
Life History Traits in Two Drosophila Species Differently Affected by Microbiota Diversity under Lead Exposure

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Laboratory Maintenance
2.2. Experimental Setup
2.3. Statistical Analysis of Life History Traits
2.4. Total DNA Extraction and 16S rRNA Sequencing
2.5. Amplicon Sequence Variant Inference and Taxonomy Assignment
2.6. Microbiome and Statistical Analysis
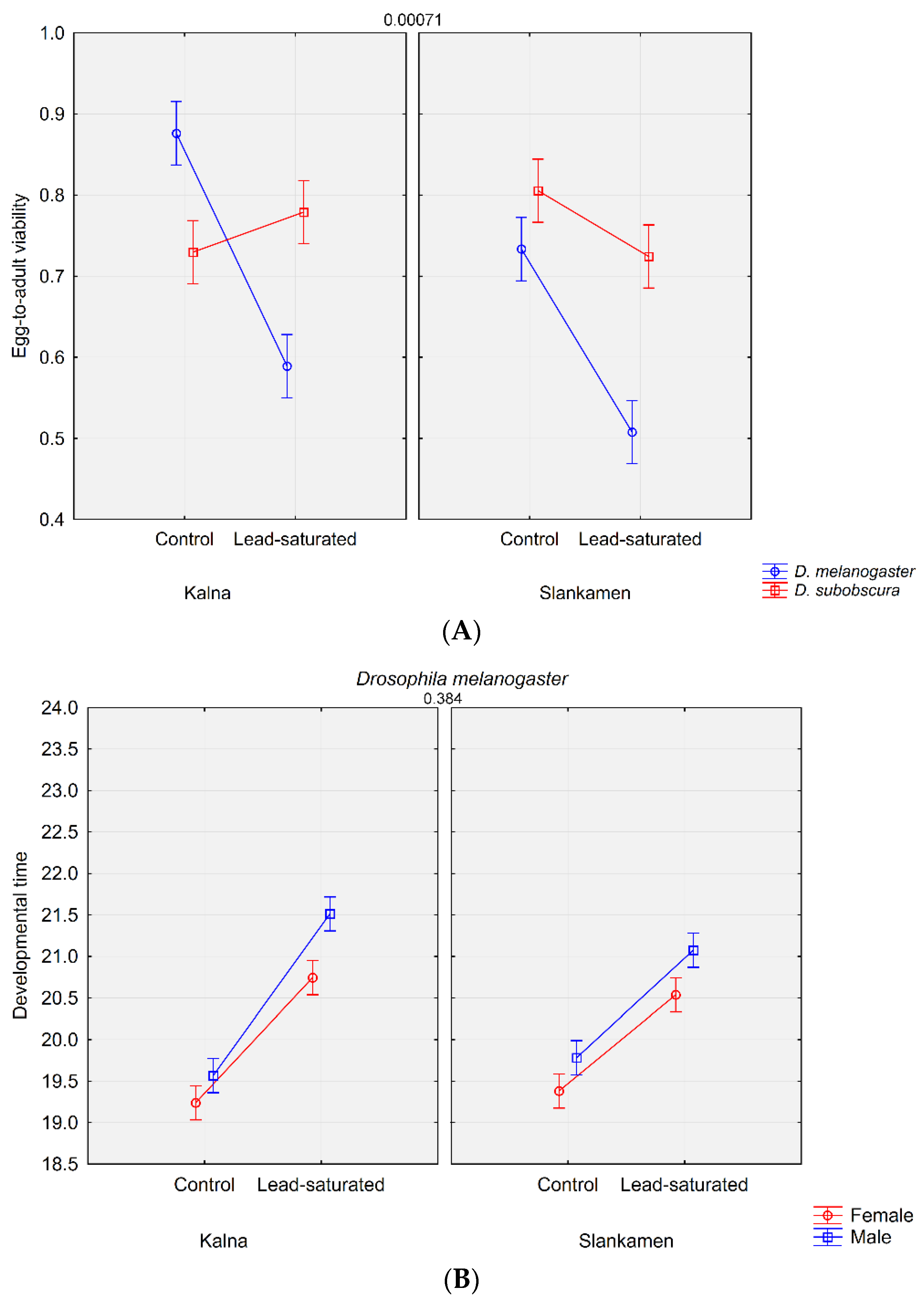
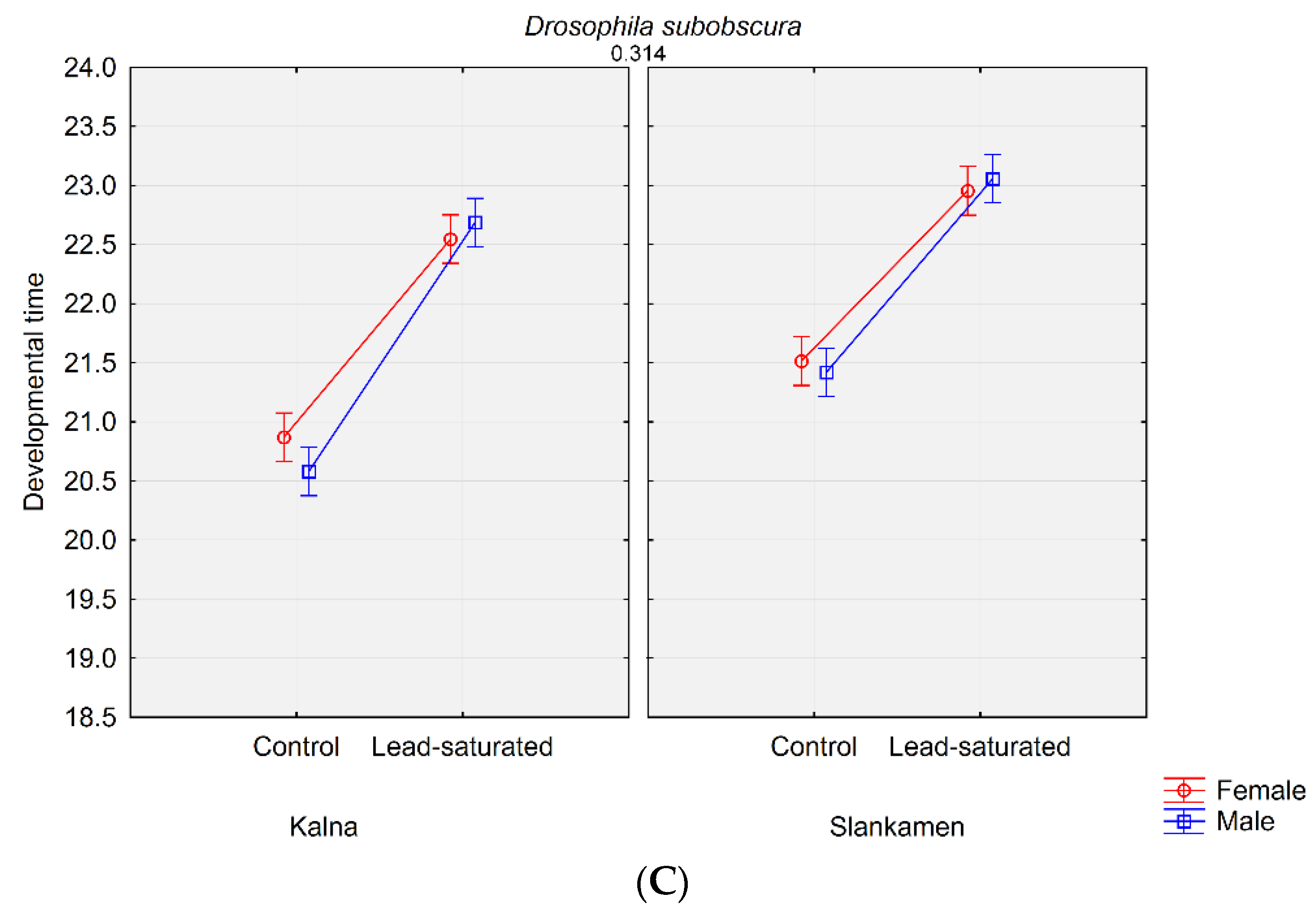
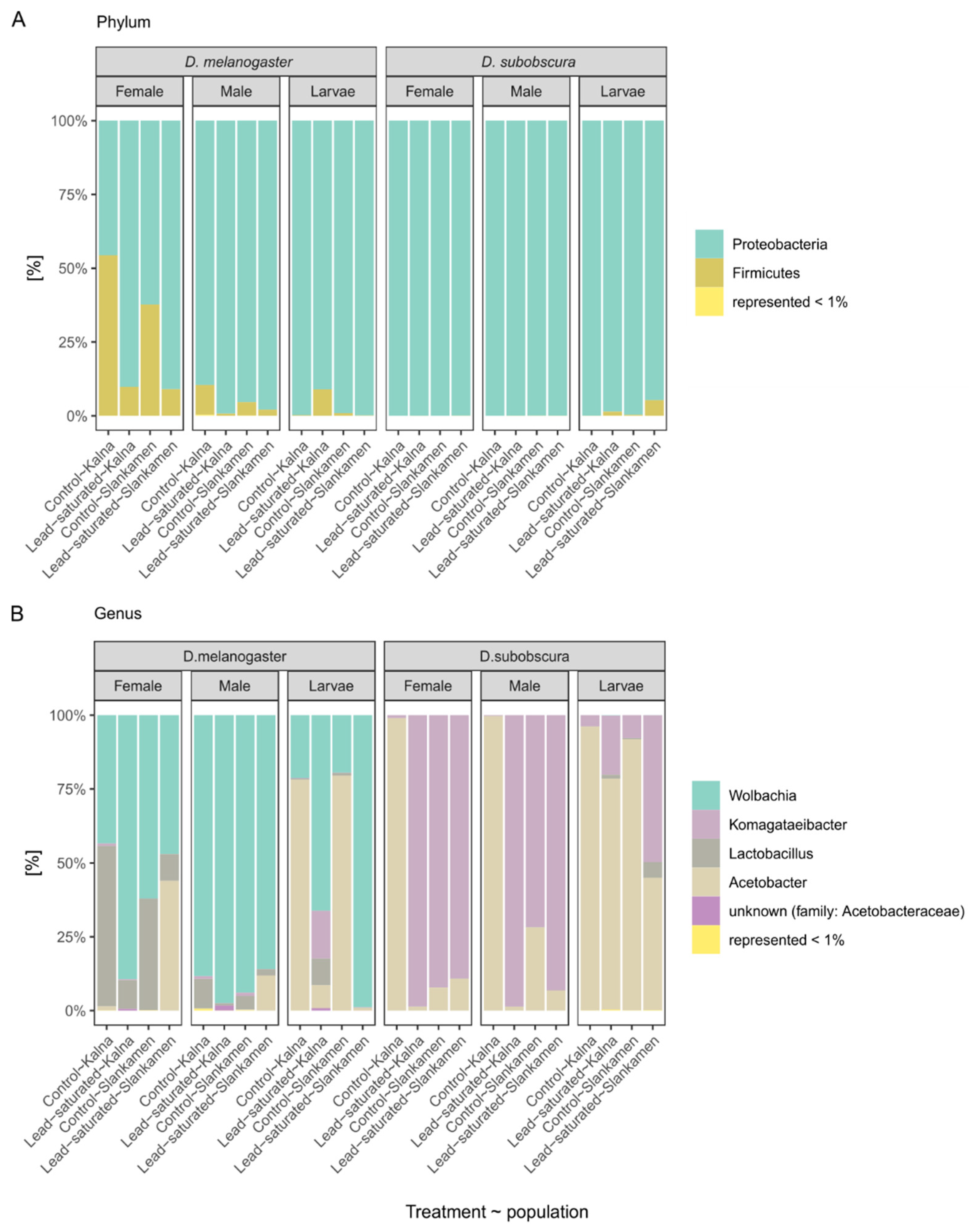
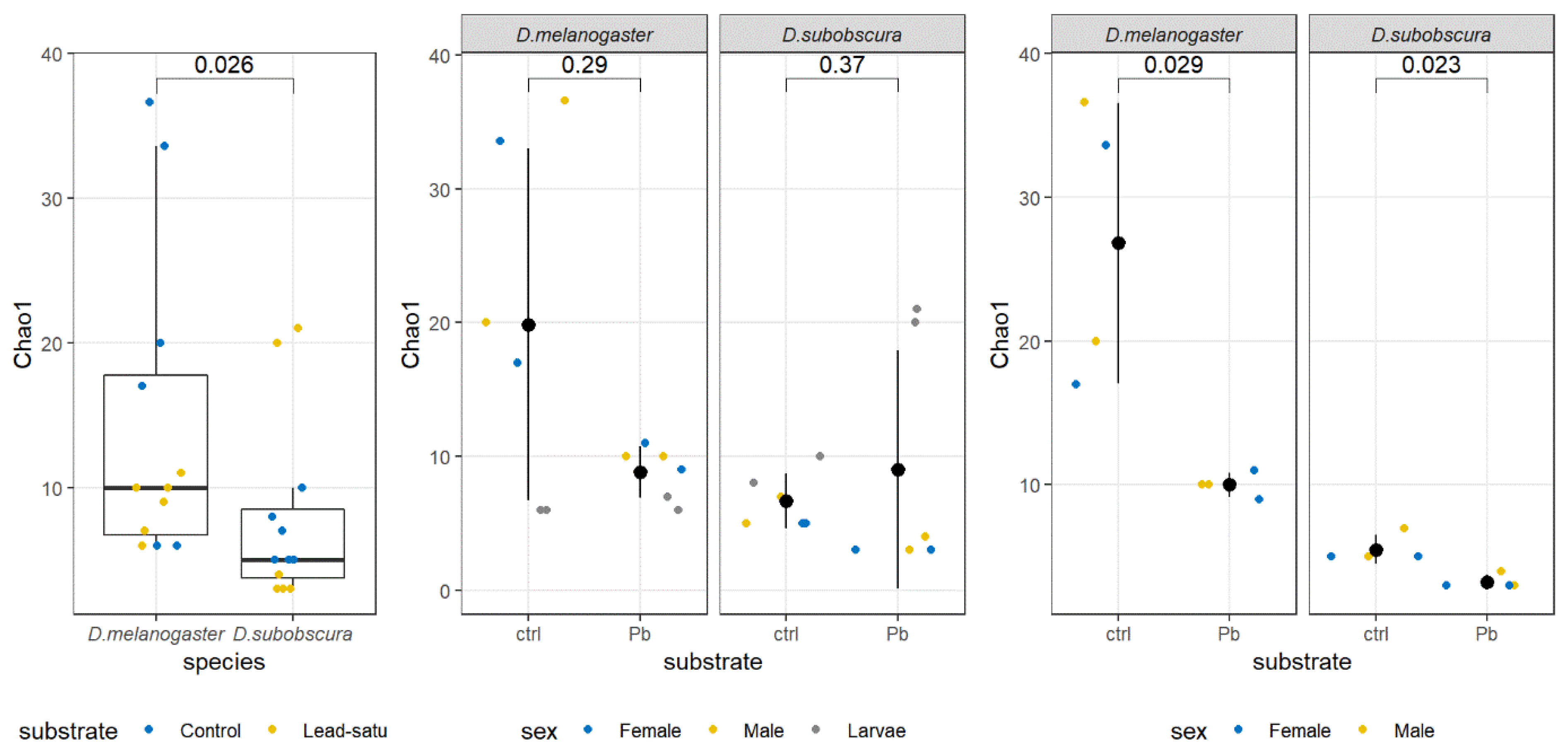
3. Results
3.1. Life History Traits Analysis
3.2. Taxonomic Abundance of Microbial Communities
3.3. Alpha Diversity Analysis
3.4. Beta Diversity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Wong, C.N.A.; Ng, P.; Douglas, A.E. Low-diversity bacterial community in the gut of the fruitfly Drosophila melanogaster. Environ. Microbiol. 2011, 13, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beribaka, M.B.; Dimkić, I.Z.; Jelić, M.Đ.; Stanković, S.M.; Pržulj, N.M.; Anđelković, M.L.; Stamenković-Radak, M.M. Altered diversity of bacterial communities in two Drosophila species under laboratory conditions and lead exposure. Arch. Biol. Sci. 2021, 73, 17–29. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Kim, S.-H.; Lee, H.-Y.; Bai, J.Y.; Nam, Y.-D.; Bae, J.-W.; Lee, D.G.; Shin, S.C.; Ha, E.-M.; Lee, W.-J. Innate Immune Homeostasis by the Homeobox Gene Caudal and Commensal-Gut Mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Iatsenko, I.; Boquete, J.-P.; Lemaitre, B. Microbiota-Derived Lactate Activates Production of Reactive Oxygen Species by the Intestinal NADPH Oxidase Nox and Shortens Drosophila Lifespan. Immunity 2018, 49, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Fast, D.; Duggal, A.; Foley, E. Monoassociation with Lactobacillus plantarum disrupts intestinal homeostasis in adult Drosophila melanogaster. MBio 2018, 9, e01114–e01118. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-Y.; Lee, S.-H.; Lee, J.-H.; Lee, W.-J.; Min, K.-J. The role of commensal microbes in the lifespan of Drosophila melanogaster. Aging 2019, 11, 4611–4640. [Google Scholar] [CrossRef] [PubMed]
- Leitão-Gonçalves, R.; Carvalho-Santos, Z.; Francisco, A.P.; Fioreze, G.T.; Anjos, M.; Baltazar, C.; Elias, A.P.; Itskov, P.M.; Piper, M.D.W.; Ribeiro, C. Commensal bacteria and essential amino acids control food choice behavior and reproduction. PLoS Biol. 2017, 15, e2000862. [Google Scholar] [CrossRef]
- Jia, Y.; Jin, S.; Hu, K.; Geng, L.; Han, C.; Kang, R.; Pang, Y.; Ling, E.; Tan, E.K.; Pan, Y.; et al. Gut microbiome modulates Drosophila aggression through octopamine signaling. Nat. Commun. 2021, 12, 2698. [Google Scholar] [CrossRef]
- Silva, V.; Palacios-Muñoz, A.; Okray, Z.; Adair, K.L.; Waddell, S.; Douglas, A.E.; Ewer, J. The impact of the gut microbiome on memory and sleep in Drosophila. J. Exp. Biol. 2021, 224. [Google Scholar] [CrossRef]
- Clark, R.I.; Salazar, A.; Yamada, R.; Fitz-Gibbon, S.; Morselli, M.; Alcaraz, J.; Rana, A.; Rera, M.; Pellegrini, M.; Ja, W.; et al. Distinct Shifts in Microbiota Composition during Drosophila Aging Impair Intestinal Function and Drive Mortality. Cell Rep. 2015, 12, 1656–1667. [Google Scholar] [CrossRef] [Green Version]
- Monchanin, C.; Devaud, J.-M.; Barron, A.B.; Lihoreau, M. Current permissible levels of metal pollutants harm terrestrial invertebrates. Sci. Total. Environ. 2021, 779, 146398. [Google Scholar] [CrossRef]
- Kalajdzic, P.; Kenig, B.; Andjelkovic, M. Drosophila subobscura flies adapted to low lead concentration carry no fitness cost. Environ. Pollut. 2015, 204, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Kenig, B.; Stamenković-Radak, M.; Andelković, M. Population specific fitness response of Drosophila subobscura to lead pollution. Insect Sci. 2013, 20, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Tanaskovic, M.; Novicic, Z.K.; Kenig, B.; Stamenkovic-Radak, M.; Andjelkovic, M. Effect of lead pollution on fitness and its dependence on heterozygosity in Drosophila subobscura. J. Genet. 2015, 94, 643–649. [Google Scholar] [CrossRef]
- Zhou, S.; Luoma, S.E.; St. Armour, G.E.; Thakkar, E.; Mackay, T.F.C.; Anholt, R.R.H. A Drosophila model for toxicogenomics: Genetic variation in susceptibility to heavy metal exposure. PLoS Genet. 2017, 13, e1006907. [Google Scholar] [CrossRef] [Green Version]
- Obadia, B.; Keebaugh, E.S.; Yamada, R.; Ludington, W.B.; Ja, W.W. Diet influences host–microbiota associations in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, E4547–E4548. [Google Scholar] [CrossRef] [Green Version]
- StatSoft Inc. STATISTICA (Data Analysis Software System), version 10, 10; StatSoft: Tulsa, OK, USA, 2011. [Google Scholar]
- Kapun, M.; Barrón, M.G.; Staubach, F.; Obbard, D.J.; Wiberg, R.A.W.; Vieira, J.; Goubert, C.; Rota-Stabelli, O.; Kankare, M.; Bogaerts-Márquez, M.; et al. Genomic Analysis of European Drosophila melanogaster Populations Reveals Longitudinal Structure, Continent-Wide Selection, and Previously Unknown DNA Viruses. Mol. Biol. Evol. 2020, 37, 2661–2678. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Kisand, V.; Cuadros, R.; Wikner, J. Phylogeny of culturable estuarine bacteria catabolizing riverine organic matter in the northern Baltic Sea. Appl. Environ. Microbiol. 2002, 68, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 2018, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavoine, S.; Dufour, A.-B.; Chessel, D. From dissimilarities among species to dissimilarities among communities: A double principal coordinate analysis. J. Theor. Biol. 2004, 228, 523–537. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R package version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 2 December 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Foster, Z.S.L.; Sharpton, T.J.; Grünwald, N.J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 2 December 2021).
- Lavasani, P.S.; Motevaseli, E.; Sanikhani, N.S.; Modarressi, M.H. Komagataeibacter xylinus as a novel probiotic candidate with high glucose conversion rate properties. Heliyon 2019, 5, e01571. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.C.; Kim, S.-H.; You, H.; Kim, B.; Kim, A.C.; Lee, K.-A.; Yoon, J.-H.; Ryu, J.-H.; Lee, W.-J. Drosophila microbiome modulates host developmental and metabolic homeostasis via insulin signaling. Science 2011, 334, 670–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Téfit, M.A.; Leulier, F. Lactobacillus plantarum favors the early emergence of fit and fertile adult Drosophila upon chronic undernutrition. J. Exp. Biol. 2017, 220, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-H.; Shang, J.; Yan, L.; Wei, T.; Xiang, L.; Wang, H.-L.; Cheng, J.; Xiao, G. Oxidative stress caused by lead (Pb) induces iron deficiency in Drosophila melanogaster. Chemosphere 2020, 243, 125428. [Google Scholar] [CrossRef]
- Martinez-Haro, M.; Green, A.J.; Mateo, R. Effects of lead exposure on oxidative stress biomarkers and plasma biochemistry in waterbirds in the field. Environ. Res. 2011, 111, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhao, X.; Yu, J.; Xie, J.; Li, C.; Liu, D.; Tang, C.; Wang, C. Lead-induced oxidative damage in rats/mice: A meta-analysis. J. Trace Elem. Med. Biol. 2020, 58, 126443. [Google Scholar] [CrossRef] [PubMed]
- Gurer-Orhan, H.; Sabır, H.U.; Özgüneş, H. Correlation between clinical indicators of lead poisoning and oxidative stress parameters in controls and lead-exposed workers. Toxicology 2004, 195, 147–154. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H.; Hu, M.; Cao, J.; Chen, D.; Liu, Z. Oxidative stress and apoptotic changes in primary cultures of rat proximal tubular cells exposed to lead. Arch. Toxicol. 2009, 83, 417–427. [Google Scholar] [CrossRef]
- Marcus, S.R.; Fiumera, A.C. Atrazine exposure affects longevity, development time and body size in Drosophila melanogaster. J. Insect Physiol. 2016, 91, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Figueira, F.H.; Aguiar, L.M.d.; Rosa, C.E.D. Embryo-larval exposure to atrazine reduces viability and alters oxidative stress parameters in Drosophila melanogaster. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2017, 191, 78–85. [Google Scholar] [CrossRef]
- Li, F.; Liu, Z.-H.; Tian, X.; Liu, T.; Wang, H.-L.; Xiao, G. Black soybean seed coat extract protects Drosophila melanogaster against Pb toxicity by promoting iron absorption. J. Funct. Foods 2020, 75, 104201. [Google Scholar] [CrossRef]
- Prasad, N.G.; Shakarad, M.; Gohil, V.M.; Sheeba, V.; Rajamani, M.; Joshi, A. Evolution of reduced pre-adult viability and larval growth rate in laboratory populations of Drosophila melanogaster selected for shorter development time. Genet. Res. 2000, 76, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chippindale, A.K.; Alipaz, J.A.; Chen, H.-W.; Rose, M.R. Experimental evolution of accelerated development in drosophila. 1. Developmental speed and larval survival. Evolution 1997, 51, 1536–1551. [Google Scholar] [CrossRef] [PubMed]
- Simhadri, R.K.; Fast, E.M.; Guo, R.; Schultz, M.J.; Vaisman, N.; Ortiz, L.; Bybee, J.; Slatko, B.E.; Frydman, H.M. The Gut Commensal Microbiome of Drosophila melanogaster Is Modified by the Endosymbiont Wolbachia. mSphere 2017, 2, e00287-17. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.H.; Seleznev, A.; Flores, H.A.; Woolfit, M.; McGraw, E.A. Gut microbiota in Drosophila melanogaster interacts with Wolbachia but does not contribute to Wolbachia-mediated antiviral protection. J. Invertebr. Pathol. 2017, 143, 18–25. [Google Scholar] [CrossRef]
- Modak, S.G.; Satish, K.; Mohan, J.; Dey, S.; Raghavendra, N.; Shakarad, M.; Joshi, A. A possible tradeoff between developmental rate and pathogen resistance in Drosophila melanogaster. J. Genet. 2009, 88, 253–256. [Google Scholar] [CrossRef]
- Sepulveda, J.; Moeller, A.H. The Effects of Temperature on Animal Gut Microbiomes. Front. Microbiol. 2020, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, A.; Castañeda, L.E. Gut Microbiota of Drosophila subobscura Contributes to Its Heat Tolerance and Is Sensitive to Transient Thermal Stress. Front. Microbiol. 2021, 12, 886. [Google Scholar] [CrossRef]
- Jiang, X.; Lin, D.; Shao, H.; Yang, X. Antioxidant properties of Komagataeibacter hansenii CGMCC 3917 and its ameliorative effects on alcohol-induced liver injury in mice. CyTA J. Food 2019, 17, 355–364. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
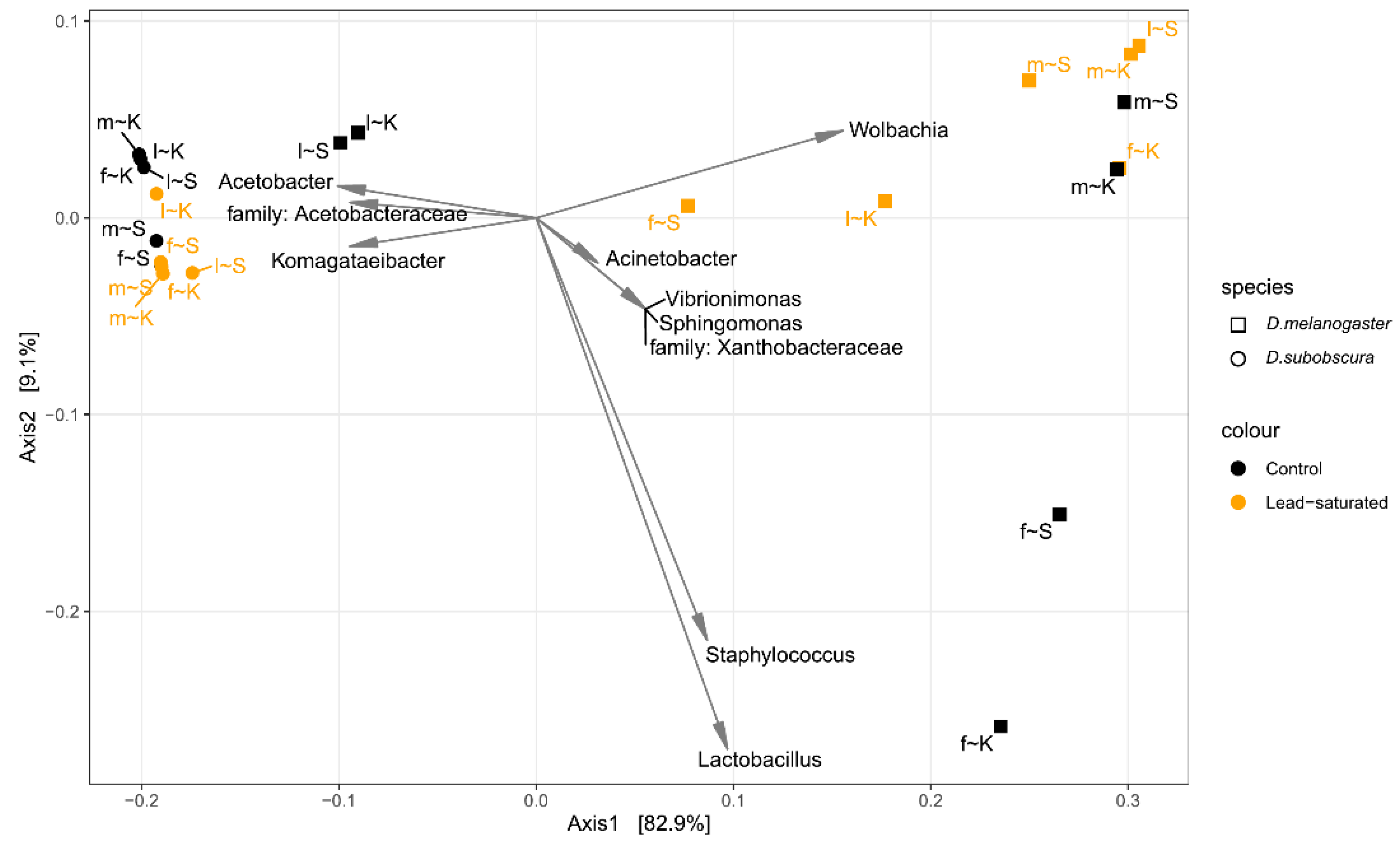{kind=link}
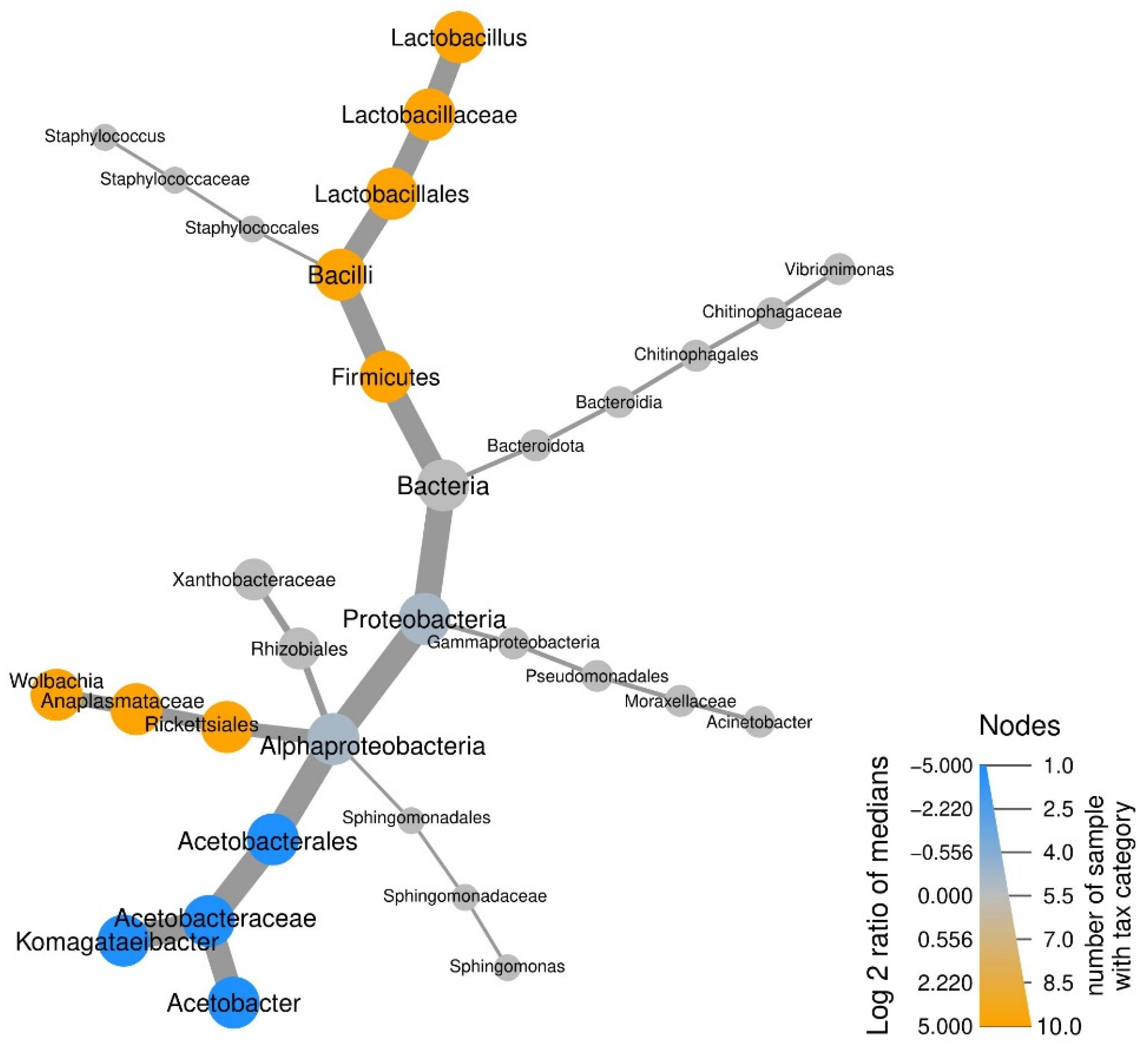{kind=link}
| Sample | Egg-to-Adult Viability | Developmental Time |
|---|---|---|
| Mean ± SE | Mean ± SE | |
| Dmel_K_St_M | 0.8763 ± 0.0068 | 19.567 ± 0.0482 |
| Dmel_K_St_F | 19.238 ± 0.0578 | |
| Dmel_Sl_St_M | 0.7333 ± 0.0180 | 19.782 ± 0.0727 |
| Dmel_Sl_St_F | 19.381 ± 0.0588 | |
| Dmel_K_C3_M | 0.5890 ± 0.0270 | 21.513 ± 0.1788 |
| Dmel_K_C3_F | 20.744 ± 0.1617 | |
| Dmel_Sl_C3_M | 0.5077 ± 0.0298 | 21.075 ± 0.1359 |
| Dmel_Sl_C3_F | 20.537 ± 0.1727 | |
| Dsub_K_St_M | 0.7297 ± 0.0166 | 20.580 ± 0.0593 |
| Dsub_K_St_F | 20.870 ± 0.0503 | |
| Dsub_Sl_St_M | 0.8053 ± 0.0165 | 21.420 ± 0.0715 |
| Dsub_Sl_St_F | 21.514 ± 0.0795 | |
| Dsub_K_C3_M | 0.7790 ± 0.0180 | 22.687 ± 0.0913 |
| Dsub_K_C3_F | 22.547 ± 0.0859 | |
| Dsub_Sl_C3_M | 0.7243 ± 0.0164 | 23.058 ± 0.1088 |
| Dsub_Sl_C3_F | 22.955 ± 0.0957 |
| Trait | Effect | df | SS | MS | F | p |
|---|---|---|---|---|---|---|
| (a) egg-to-adult viability | Population | 1 | 0.1550 | 0.1550 | 13.21 | 0.000343 |
| Substrate | 1 | 1.1125 | 1.1125 | 94.79 | 0.000000 | |
| Species | 1 | 0.4133 | 0.4133 | 35.22 | 0.000000 | |
| Population × Substrate | 1 | 0.0177 | 0.0177 | 1.51 | 0.220906 | |
| Population × Species | 1 | 0.2257 | 0.2257 | 19.23 | 0.000018 | |
| Substrate × Species | 1 | 0.8688 | 0.8688 | 74.03 | 0.000000 | |
| Population × Substrate × Species | 1 | 0.1382 | 0.1382 | 11.78 | 0.000709 | |
| (b) developmental time | Population | 1 | 7.3 | 7.3 | 22.3 | 0.000003 |
| Substrate | 1 | 305.5 | 305.5 | 931.2 | 0.000000 | |
| Species | 1 | 356.7 | 356.7 | 1087.1 | 0.000000 | |
| Sex | 1 | 6.7 | 6.7 | 20.6 | 0.000007 | |
| Population × Substrate | 1 | 5.5 | 5.5 | 16.7 | 0.000052 | |
| Population × Species | 1 | 12.2 | 12.2 | 37.2 | 0.000000 | |
| Substrate × Species | 1 | 1.7 | 1.7 | 5.3 | 0.022203 | |
| Population × Sex | 1 | 0.0 | 0.0 | 0.0 | 1.000000 | |
| Substrate × Sex | 1 | 2.7 | 2.7 | 8.3 | 0.004196 | |
| Species × Sex | 1 | 8.9 | 8.9 | 27.1 | 0.000000 | |
| Population × Substrate × Species | 1 | 0.2 | 0.2 | 0.5 | 0.476053 | |
| Population × Substrate × Sex | 1 | 0.5 | 0.5 | 1.6 | 0.199702 | |
| Population × Species × Sex | 1 | 0.2 | 0.2 | 0.6 | 0.445763 | |
| Substrate × Species × Sex | 1 | 0.0 | 0.0 | 0.0 | 0.906872 | |
| Population × Substrate × Species × Sex | 1 | 0.0 | 0.0 | 0.0 | 0.869297 |
| Sample | Observed | Chao1 | se.chao1 | Shannon | Gini-Simpson | InvSimpson |
|---|---|---|---|---|---|---|
| Dmel_K_St_M | 36 | 36.6 | 1.18 | 0.57 | 0.21 | 1.27 |
| Dmel_K_St_F | 28 | 33.6 | 5.34 | 1.29 | 0.64 | 2.76 |
| Dmel_Sl_St_M | 20 | 20 | 0 | 0.34 | 0.12 | 1.13 |
| Dmel_Sl_St_F | 16 | 17 | 2.29 | 1.17 | 0.56 | 2.29 |
| Dmel_K_C3_M | 10 | 10 | 0 | 0.14 | 0.05 | 1.05 |
| Dmel_K_C3_F | 11 | 11 | 0 | 0.46 | 0.20 | 1.24 |
| Dmel_Sl_C3_M | 10 | 10 | 0 | 0.49 | 0.24 | 1.31 |
| Dmel_Sl_C3_F | 9 | 9 | 0 | 1.07 | 0.58 | 2.41 |
| Dsub_K_St_M | 5 | 5 | 0 | 0.05 | 0.02 | 1.02 |
| Dsub_K_St_F | 5 | 5 | 0 | 0.11 | 0.03 | 1.04 |
| Dsub_Sl_St_M | 7 | 7 | 0.46 | 0.60 | 0.41 | 1.69 |
| Dsub_Sl_St_F | 5 | 5 | 0.22 | 0.29 | 0.15 | 1.17 |
| Dsub_K_C3_M | 3 | 3 | 0 | 0.07 | 0.03 | 1.03 |
| Dsub_K_C3_F | 3 | 3 | 0 | 0.08 | 0.03 | 1.03 |
| Dsub_Sl_C3_M | 4 | 4 | 0 | 0.25 | 0.13 | 1.15 |
| Dsub_Sl_C3_F | 3 | 3 | 0 | 0.35 | 0.20 | 1.25 |
| Dmel_St_K_L | 6 | 6 | 0 | 0.56 | 0.35 | 1.53 |
| Dmel_St_Sl_L | 6 | 6 | 0.46 | 0.55 | 0.33 | 1.48 |
| Dsub_St_K_L | 8 | 8 | 0 | 0.18 | 0.08 | 1.09 |
| Dsub_St_Sl_L | 10 | 10 | 0.47 | 0.42 | 0.19 | 1.24 |
| Dmel_C3_K_L | 6 | 6 | 0.46 | 1.02 | 0.52 | 2.08 |
| Dmel_C3_Sl_L | 7 | 7 | 0 | 0.07 | 0.02 | 1.02 |
| Dsub_C3_K_L | 20 | 20 | 0 | 0.61 | 0.35 | 1.54 |
| Dsub_C3_Sl_L | 18 | 21 | 4.15 | 0.89 | 0.55 | 2.22 |
| Effect | df | SS | MS | F | R2 | p |
|---|---|---|---|---|---|---|
| Species | 1 | 0.893082 | 0.893082 | 132.2773 | 0.651498 | 0.001 |
| Substrate | 1 | 0.046581 | 0.046581 | 6.899272 | 0.033981 | 0.02 |
| Sex | 2 | 0.100009 | 0.050004 | 7.406304 | 0.072956 | 0.008 |
| Species × Substrate | 1 | 0.034041 | 0.034041 | 5.041911 | 0.024833 | 0.029 |
| Species × Sex | 2 | 0.08629 | 0.043145 | 6.390341 | 0.062948 | 0.013 |
| Substrate × Sex | 2 | 0.065261 | 0.032631 | 4.833024 | 0.047608 | 0.014 |
| Species × Substrate × Sex | 2 | 0.06453 | 0.032265 | 4.778869 | 0.047074 | 0.02 |
| Residuals | 12 | 0.081019 | 0.006752 | 0.059103 | ||
| Total | 23 | 1.370812 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beribaka, M.; Jelić, M.; Tanasković, M.; Lazić, C.; Stamenković-Radak, M. Life History Traits in Two Drosophila Species Differently Affected by Microbiota Diversity under Lead Exposure. Insects 2021, 12, 1122. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121122
Beribaka M, Jelić M, Tanasković M, Lazić C, Stamenković-Radak M. Life History Traits in Two Drosophila Species Differently Affected by Microbiota Diversity under Lead Exposure. Insects. 2021; 12(12):1122. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121122
Chicago/Turabian StyleBeribaka, Mirjana, Mihailo Jelić, Marija Tanasković, Cvijeta Lazić, and Marina Stamenković-Radak. 2021. "Life History Traits in Two Drosophila Species Differently Affected by Microbiota Diversity under Lead Exposure" Insects 12, no. 12: 1122. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121122