
Evaluation of Natural and Factitious Food Sources for Pronematus ubiquitus on Tomato Plants

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mites and Plants
2.2. Experiment 1: Pronematus ubiquitus Strain Comparison
2.3. Experiments 2 and 3: Oviposition of P. ubiquitus on Alternative Foods
2.4. Experiment 4: Frequency of Application of Typha Angustifolia for Pre-Establishing P. ubiquitus
3. Results
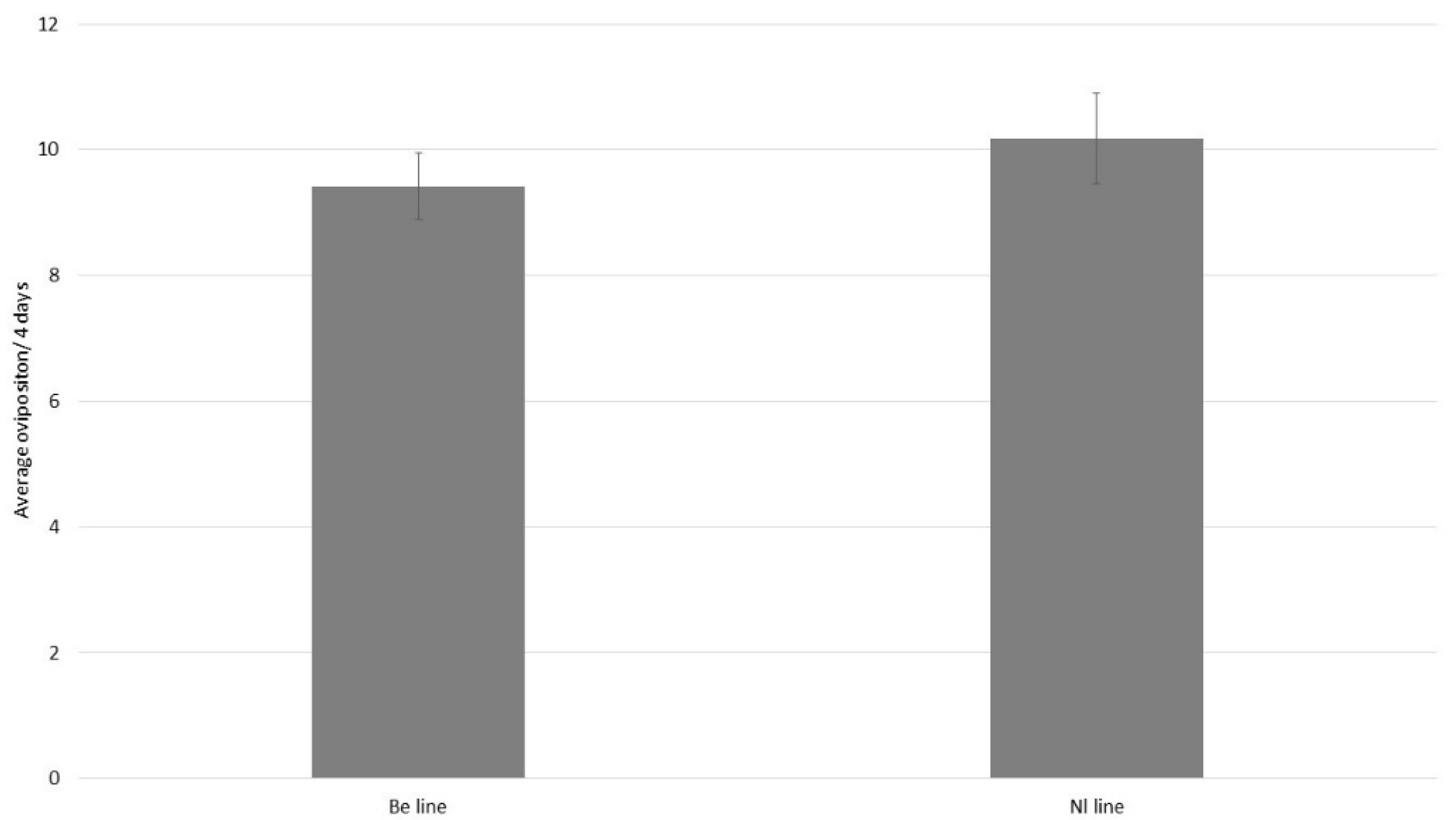
3.1. Experiment 1: Pronematus ubiquitus Strain Comparison
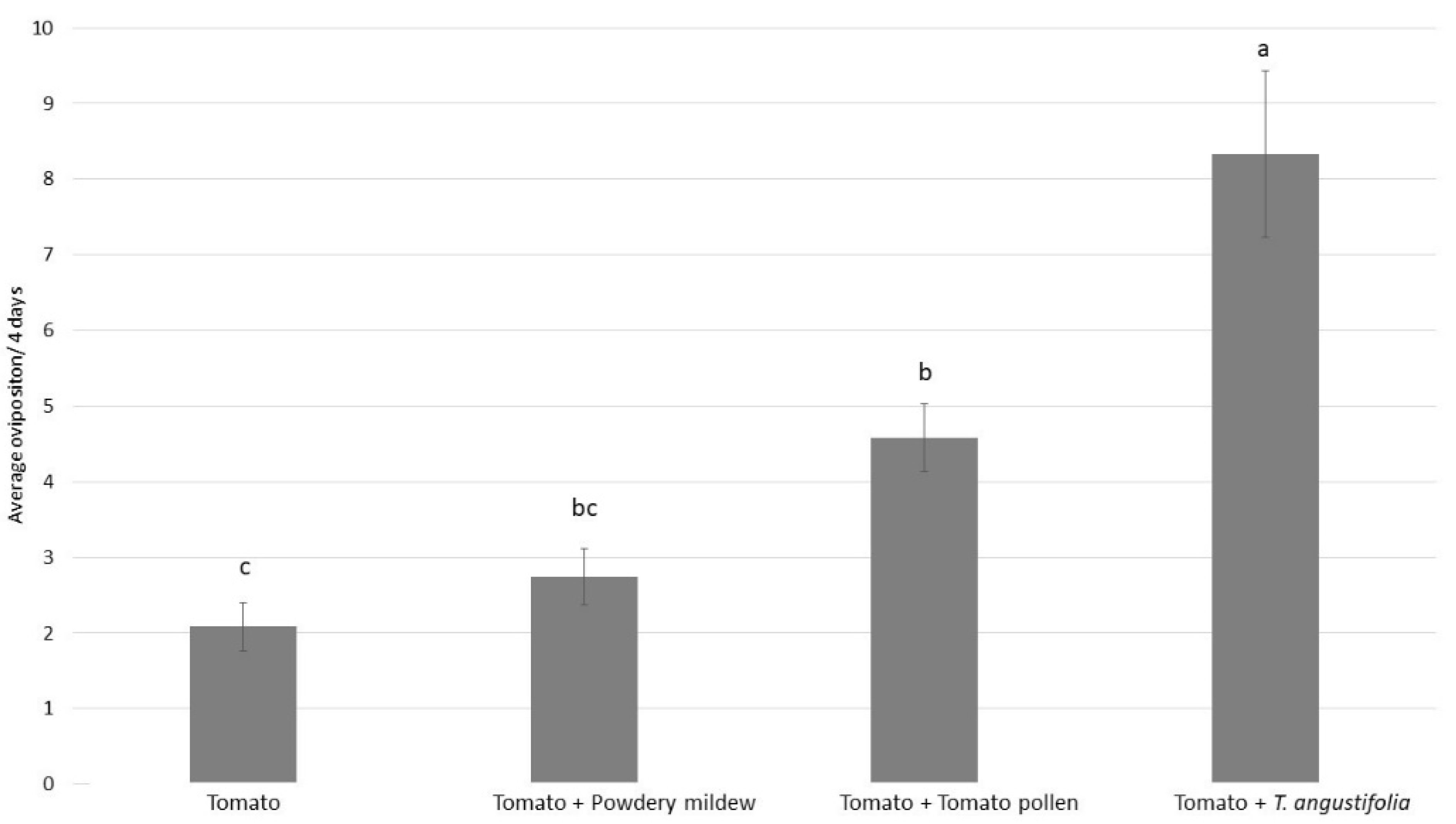
3.2. Experiment 2: Oviposition of P. ubiquitus on Naturally Occurring Food Sources on Tomato Crops
3.3. Experiment 3: Oviposition of P. ubiquitus on Factitious Food Sources
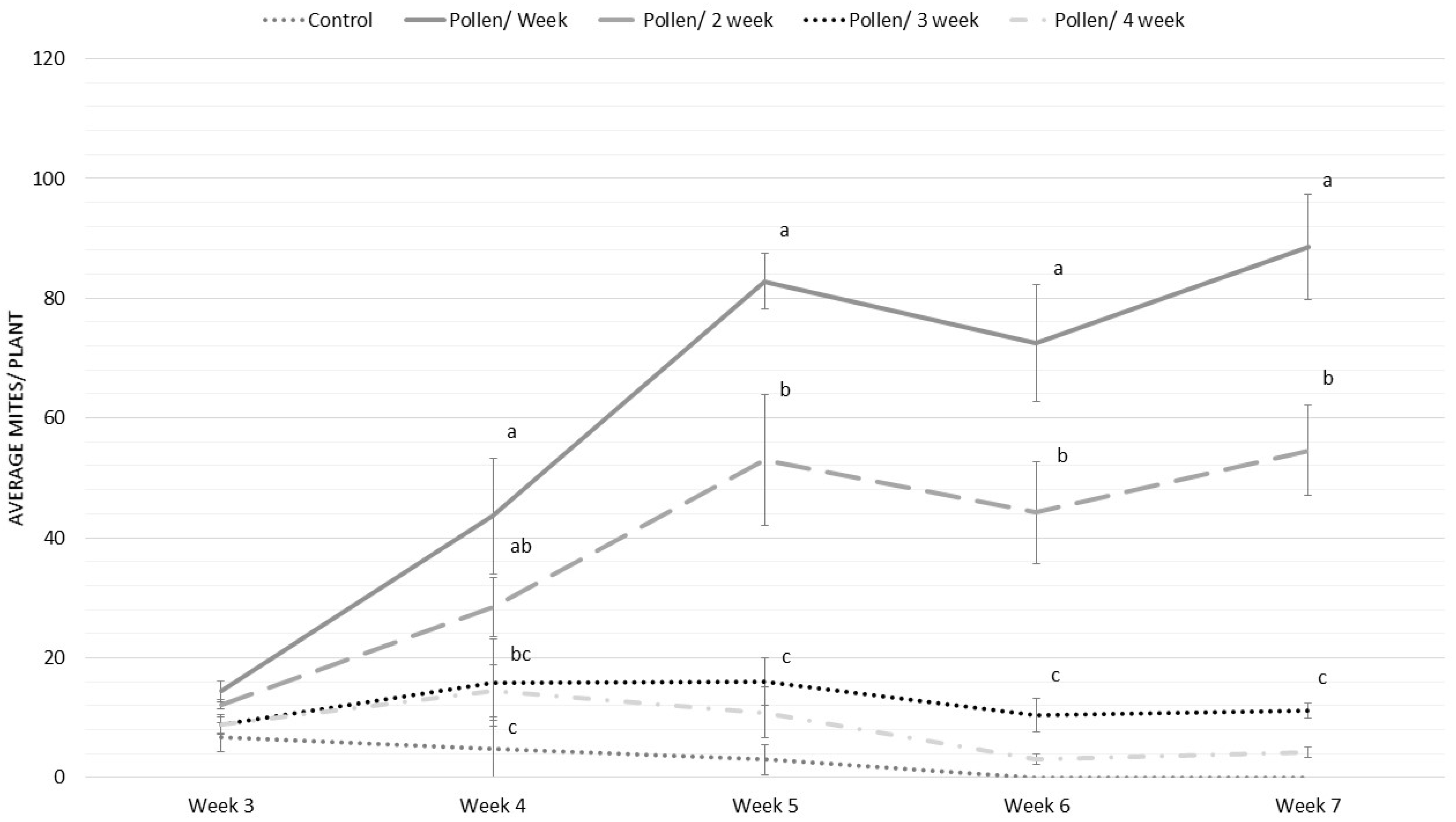
3.4. Experiment 4: Frequency of Application of T. angustifolia for Pre-Establishing P. ubiquitus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can Generalist Predators be Effective Biocontrol Agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messelink, G.J.; Bennison, J.; Alomar, O.; Ingegno, B.L.; Tavella, L.; Shipp, L.; Palevsky, E.; Wäckers, F.L. Approaches to conserving natural enemy populations in greenhouse crops: Current methods and future prospects. BioControl 2014, 59, 377–393. [Google Scholar] [CrossRef]
- Pijnakker, J.; Moerkens, R.; Vangansbeke, D.; Duarte, M.; Bellinkx, S.; Benavente, A.; Merckx, J.; Stevens, I.; Wäckers, F. Dual protection: A tydeoid mite effectively controls both a problem pest and a key pathogen in tomato. Pest Manag. Sci. 2021, 78, 355–361. [Google Scholar] [CrossRef]
- Messelink, G.J.; van Maanen, R.; van Steenpaal, S.E.; Janssen, A. Biological control of thrips and whiteflies by a shared predator: Two pests are better than one. Biol. Control 2008, 44, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Leman, A.; Ingegno, B.L.; Tavella, L.; Janssen, A.; Messelink, G.J. The omnivorous predator Macrolophus pygmaeus, a good candidate for the control of both greenhouse whitefly and poinsettia thrips on gerbera plants. Insect Sci. 2020, 27, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Goleva, I.; Zebitz, C.P.W. Suitability of different pollen as alternative food for the predatory mite Amblyseius swirskii (Acari, Phytoseiidae). Exp. Appl. Acarol. 2013, 61, 259–283. [Google Scholar] [CrossRef]
- Mendoza, J.E.; Balanza, V.; Cifuentes, D.; Bielza, P. Genetic improvement of Orius laevigatus for better fitness feeding on pollen. J. Pest Sci. 2021, 94, 729–742. [Google Scholar] [CrossRef]
- Leman, A.; Messelink, G.J. Supplemental food that supports both predator and pest: A risk for biological control? Exp. Appl. Acarol. 2015, 65, 511–524. [Google Scholar] [CrossRef]
- Wäckers, F.L. Suitability of (Extra-) Floral Nectar, Pollen, and Honeydew as Insect Food Sources. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Cambridge University Press: Cambridge, UK, 2005; pp. 17–74. [Google Scholar]
- Duarte, M.V.A.; Venzon, M.; Bittencourt, M.C.D.S.; Rodriguez-Cruz, F.A.; Pallini, A.; Janssen, A. Alternative food promotes broad mite control on chilli pepper plants. Biocontrol 2015, 60, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Xiao, Y.; McKenzie, C.L.; Osborne, L.S. Early establishment of the phytoseiid mite Amblyseius swirskii (Acari: Phytoseiidae) on pepper seedlings in a Predator-in-first approach. Exp. Appl. Acarol. 2015, 65, 465–481. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Nguyen, D.T.; Audenaert, J.; Gobin, B.; Tirry, L.; De Clercq, P. Establishment of Amblyseius swirskii in greenhouse crops using food supplements. Syst. Appl. Acarol. 2016, 21, 1174–1184. [Google Scholar] [CrossRef]
- McGregor, R.; Crisp, K.; Castiglia, C. Feeding lifestyles of the Phytoseiidae revisited: Searching for a factitious rearing host for Neoseiulus fallacis (Acari: Phytoseiidae). BioControl 2020, 65, 593–599. [Google Scholar] [CrossRef]
- Calvo, F.J.; Knapp, M.; van Houten, Y.M.; Hoogerbrugge, H.; Belda, J.E. Amblyseius swirskii: What made this predatory mite such a successful biocontrol agent? Exp. Appl. Acarol. 2015, 65, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Massaro, M.; Montrazi, M.; Melo, J.W.S.; de Moraes, G.J. Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae). Insects 2021, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Knapp, M.; van Houten, Y.; van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: Current status and future prospects. Acarologia 2018, 58, 72–82. [Google Scholar] [CrossRef]
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ramakers, P.M.J. Biological control of Thrips tabaci (Thysanoptera: Thripidae) with Amblyseius spp. (Acari: Phytoseiidae). Bull. SROP 1980, 3, 203–207. [Google Scholar]
- Messelink, G.J.; van Steenpaal, S.E.F.; Ramakers, P.M.J. Evaluation of phytoseiid predators for control of western flower thrips on greenhouse cucumber. BioControl 2006, 51, 753–768. [Google Scholar] [CrossRef]
- Palevsky, E.; Walzer, A.; Gal, S.; Schausberger, P. Evaluation of dry-adapted strains of the predatory mite Neoseiulus californicus for spider mite control on cucumber, strawberry and pepper. Exp. Appl. Acarol. 2008, 45, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Control of Bemisia tabaci and Frankliniella occidentalis in cucumber by Amblyseius swirskii. BioControl 2011, 56, 185–192. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Biological control-based IPM in sweet pepper greenhouses using Amblyseius swirskii (Acari: Phytoseiidae). Biocontrol Sci. Technol. 2012, 22, 1398–1416. [Google Scholar] [CrossRef]
- Labbé, R.M.; Gagnier, D.; Shipp, L. Comparison of Transeius montdorensis Acari: Phytoseiidae) to other phytoseiid mites for the short-season suppression of western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Environ. Entomol. 2019, 48, 335–342. [Google Scholar] [CrossRef]
- Park, H.-H.; Shipp, L.; Buitenhuis, R.; Ahn, J.J. Life history parameters of a commercially available Amblyseius swirskii (Acari: Phytoseiidae) fed on cattail (Typha latifolia) pollen and tomato russet mite (Aculops lycopersici). J. Asia-Pac. Entomol. 2011, 14, 497–501. [Google Scholar] [CrossRef]
- Paspati, A.; Rambla, J.L.; Gresa, M.P.L.; Arbona, V.; Gómez-Cadenas, A.; Granell, A.; González-Cabrera, J.; Urbaneja, A. Tomato trichomes are deadly hurdles limiting the establishment of Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Biol. Control 2021, 157, 104572. [Google Scholar] [CrossRef]
- Vervaet, L.; De Vis, R.; de Clercq, P.; van Leeuwen, T. Is the emerging mite pest Aculops lycopersici controllable? Global and genome-based insights in its biology and management. Pest Manag. Sci. 2021, 77, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Drukker, B.; Janssen, A.; Ravensberg, W.; Sabelis, M.W. Improved control capacity of the mite predator Phytoseiulus persimilis (Acari: Phytoseiidae) on tomato. Exp. Appl. Acarol. 1997, 21, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Van Haren, R.J.; Steenhuis, M.M.; Sabelis, M.W.; De Ponti, O.D. Tomato stem trichomes and dispersal success of Phytoseiulus persimilis relative to its prey Tetranychus urticae. Exp. Appl. Acarol. 1987, 3, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Kawai, A.; Haque, M.M. Population dynamics of tomato russet mite, Aculops lycopersici (Massee) and its natural enemy, Homeopronematus anconai (Baker). Jarq-Jpn. Agr. Res. Q. 2004, 38, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Aysan, E.; Kumral, N.A. Tritrophic relationships among tomato cultivars, the rust mite, Aculops lycopersici (Massee) (Eriophyidae), and its predators. Acarologia 2018, 58, 5–17. [Google Scholar] [CrossRef]
- Van Houten, Y.M.; Hoogerbrugge, H.; Knapp, M. Potential of Pronematus ubiquitus to control tomato russet mite, Aculops lycopersici. IOBC/WPRS Bull. 2020, 149, 87–92. [Google Scholar]
- Baradaran, P.; Arbabi, M. Population abundance of Pronematus ubiquitus (McGregor, 1932) (Acari: Tydeidae) on different fig varieties. J. Entomol. Res. 2009, 1, 177–183. [Google Scholar]
- Dean, H.A. Predators of Oligonychus pratensis (Banks), Tetranychidae. Ann. Entomol. Soc. Am. 1957, 50, 164–165. [Google Scholar] [CrossRef]
- McCoy, C.W.; Selhime, A.G.; Kanavel, R.F. The Feeding Behavior and Biology of Parapronematus acaciae (Acarina: Tydeidae). Fla. Entomol. 1969, 52, 13. [Google Scholar] [CrossRef]
- Knop, N.F.; Hoy, M.A. Biology of a tydeid mite, Homeopronematus anconai (n. comb.) (Acari: Tydeidae), important in San Joaquin valley vineyards. Hilgardia 1983, 51, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Knop, N.F.; Hoy, M.A. Factors Limiting the Utility of Homeopronematus anconai (Acari: Tydeidae) in Integrated Pest Management in San Joaquin Valley Vineyards. J. Econ. Entomol. 1983, 76, 1181–1186. [Google Scholar] [CrossRef]
- Hessein, N.A.; Perring, T.M. The importance of alternate foods for the mite Homeopronematus anconai (Acari: Tydeidae). Ann. Entomol. Soc. Am. 1988, 81, 488–492. [Google Scholar] [CrossRef]
- Pijnakker, J.; Vangansbeke, D.; Duarte, M.; Moerkens, R.; Wäckers, F.L. Predators and Parasitoids-in-First: From Inundative Releases to Preventative Biological Control in Greenhouse Crops. Front. Sustain Food Syst. 2020, 4, 245. [Google Scholar] [CrossRef]
- Benson, C.M.; Labbe, R.M. Exploring the Role of Supplemental Foods for Improved Greenhouse Biological Control. Ann. Entomol. Soc. Am. 2021, 114, 302–321. [Google Scholar] [CrossRef]
- Van Rijn, P.C.J.; van Houten, Y.M.; Sabelis, M.W. How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 2002, 83, 2664–2679. [Google Scholar]
- Pijnakker, J.; Arijs, Y.; de Souza, A.; Cellier, M.; Wäckers, F. The use of Typha angustifolia (cattail) pollen to establish the predatory mites Amblyseius swirskii, Iphiseius degenerans, Euseius ovalis and Euseius gallicus in glasshouse crops, Integrated Control of Plant-FeedingMites. IOBC/WPRS Bull. 2016, 120, 47–52. [Google Scholar]
- Labbé, R.M.; Gagnier, D.; Kostic, A.; Shipp, L. The function of supplemental foods for improved crop establishment of generalist predators Orius insidiosus and Dicyphus hesperus. Sci. Rep. 2018, 8, 17790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenard, N.; Sluydts, V.; Christianen, E.; Bosmans, L.; De Bruyn, L.; Moerkens, R.; Liers, H. Biweekly supplementation with Artemia spp. cysts allows efficient population establishment by Macrolophus pygmaeus in sweet pepper. Entomol. Exp. Appl. 2019, 167, 406–414. [Google Scholar] [CrossRef]
- Hoogerbrugge, H.; van Houten, Y.; van Baal, E.; Bolckmans, K. Alternative food sources to enable establishment of Amblyseius swirskii (Athias-Henriot) on chrysanthemum without pest presence. IOBC/WPRS Bull. 2008, 32, 79. [Google Scholar]
- Pirayeshfar, F.; Safavi, S.A.; Sarraf Moayeri, H.R.; Messelink, G.J. The potential of highly nutritious frozen stages of Tyrophagus putrescentiae as a supplemental food source for the predatory mite Amblyseius swirskii. Biocontrol Sci. Technol. 2020, 30, 403–417. [Google Scholar] [CrossRef]
- Pirayeshfar, F.; Safavi, S.A.; Moayeri, H.R.S.; Messelink, G.J. Provision of astigmatid mites as supplementary food increases the density of the predatory mite Amblyseius swirskii in greenhouse crops, but does not support the omnivorous pest, western flower thrips. BioControl 2021, 66, 1–12. [Google Scholar] [CrossRef]
- Muñoz-Cárdenas, K.; Ersin, F.; Pijnakker, J.; van Houten, Y.; Hoogerbrugge, H.; Leman, A.; Janssen, A. Supplying high-quality alternative prey in the litter increases control of an above-ground plant pest by a generalist predator. Biological control 2017, 105, 19–26. [Google Scholar] [CrossRef]
- Li, L.; Jiao, R.; Yu, L.; He, X.Z.; He, L.; Xu, C.; Zhang, L.; Liu, J. Functional response and prey stage preference of Neoseiulus barkeri on Tarsonemus confusus. System. Appl. Acarol. 2018, 23, 2244–2258. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Duarte, M.V.; Merckx, J.; Benavente, A.; Magowski, W.L.; França, S.C.; Wäckers, F.L. Impact of a tarsonemid prey mite and its fungal diet on the reproductive performance of a predatory mite. Exp. Appl. Acarol. 2021, 83, 313–323. [Google Scholar] [CrossRef]
- Farfan, M.A.; Coffey, J.; Schmidt-Jeffris, R.A. Evaluation of Tarsonemus bilobatus and Podosphaera xanthii as suitable resources for Proprioseiopsis mexicanus in cucurbit systems in the Southeast USA. Exp. Appl. Acarol. 2021, 85, 31–40. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Chichester, UK, 2007; p. 950. [Google Scholar]
- Lenth, R. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Core Team, R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Castagnoli, M.; Simoni, S.; Nachman, G. Short-term changes in consumption and oviposition rates of Neoseiulus californicus strains (Acari: Phytoseiidae) after a diet shift. Exp. Appl. Acarol. 2001, 25, 969–983. [Google Scholar] [CrossRef]
- Perring, T.M.; Lackey, L.J. Temperature and humidity effects on mortality and pre-adult development of two Phytoseiulus persimilis strains (Acari: Phytoseiidae). Int. J. Acarol. 1989, 15, 47–52. [Google Scholar] [CrossRef]
- Samaras, K.; Pappas, M.L.; Fytas, E.; Broufas, G.D. Pollen suitability for the development and reproduction of Amblydromalus limonicus (Acari: Phytoseiidae). Biocontrol 2015, 60, 773–782. [Google Scholar] [CrossRef]
- Van Rijn, P.C.J.; Tanigoshi, L.K. Pollen as food for the predatory mites Iphiseius degenerans and Neoseiulus cucumeris (Acari: Phytoseiidae): Dietary range and life history. Exp. Appl. Acarol. 1999, 23, 785–802. [Google Scholar] [CrossRef]
- Bohs, L. Transfer of Cyphomandra (Solanaceae) and its species to Solanum. Taxon 1995, 44, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Buchmann, S.L.; Hurley, J.P. A biophysical model for buzz pollination in angiosperms. J. Theor. Biol. 1978, 72, 639–657. [Google Scholar] [CrossRef]
- Van Ravestijn, W.; Van der Sande, J. Use of bumblebees for the pollination of glasshouse tomatoes. In VI International Symposium on Pollination (1990, August) ISHS. Acta Hortic. 1991, 288, 204–212. [Google Scholar] [CrossRef]
- Poulton, J.L.; Koide, R.T.; Stephenson, A.G. Effects of mycorrhizal infection and soil phosphorus availability on in vitro and in vivo pollen performance in Lycopersicon esculentum (Solanaceae). Am. J. Bot. 2001, 88, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Shaked, R.; Peet, M.M.; Pharr, D.M.; Zamski, E.; Rosenfeld, K.; Pressman, E. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci. Hortic. 2006, 109, 212–217. [Google Scholar] [CrossRef]
- Lefebvre, D.; Pierre, J. Hive weight as an indicator of bumblebee colony growth. J. Apicult. Res. 2006, 45, 217–218. [Google Scholar] [CrossRef]
- English-Loeb, G.; Norton, A.P.; Gadoury, D.M.; Seem, R.C.; Wilcox, W.F. Control of Powdery Mildew in Wild and Cultivated Grapes by a Tydeid Mite. Biol. Control 1999, 14, 97–103. [Google Scholar] [CrossRef]
- English-Loeb, G.; Norton, A.P.; Gadoury, D.; Seem, R.; Wilcox, W. Biological control of grape powdery mildew using mycophagous mites. Plant Dis. 2007, 91, 421–429. [Google Scholar] [CrossRef]
- Pappas, M.L.; Steppuhn, A.; Broufas, G.D. The role of phytophagy by predators in shaping plant interactions with their pests. Commun. Integr. Biol. 2016, 9, e0127251. [Google Scholar] [CrossRef] [PubMed]
- Travis, J.; Keen, W.H.; Juilianna, J. The role of relative body size in a predator-prey relationship between dragonfly naiads and larval anurans. Oikos 1985, 45, 59–65. [Google Scholar] [CrossRef]
- Emmerson, M.C.; Raffaelli, D. Predator–prey body size, interaction strength and the stability of a real food web. J. Anim. Ecol. 2004, 73, 399–409. [Google Scholar] [CrossRef]
- Pappas, M.L.; Xanthis, C.; Samaras, K.; Koveos, D.S.; Broufas, G.D. Potential of the predatory mite Phytoseius finitimus (Acari: Phytoseiidae) to feed and reproduce on greenhouse pests. Exp. Appl. Acarol. 2013, 61, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Samaras, K.; Pappas, M.L.; Pekas, A.; Wäckers, F.; Broufas, G.D. Benefits of a balanced diet? Mixing prey with pollen is advantageous for the phytoseiid predator Amblydromalus limonicus. Biol. Control 2021, 155, 104531. [Google Scholar] [CrossRef]
- Asgari, F.; Moayeri, H.R.S.; Kavousi, A.; Enkegaard, A.; Chi, H. Demography and mass rearing of Amblyseius swirskii (Acari: Phytoseiidae) fed on two species of stored-product mites and their mixture. J. Econ. Entomol. 2020, 113, 2604–2612. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, M.V.A.; Vangansbeke, D.; Pijnakker, J.; Moerkens, R.; Benavente, A.; Arijs, Y.; Saucedo, A.L.F.; Wäckers, F. Evaluation of Natural and Factitious Food Sources for Pronematus ubiquitus on Tomato Plants. Insects 2021, 12, 1111. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121111
Duarte MVA, Vangansbeke D, Pijnakker J, Moerkens R, Benavente A, Arijs Y, Saucedo ALF, Wäckers F. Evaluation of Natural and Factitious Food Sources for Pronematus ubiquitus on Tomato Plants. Insects. 2021; 12(12):1111. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121111
Chicago/Turabian StyleDuarte, Marcus V. A., Dominiek Vangansbeke, Juliette Pijnakker, Rob Moerkens, Alfredo Benavente, Yves Arijs, Ana Lizbeth Flores Saucedo, and Felix Wäckers. 2021. "Evaluation of Natural and Factitious Food Sources for Pronematus ubiquitus on Tomato Plants" Insects 12, no. 12: 1111. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121111