22, 206–212. [Google Scholar] [CrossRef]
- Andreadis, T.G.; Anderson, J.F.; Vossbrinck, C.R.; Main, A.J. Epidemiology of West Nile virus in Connecticut: A five-year analysis of mosquito data 1999–2003. Vector-Borne Zoonotic Dis. 2004, 4, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, R.R.; Gottfried, K.L.; Apperson, C.S.; Davis, B.S.; Erwin, P.C.; Smith, A.B.; Panella, N.A.; Powell, E.E.; Nasci, R.S. First isolation of La Crosse virus from naturally infected Aedes albopictus. Emerg. Infect. Dis. 2001, 7, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization/World Health Organization. Zika Epidemiological Update–21 April 2016; PAHO/WHO: Washington, DC, USA, 2016. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Observed Density Per Container | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Container Type | Total Number of Water-Filled Containers | Sampled Containers | Aedes albopictus | Culex pipiens | Aedes albopictus + Culex pipiens | Neither | Proportion Culex pipiens (N) | Mean Density (Larvae/mL) | SD |
| Early season (May) | |||||||||
| Bucket | 38 | 5 | 2 | 3 | 0 | 0 | 0.692 (45) | 0.065 | 0.045 |
| Fence pole | 78 | 11 | 2 | 2 | 3 | 4 | 0.813 (87) | 0.149 | 0.286 |
| Trash can | 90 | 14 | 1 | 11 | 1 | 1 | 0.907 (567) | 0.284 | 0.218 |
| Discarded plastic | 203 | 32 | 1 | 18 | 4 | 9 | 0.957 (667) | 0.161 | 0.175 |
| Dumped tire | 75 | 46 | 2 | 27 | 7 | 10 | 0.921 (1136) | 0.242 | 0.375 |
| Discarded styrofoam | 77 | 7 | 0 | 2 | 0 | 5 | 0.936 (117) | 0.558 | 0.323 |
| Middle season (July–August) | |||||||||
| Bucket | 41 | 19 | 7 | 3 | 5 | 4 | 0.621 (174) | 0.154 | 0.163 |
| Fence pole | 85 | 25 | 15 | 0 | 1 | 9 | 0.013 (147) | 0.295 | 0.491 |
| Trash can | 56 | 18 | 6 | 5 | 7 | 0 | 0.556 (266) | 0.197 | 0.258 |
| Discarded plastic | 112 | 46 | 21 | 4 | 17 | 4 | 0.332 (449) | 0.366 | 0.554 |
| Dumped tire | 72 | 59 | 29 | 3 | 22 | 5 | 0.162 (308) | 0.316 | 0.359 |
| Discarded styrofoam | 50 | 12 | 10 | 0 | 1 | 1 | 0.116 (41) | 0.886 | 0.816 |
| Late season (September) | |||||||||
| Bucket | 46 | 11 | 4 | 2 | 5 | 0 | 0.257 (38) | 0.217 | 0.181 |
| Fence pole | 129 | 27 | 26 | 1 | 0 | 0 | 0.005 (1) | 0.154 | 0.198 |
| Trash can | 71 | 15 | 5 | 4 | 6 | 0 | 0.455 (80) | 0.163 | 0.183 |
| Discarded plastic | 99 | 11 | 6 | 0 | 4 | 1 | 0.276 (214) | 0.234 | 0.151 |
| Dumped tire | 57 | 27 | 13 | 4 | 10 | 0 | 0.255 (130) | 0.228 | 0.232 |
| Discarded styrofoam | 31 | 0 | - | - | - | - | - | - | - |
| Nutrient | Test Strip (mg/L) | Spectrophotometer (mg/L) | df | t | P | r | p | ||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||||||
| Nitrate | 5.21 | 10.34 | 15.64 | 19.21 | 98 | −9.30 | <0.0001 | 0.885 | <0.0001 |
| Ammonia | 0.67 | 0.99 | 1.52 | 1.95 | 94 | −6.82 | <0.0001 | 0.855 | <0.0001 |
| Phosphate | 15.96 | 16.09 | 8.80 | 14.44 | 93 | 6.89 | <0.0001 | 0.787 | <0.0001 |
| Canonical Variates | Standardized Canonical Coefficients | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | Pillai’s Trace (F) | df | p | Variate Number | Percent Variation | p | Nitrate | Ammonia | Phosphate |
| Container type | 1.87 | 15, 54 | 0.0481 | 1 | 95.6 | 0.0017 | 0.89 | 1.42 | −0.53 |
| 2 | 3.4 | 0.8680 | 0.72 | −0.76 | 0.88 | ||||
| 3 | 1.0 | 0.8314 | −1.71 | −1.62 | 0.41 | ||||
| Functional vs. Discarded | 9.86 | 3, 16 | 0.0006 | 1 | 100 | 0.0006 | 0.69 | 1.60 | −0.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leisnham, P.T.; LaDeau, S.L.; Saunders, M.E.M.; Villena, O.C. Condition-Specific Competitive Effects of the Invasive Mosquito Aedes albopictus on the Resident Culex pipiens among Different Urban Container Habitats May Explain Their Coexistence in the Field. Insects 2021, 12, 993. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12110993
Leisnham PT, LaDeau SL, Saunders MEM, Villena OC. Condition-Specific Competitive Effects of the Invasive Mosquito Aedes albopictus on the Resident Culex pipiens among Different Urban Container Habitats May Explain Their Coexistence in the Field. Insects. 2021; 12(11):993. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12110993
Chicago/Turabian StyleLeisnham, Paul T., Shannon L. LaDeau, Megan E. M. Saunders, and Oswaldo C. Villena. 2021. "Condition-Specific Competitive Effects of the Invasive Mosquito Aedes albopictus on the Resident Culex pipiens among Different Urban Container Habitats May Explain Their Coexistence in the Field" Insects 12, no. 11: 993. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12110993